
Posteroanterior Cervical Transcutaneous Spinal Cord Stimulation: Interactions with Cortical and Peripheral Nerve Stimulation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design
2.2. Participants
2.3. General Protocol
2.4. Electromyography (EMG)
2.5. Transcutaneous Spinal Cord Stimulation (TSCS)
2.6. Transcranial Magnetic Stimulation (TMS)
2.7. Peripheral Nerve Stimulation (PNS)
2.8. Hemodynamic Data Collection
2.9. TSCS-TMS Interactions
2.10. TSCS-PNS Interactions: F-Waves
2.11. TSCS-PNS Interactions: H-Reflexes
2.12. Data Analysis
2.13. Statistical Analysis
3. Results
3.1. Participants
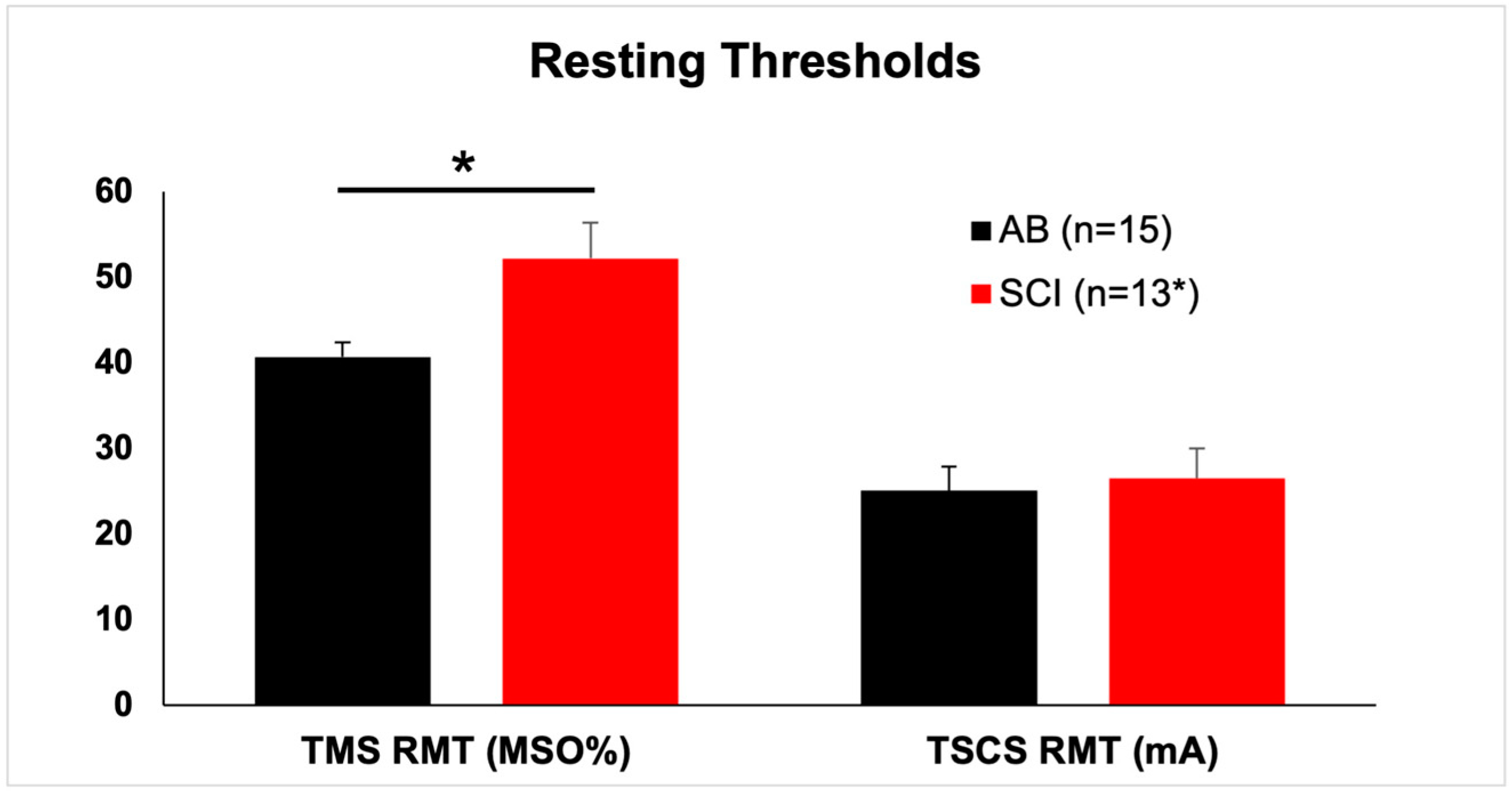
3.2. TMS Responses
3.3. TSCS Responses
3.4. PNS (F-Wave) Responses
3.5. PNS (H-Reflex) Responses
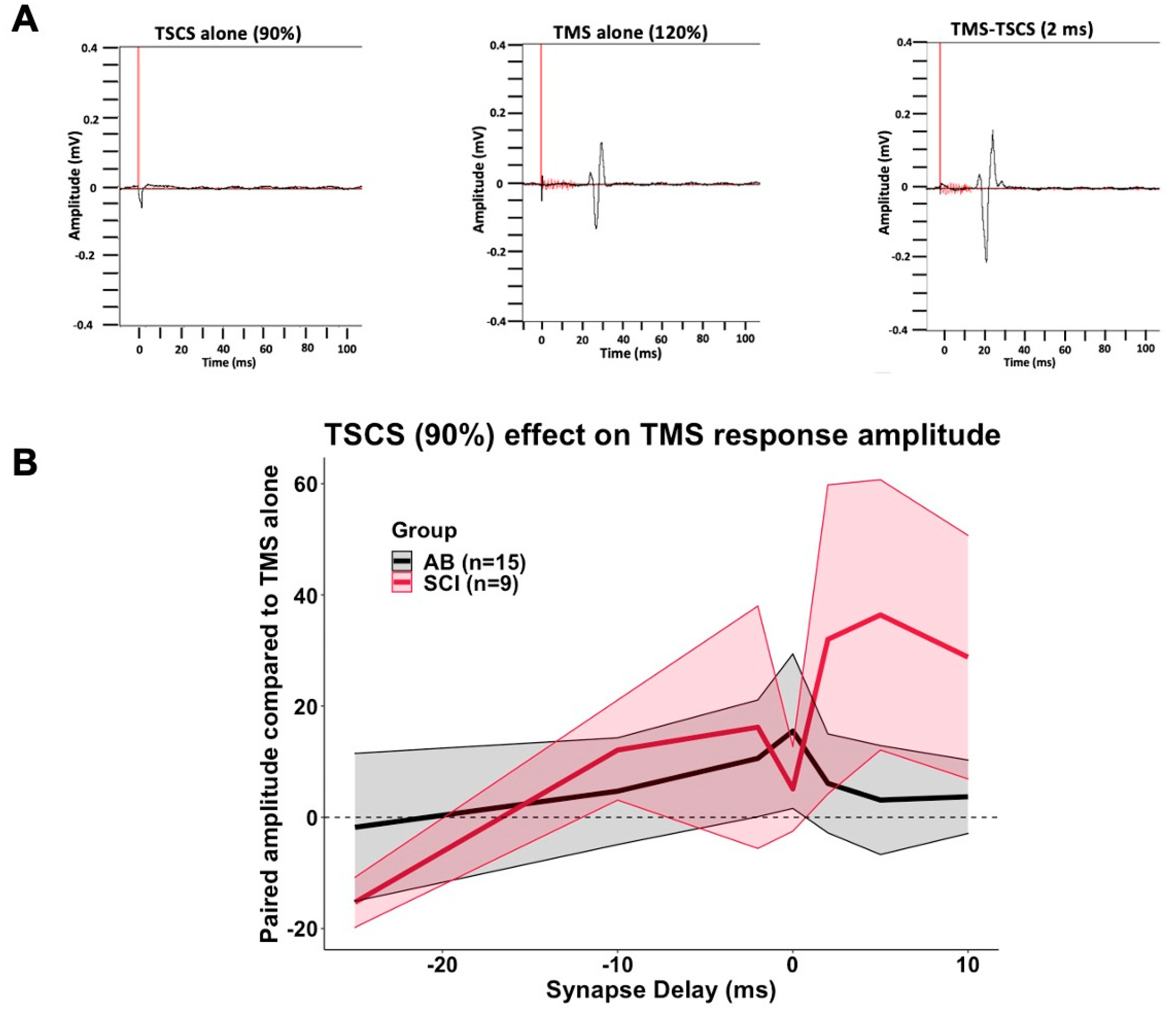
3.6. TSCS-TMS Interactions
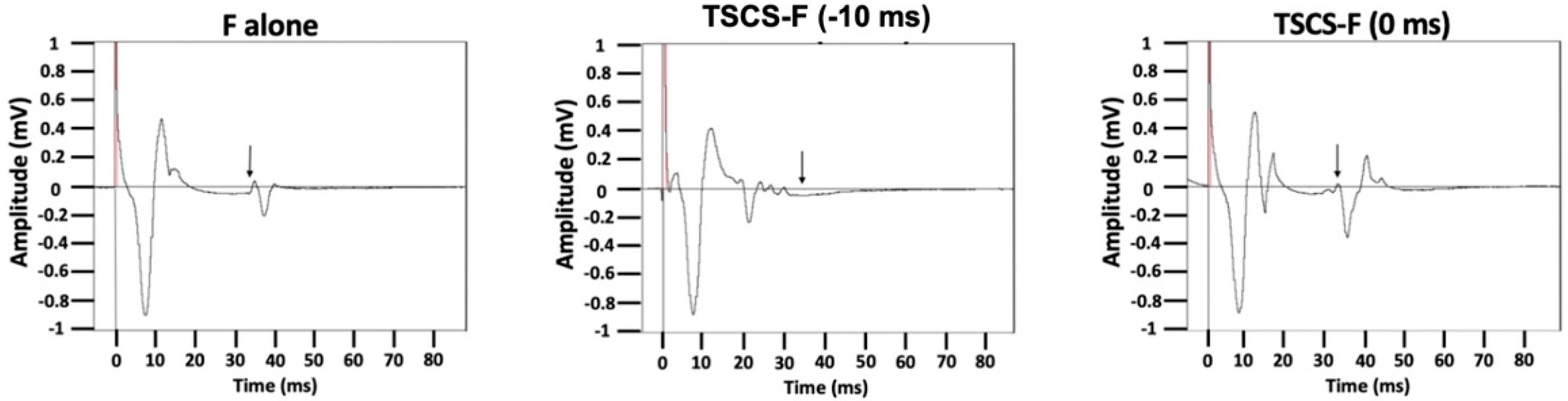
3.7. TSCS-PNS Interactions: F-Waves
3.8. TSCS-PNS Interactions: H-Reflexes
3.9. Safety and Hemodynamic Responses
4. Discussion
4.1. Limitations
4.2. Further Information and Experiments Needed
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harkema, S.; Gerasimenko, Y.; Hodes, J.; Burdick, J.; Angeli, C.; Chen, Y.; Ferreira, C.; Willhite, A.; Rejc, E.; Grossman, R.G.; et al. Effect of epidural stimulation of the lumbosacral spinal cord on voluntary movement, standing, and assisted stepping after motor complete paraplegia: A case study. Lancet 2011, 377, 1938–1947. [Google Scholar] [CrossRef] [Green Version]
