
Lipidomics Reveals Elevated Plasmalogens in Women with Obesity Who Develop Preeclampsia
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Preeclampsia Definition
2.3. Sample Collection
2.4. Lipid Concentrations
2.5. Lipidomics
2.6. Statistical Analysis of Clinical Data
2.7. Broad Lipid Classes from Lipidomic Data
2.8. Lipidomic Statistical Analysis
2.8.1. Pre-Processing of Lipidomics Data
2.8.2. Univariate Analysis
2.8.3. Principal Component Analysis
2.8.4. Sparse Partial Least Squares Discriminant Analysis
3. Results
3.1. Maternal Demographics and Pregnancy Characteristics
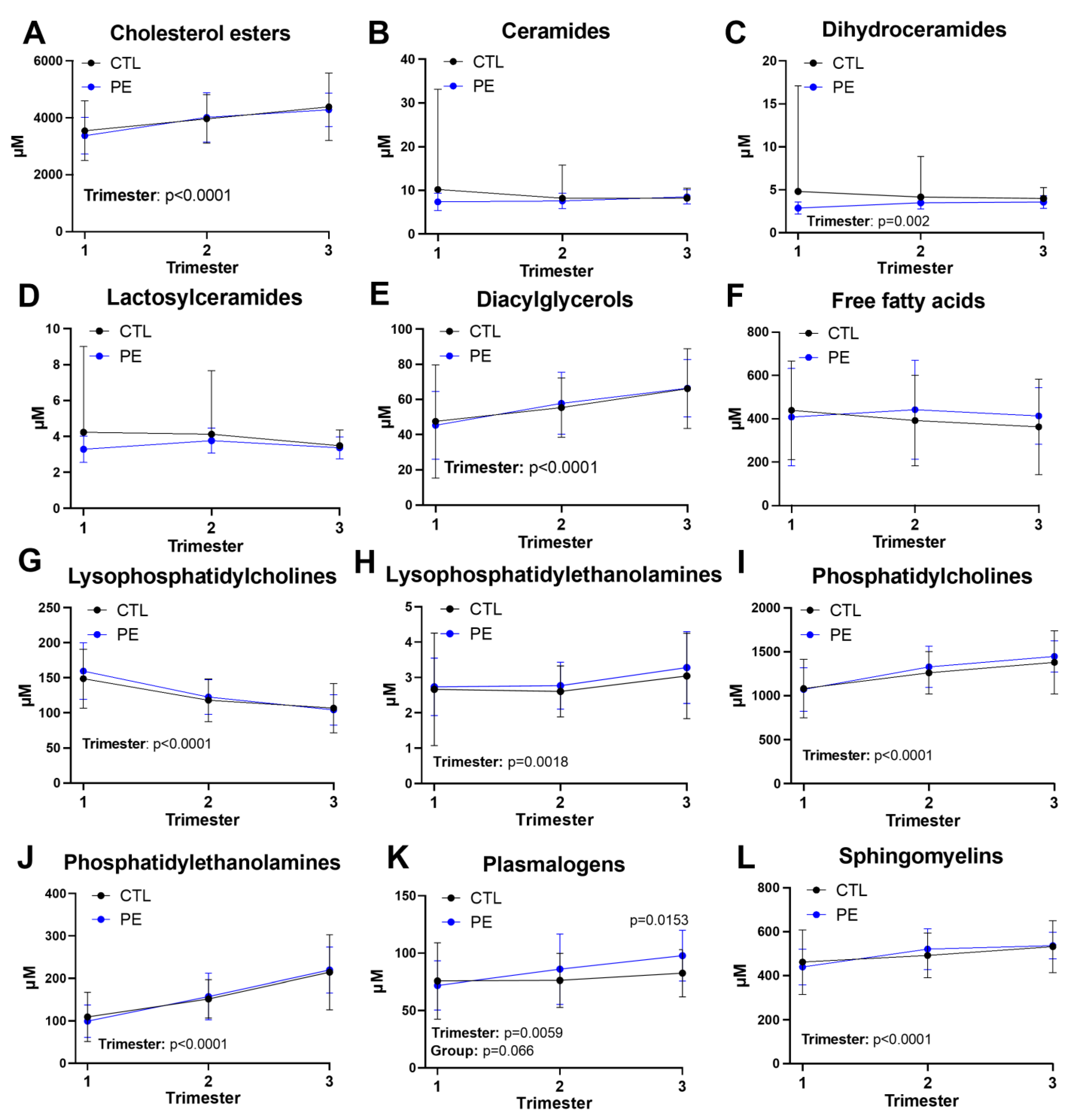
3.2. Dynamics of Broad Lipid Classes during Pregnancy
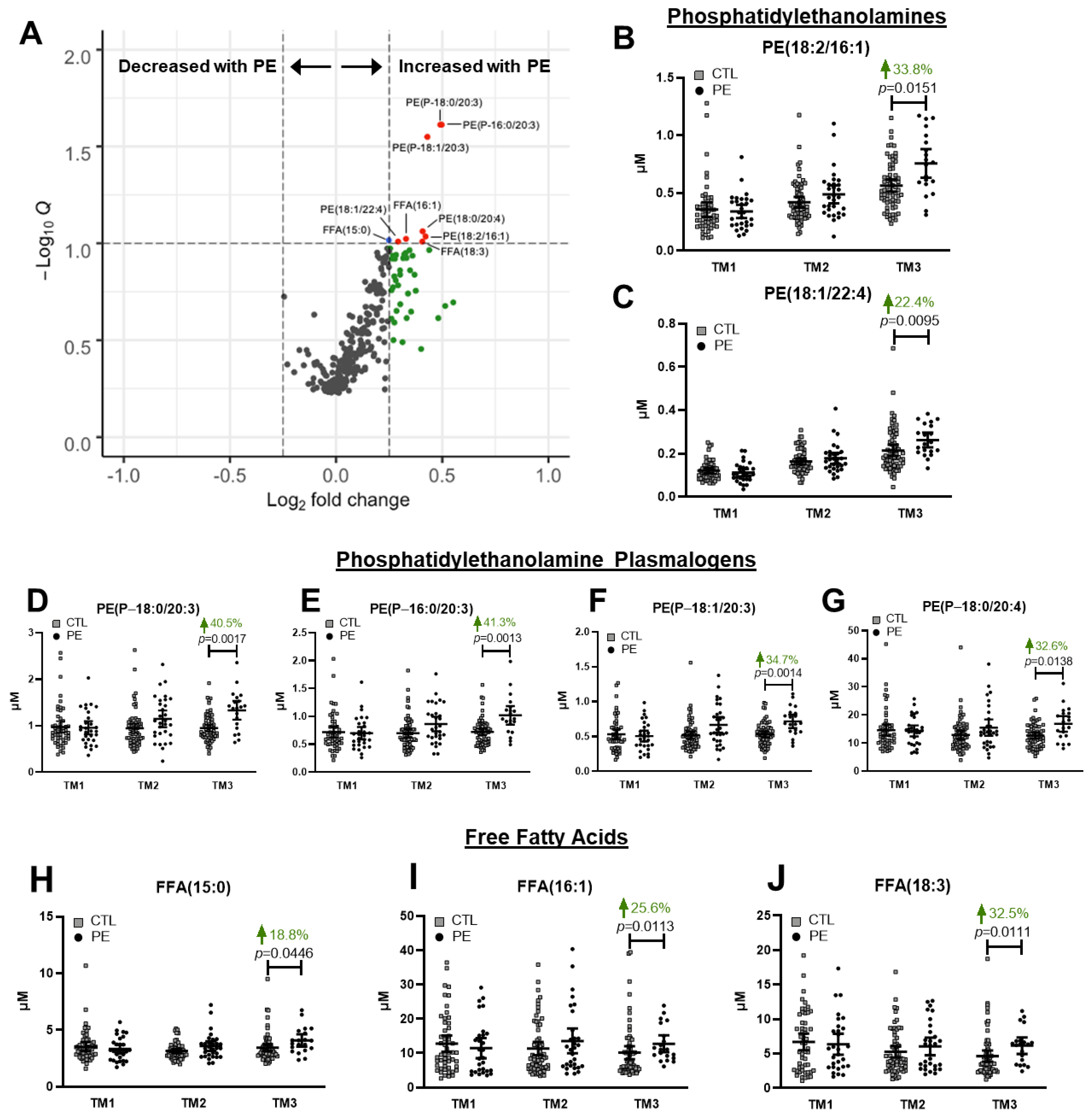
3.3. Third Trimester Differences in Individual Lipid Species between Uncomplicated and Preeclamptic Pregnancies
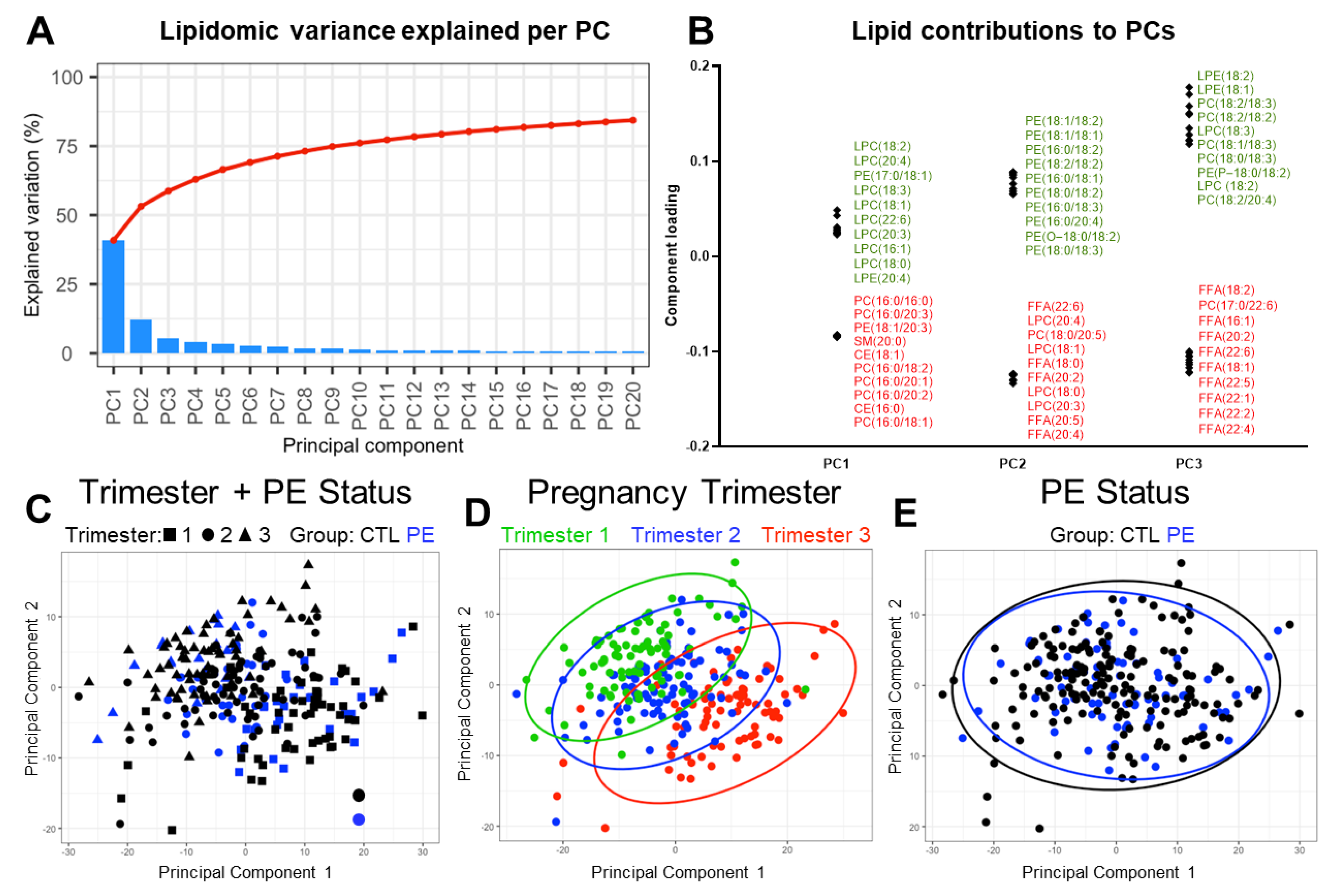
3.4. Exploratory Analysis of Factors Driving Plasma Lipidomic Variation
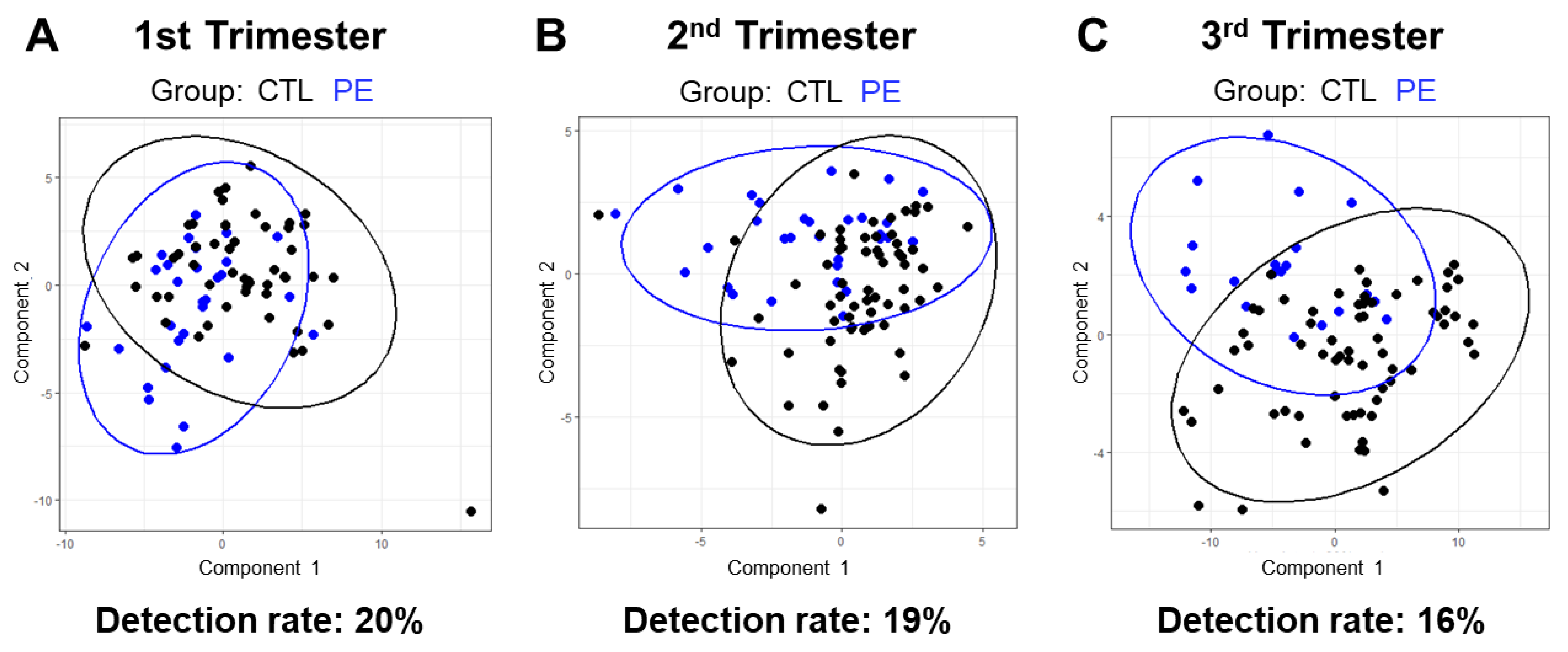
3.5. Lipidomic-Based Predictive Model of Pregnancy Outcome
3.6. Effects of Race and Fetal Sex on Plasma Lipids
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chappell, L.C.; Cluver, C.A.; Tong, S. Pre-eclampsia. Lancet 2021, 398, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Abalos, E.; Cuesta, C.; Grosso, A.L.; Chou, D.; Say, L. Global and regional estimates of preeclampsia and eclampsia: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Le Ray, I.; Zhu, J.; Zhang, J.; Hua, J.; Reilly, M. Preeclampsia Prevalence, Risk Factors, and Pregnancy Outcomes in Sweden and China. JAMA Netw. Open 2021, 4, e218401. [Google Scholar] [CrossRef]
- Lisonkova, S.; Joseph, K.S. Incidence of preeclampsia: Risk factors and outcomes associated with early- versus late-onset disease. Am. J. Obstet. Gynecol. 2013, 209, 544.e1–544.e12. [Google Scholar] [CrossRef]
- Rolnik, D.L.; Nicolaides, K.H.; Poon, L.C. Prevention of preeclampsia with aspirin. Am. J. Obstet. Gynecol. 2022, 226, S1108–S1119. [Google Scholar] [CrossRef] [PubMed]
- Askie, L.M.; Duley, L.; Henderson-Smart, D.J.; Stewart, L.A.; Group, P.C. Antiplatelet agents for prevention of pre-eclampsia: A meta-analysis of individual patient data. Lancet 2007, 369, 1791–1798. [Google Scholar] [CrossRef]
- Hofmeyr, G.J.; Lawrie, T.A.; Atallah, A.N.; Torloni, M.R. Calcium supplementation during pregnancy for preventing hypertensive disorders and related problems. Cochrane Database Syst. Rev. 2018, 10, CD001059. [Google Scholar] [CrossRef]
- Roberts, J.M.; Rich-Edwards, J.W.; McElrath, T.F.; Garmire, L.; Myatt, L.; Global Pregnancy, C. Subtypes of Preeclampsia: Recognition and Determining Clinical Usefulness. Hypertension 2021, 77, 1430–1441. [Google Scholar] [CrossRef]
- Bodnar, L.M.; Ness, R.B.; Markovic, N.; Roberts, J.M. The risk of preeclampsia rises with increasing prepregnancy body mass index. Ann. Epidemiol. 2005, 15, 475–482. [Google Scholar] [CrossRef]
- He, X.J.; Dai, R.X.; Hu, C.L. Maternal prepregnancy overweight and obesity and the risk of preeclampsia: A meta-analysis of cohort studies. Obes. Res. Clin. Pract. 2020, 14, 27–33. [Google Scholar] [CrossRef]