- Angeli, C.A.; Edgerton, V.R.; Gerasimenko, Y.P.; Harkema, S.J. Altering spinal cord excitability enables voluntary movements after chronic complete paralysis in humans. Brain 2014, 137, 1394–1409. [Google Scholar] [CrossRef] [Green Version]
- Angeli, C.A.; Boakye, M.; Morton, R.A.; Vogt, J.; Benton, K.; Chen, Y.; Ferreira, C.K.; Harkema, S.J. Recovery of Over-Ground Walking after Chronic Motor Complete Spinal Cord Injury. N. Engl. J. Med. 2018, 379, 1244–1250. [Google Scholar] [CrossRef]
- Gad, P.; Lee, S.; Terrafranca, N.; Zhong, H.; Turner, A.; Gerasimenko, Y.; Edgerton, V.R. Non-Invasive Activation of Cervical Spinal Networks after Severe Paralysis. J. Neurotrauma 2018, 35, 2145–2158. [Google Scholar] [CrossRef]
- Inanici, F.; Samejima, S.; Gad, P.; Edgerton, V.R.; Hofstetter, C.P.; Moritz, C.T. Transcutaneous Electrical Spinal Stimulation Promotes Long-Term Recovery of Upper Extremity Function in Chronic Tetraplegia. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Inanici, F.; Brighton, L.N.; Samejima, S.; Hofstetter, C.P.; Moritz, C.T. Transcutaneous Spinal Cord Stimulation Restores Hand and Arm Function after Spinal Cord Injury. IEEE Trans. Neural Syst. Rehabil. Eng. 2021, 29, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Hofstoetter, U.S.; Krenn, M.; Danner, S.M.; Hofer, C.; Kern, H.; McKay, W.B.; Mayr, W.; Minassian, K. Augmentation of Voluntary Locomotor Activity by Transcutaneous Spinal Cord Stimulation in Motor-Incomplete Spinal Cord-Injured Individuals. Artif. Organs 2015, 39, E176–E186. [Google Scholar] [CrossRef]