- Leung, T.Y.; Leung, T.N.; Sahota, D.S.; Chan, O.K.; Chan, L.W.; Fung, T.Y.; Lau, T.K. Trends in maternal obesity and associated risks of adverse pregnancy outcomes in a population of Chinese women. BJOG 2008, 115, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Meazaw, M.W.; Chojenta, C.; Muluneh, M.D.; Loxton, D. Systematic and meta-analysis of factors associated with preeclampsia and eclampsia in sub-Saharan Africa. PLoS ONE 2020, 15, e0237600. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Bodnar, L.M.; Patrick, T.E.; Powers, R.W. The Role of Obesity in Preeclampsia. Pregnancy Hypertens. 2011, 1, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.C.; Begum, S.; Seed, P.T.; Badran, D.; Briley, A.L.; Gill, C.; Godfrey, K.M.; Lawlor, D.A.; Nelson, S.M.; Patel, N.; et al. Gestational diabetes modifies the association between PlGF in early pregnancy and preeclampsia in women with obesity. Pregnancy Hypertens. 2018, 13, 267–272. [Google Scholar] [CrossRef]
- Vieira, M.C.; Poston, L.; Fyfe, E.; Gillett, A.; Kenny, L.C.; Roberts, C.T.; Baker, P.N.; Myers, J.E.; Walker, J.J.; McCowan, L.M.; et al. Clinical and biochemical factors associated with preeclampsia in women with obesity. Obesity 2017, 25, 460–467. [Google Scholar] [CrossRef]
- Ding, M.; Rexrode, K.M. A Review of Lipidomics of Cardiovascular Disease Highlights the Importance of Isolating Lipoproteins. Metabolites 2020, 10, 163. [Google Scholar] [CrossRef]
- Lee, S.M.; Kang, Y.; Lee, E.M.; Jung, Y.M.; Hong, S.; Park, S.J.; Park, C.W.; Norwitz, E.R.; Lee, D.Y.; Park, J.S. Metabolomic biomarkers in midtrimester maternal plasma can accurately predict the development of preeclampsia. Sci. Rep. 2020, 10, 16142. [Google Scholar] [CrossRef]
- Ubhi, B.K. Direct Infusion-Tandem Mass Spectrometry (DI-MS/MS) Analysis of Complex Lipids in Human Plasma and Serum Using the Lipidyzer Platform. Methods Mol. Biol. 2018, 1730, 227–236. [Google Scholar] [CrossRef]
- Global Pregnancy, C.; Schalekamp-Timmermans, S.; Arends, L.R.; Alsaker, E.; Chappell, L.; Hansson, S.; Harsem, N.K.; Jalmby, M.; Jeyabalan, A.; Laivuori, H.; et al. Fetal sex-specific differences in gestational age at delivery in pre-eclampsia: A meta-analysis. Int. J. Epidemiol. 2017, 46, 632–642. [Google Scholar] [CrossRef]
- Practice, A.C.o.O. Practice bulletin# 33: Diagnosis and management of preeclampsia and eclampsia. Obstet. Gynecol. 2002, 99, 159–167. [Google Scholar]
- Ubhi, B.K.; Novel Chemical Standards Kits Enable Facile Lipid Quantitation. SCIEX Technical Application Note RUO-MKT-02-3879-A. 2016. Available online: https://sciex.com/tech-notes/life-science-research/lipidomics/novel-chemical-standards-kits-enable-facile-lipid-quantitation (accessed on 10 April 2023).
- Smilde, A.K.; van der Werf, M.J.; Bijlsma, S.; van der Werff-van der Vat, B.J.; Jellema, R.H. Fusion of mass spectrometry-based metabolomics data. Anal. Chem. 2005, 77, 6729–6736. [Google Scholar] [CrossRef] [PubMed]
- Troyanskaya, O.; Cantor, M.; Sherlock, G.; Brown, P.; Hastie, T.; Tibshirani, R.; Botstein, D.; Altman, R.B. Missing value estimation methods for DNA microarrays. Bioinformatics 2001, 17, 520–525. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.A. mixOmics: An R package for ’omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed]