- Gerasimenko, Y.P.; Lu, D.C.; Modaber, M.; Zdunowski, S.; Gad, P.; Sayenko, D.G.; Morikawa, E.; Haakana, P.; Ferguson, A.; Roy, R.R.; et al. Noninvasive Reactivation of Motor Descending Control after Paralysis. J. Neurotrauma 2015, 32, 1968–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minassian, K.; Hofstoetter, U.S.; Danner, S.M.; Mayr, W.; Bruce, J.A.; McKay, W.B.; Tansey, K.E. Spinal Rhythm Generation by Step-Induced Feedback and Transcutaneous Posterior Root Stimulation in Complete Spinal Cord–Injured Individuals. Neurorehabilit. Neural Repair 2016, 30, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Milosevic, M.; Masugi, Y.; Sasaki, A.; Sayenko, D.G.; Nakazawa, K. On the reflex mechanisms of cervical transcutaneous spinal cord stimulation in human subjects. J. Neurophysiol. 2019, 121, 1672–1679. [Google Scholar] [CrossRef]
- Wu, Y.-K.; Levine, J.M.; Wecht, J.R.; Maher, M.T.; Limonta, J.M.; Saeed, S.; Santiago, T.M.; Bailey, E.; Kastuar, S.; Guber, K.S.; et al. Posteroanterior cervical transcutaneous spinal stimulation targets ventral and dorsal nerve roots. Clin. Neurophysiol. 2020, 131, 451–460. [Google Scholar] [CrossRef]
- Formento, E.; Minassian, K.; Wagner, F.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Rowald, A.; Bloch, J.; Micera, S.; Capogrosso, M.; Courtine, G. Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury. Nat. Neurosci. 2018, 21, 1728–1741. [Google Scholar] [CrossRef] [PubMed]
- Minassian, K.; Persy, I.; Rattay, F.; Pinter, M.; Kern, H.; Dimitrijevic, M. Human lumbar cord circuitries can be activated by extrinsic tonic input to generate locomotor-like activity. Hum. Mov. Sci. 2007, 26, 275–295. [Google Scholar] [CrossRef]