- Grimes, S.B.; Wild, R. Effect of Pregnancy on Lipid Metabolism and Lipoprotein Levels. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Hofland, J., Dungan, K., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Tan, M.Y.; Wright, D.; Syngelaki, A.; Akolekar, R.; Cicero, S.; Janga, D.; Singh, M.; Greco, E.; Wright, A.; Maclagan, K.; et al. Comparison of diagnostic accuracy of early screening for pre-eclampsia by NICE guidelines and a method combining maternal factors and biomarkers: Results of SPREE. Ultrasound Obstet. Gynecol. 2018, 51, 743–750. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Wiesner, P.; Leidl, K.; Boettcher, A.; Schmitz, G.; Liebisch, G. Lipid profiling of FPLC-separated lipoprotein fractions by electrospray ionization tandem mass spectrometry. J. Lipid Res. 2009, 50, 574–585. [Google Scholar] [CrossRef]
- Engelmann, B. Plasmalogens: Targets for oxidants and major lipophilic antioxidants. Biochem. Soc. Trans. 2004, 32, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Felde, R.; Spiteller, G. Plasmalogen oxidation in human serum lipoproteins. Chem. Phys. Lipids 1995, 76, 259–267. [Google Scholar] [CrossRef]
- Bozelli, J.C., Jr.; Azher, S.; Epand, R.M. Plasmalogens and Chronic Inflammatory Diseases. Front. Physiol. 2021, 12, 730829. [Google Scholar] [CrossRef]
- Donovan, E.L.; Pettine, S.M.; Hickey, M.S.; Hamilton, K.L.; Miller, B.F. Lipidomic analysis of human plasma reveals ether-linked lipids that are elevated in morbidly obese humans compared to lean. Diabetol. Metab. Syndr. 2013, 5, 24. [Google Scholar] [CrossRef]
- Heilbronn, L.K.; Coster, A.C.; Campbell, L.V.; Greenfield, J.R.; Lange, K.; Christopher, M.J.; Meikle, P.J.; Samocha-Bonet, D. The effect of short-term overfeeding on serum lipids in healthy humans. Obesity 2013, 21, E649–E659. [Google Scholar] [CrossRef] [PubMed]
- Piko, P.; Pal, L.; Szucs, S.; Kosa, Z.; Sandor, J.; Adany, R. Obesity-Related Changes in Human Plasma Lipidome Determined by the Lipidyzer Platform. Biomolecules 2021, 11, 326. [Google Scholar] [CrossRef] [PubMed]
- Sween, L.K.; Althouse, A.D.; Roberts, J.M. Early-pregnancy percent body fat in relation to preeclampsia risk in obese women. Am. J. Obstet. Gynecol. 2015, 212, 84.e1–84.e7. [Google Scholar] [CrossRef]
- Huynh, K.; Barlow, C.K.; Jayawardana, K.S.; Weir, J.M.; Mellett, N.A.; Cinel, M.; Magliano, D.J.; Shaw, J.E.; Drew, B.G.; Meikle, P.J. High-Throughput Plasma Lipidomics: Detailed Mapping of the Associations with Cardiometabolic Risk Factors. Cell Chem. Biol. 2019, 26, 71–84.e4. [Google Scholar] [CrossRef]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Vance, J.E. Lipoproteins secreted by cultured rat hepatocytes contain the antioxidant 1-alk-1-enyl-2-acylglycerophosphoethanolamine. Biochim. Biophys. Acta 1990, 1045, 128–134. [Google Scholar] [CrossRef]
- Ecker, J.; Liebisch, G. Application of stable isotopes to investigate the metabolism of fatty acids, glycerophospholipid and sphingolipid species. Prog. Lipid Res. 2014, 54, 14–31. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Z.; Jia, J.; Chiba, H.; Hui, S.P. Quantitative and Comparative Investigation of Plasmalogen Species in Daily Foodstuffs. Foods 2021, 10, 124. [Google Scholar] [CrossRef]
- Ikem, E.; Halldorsson, T.I.; Birgisdóttir, B.E.; Rasmussen, M.A.; Olsen, S.F.; Maslova, E. Dietary patterns and the risk of pregnancy-associated hypertension in the Danish National Birth Cohort: A prospective longitudinal study. BJOG 2019, 126, 663–673. [Google Scholar] [CrossRef]