- Gerasimenko, Y.; Gorodnichev, R.; Machueva, E.; Pivovarova, E.; Semyenov, D.; Savochin, A.; Roy, R.R.; Edgerton, V.R. Novel and Direct Access to the Human Locomotor Spinal Circuitry. J. Neurosci. 2010, 30, 3700–3708. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.M.; Pal, A.; Gupta, D.; Carmel, J.B. Paired motor cortex and cervical epidural electrical stimulation timed to converge in the spinal cord promotes lasting increases in motor responses. J. Physiol. 2017, 595, 6953–6968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Screening questionnaire before TMS: An update. Clin. Neurophysiol. 2011, 122, 1686. [Google Scholar] [CrossRef]
- Robinson, L.R.; Jantra, P.; MacLean, I.C. Central motor conduction times using transcranial stimulation and F Wave latencies. Muscle Nerve 1988, 11, 174–180. [Google Scholar] [CrossRef]
- Bunday, K.L.; Tazoe, T.; Rothwell, J.C.; Perez, M.A. Subcortical Control of Precision Grip after Human Spinal Cord Injury. J. Neurosci. 2014, 34, 7341–7350. [Google Scholar] [CrossRef] [Green Version]
- Bunday, K.L.; Perez, M.A. Motor Recovery after Spinal Cord Injury Enhanced by Strengthening Corticospinal Synaptic Transmission. Curr. Biol. 2012, 22, 2355–2361. [Google Scholar] [CrossRef] [Green Version]
- Mills, K.; Murray, N. Electrical stimulation over the human vertebral column: Which neural elements are excited? Electroencephalogr. Clin. Neurophysiol. 1986, 63, 582–589. [Google Scholar] [CrossRef]