- Kenny, L.C.; Broadhurst, D.I.; Dunn, W.; Brown, M.; North, R.A.; McCowan, L.; Roberts, C.; Cooper, G.J.; Kell, D.B.; Baker, P.N. Robust early pregnancy prediction of later preeclampsia using metabolomic biomarkers. Hypertension 2010, 56, 741–749. [Google Scholar] [CrossRef]
- Walsh, S.W. Eicosanoids in preeclampsia. Prostaglandins Leukot. Essent. Fatty Acids 2004, 70, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Watrous, J.D.; Niiranen, T.J.; Lagerborg, K.A.; Henglin, M.; Xu, Y.J.; Rong, J.; Sharma, S.; Vasan, R.S.; Larson, M.G.; Armando, A.; et al. Directed Non-targeted Mass Spectrometry and Chemical Networking for Discovery of Eicosanoids and Related Oxylipins. Cell Chem. Biol. 2019, 26, 433–442.e4. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uncomplicated Pregnancy (n = 67) | Preeclampsia (n = 33) | p-Value | |
|---|---|---|---|
| Age at delivery (years) | 23 (21, 26) | 24 (21, 27) | 0.61 |
| BMI pre-pregnancy (kg/m2) | 36.2 (33.0, 41.2) | 36.0 (32.1, 40.7) | 0.67 |
| Race (n,%) | |||
| Black | 44, 66% | 22, 67% | 0.92 |
| White | 22, 33% | 9, 27% | 0.57 |
| Other | 1, 1% | 2, 6% | |
| Cigarette smokers (n,%) | 26, 39% | 15, 45% | 0.53 |
| Primiparous (n,%) | 53, 73% | 23, 70% | 0.31 |
| Gestational age wks. at sampling | |||
| First trimester | 8.9 ± 2.3 (n = 52) | 8.1 ± 1.7 (n = 29) | 0.28 |
| Second trimester | 19.8 ± 1.3 (n = 63) | 19.7 ± 1.7 (n = 30) | 0.43 |
| Third trimester | 34.9 ± 0.8 (n = 65) | 35.1 ± 1.2 (n = 20) | 0.51 |
| Gestational age at delivery (wks) | 39 (38, 40) | 38 (35, 39) | <0.0001 |
| Birth weight gm | 3335 (3080, 3636) | 2820 (2282, 3445) | <0.0001 |
| Birth weight percentile 1 | 62 (36, 77) | 30 (10, 59) | 0.017 |
| First trimester blood pressure 2 | |||
| Systolic (mmHg) | 116.5 ± 8.3 | 116.2 ± 7.9 | 0.86 |
| Diastolic (mmHg) | 70.3 ± 6.8 | 73.3 ± 6.5 | 0.06 |
| Second trimester blood pressure 2 | |||
| Systolic (mmHg) | 113.3 ± 9.0 | 116.8 ± 8.6 | 0.073 |
| Diastolic (mmHg) | 66.8 ± 7.5 | 70.1 ± 7.9 | 0.049 |
| Third trimester blood pressure 2 | |||
| Systolic (mmHg) | 113.3 ± 10.2 | 119.0 ± 7.7 | 0.021 |
| Diastolic (mmHg) | 67.2 ± 8.4 | 71.5 ± 8.1 | 0.041 |
| Pre-delivery blood pressure2 | |||
| Systolic (mmHg) | 123 ± 8 | 146 ± 8 | <0.0001 |
| Diastolic (mmHg) | 71 ± 7 | 89 ± 8 | <0.0001 |
| Infant sex (n,% female) | 33, 49% | 14, 42% | |
| Preterm delivery < 37 wks (n,%) | 1, 1% | 12, 36% | <0.0001 |
| Severe preeclampsia (n,%) | 12, 36% | ||
| Gestational diabetes (n,%) | 6, 9% | 2, 6% | 0.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, I.M.; Albertolle, M.E.; Layden, A.J.; Tao, S.Y.; Fisher, S.J.; Gandley, R.E.; Roberts, J.M. Lipidomics Reveals Elevated Plasmalogens in Women with Obesity Who Develop Preeclampsia. J. Clin. Med. 2023, 12, 2970. https://doi.org/10.3390/jcm12082970
Williams IM, Albertolle ME, Layden AJ, Tao SY, Fisher SJ, Gandley RE, Roberts JM. Lipidomics Reveals Elevated Plasmalogens in Women with Obesity Who Develop Preeclampsia. Journal of Clinical Medicine. 2023; 12(8):2970. https://doi.org/10.3390/jcm12082970
Chicago/Turabian StyleWilliams, Ian M., Matthew E. Albertolle, Alexander J. Layden, Sunny Y. Tao, Susan J. Fisher, Robin E. Gandley, and James M. Roberts. 2023. "Lipidomics Reveals Elevated Plasmalogens in Women with Obesity Who Develop Preeclampsia" Journal of Clinical Medicine 12, no. 8: 2970. https://doi.org/10.3390/jcm12082970