- Stefan, K.; Kunesch, E.; Cohen, L.G.; Benecke, R.; Classen, J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain 2000, 123, 572–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, L.; Ibrahim, M.M.; Santora, D.; Knikou, M. Paired associative transspinal and transcortical stimulation produces plasticity in human cortical and spinal neuronal circuits. J. Neurophysiol. 2016, 116, 904–916. [Google Scholar] [CrossRef] [Green Version]
- Harel, N.Y.; Carmel, J.B. Paired Stimulation to Promote Lasting Augmentation of Corticospinal Circuits. Neural Plast. 2016, 2016, 7043767. [Google Scholar] [CrossRef] [PubMed]
- Roy, F.D.; Bosgra, D.; Stein, R.B. Interaction of transcutaneous spinal stimulation and transcranial magnetic stimulation in human leg muscles. Exp. Brain Res. 2014, 232, 1717–1728. [Google Scholar] [CrossRef]
- Ziemann, U.; Ilić, T.V.; Alle, H.; Meintzschel, F. Estimated magnitude and interactions of cortico-motoneuronal and Ia afferent input to spinal motoneurones of the human hand. Neurosci. Lett. 2004, 364, 48–52. [Google Scholar] [CrossRef]
- Cortes, M.; Thickbroom, G.W.; Valls-Sole, J.; Pascual-Leone, A.; Edwards, D.J. Spinal associative stimulation: A non-invasive stimulation paradigm to modulate spinal excitability. Clin. Neurophysiol. 2011, 122, 2254–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.L.; Martin, P.G. Voluntary Motor Output Is Altered by Spike-Timing-Dependent Changes in the Human Corticospinal Pathway. J. Neurosci. 2009, 29, 11708–11716. [Google Scholar] [CrossRef]
- D’amico, J.M.; Dongés, S.C.; Taylor, J.L. Paired corticospinal-motoneuronal stimulation increases maximal voluntary acti-vation of human adductor pollicis. J. Neurophysiol. 2018, 119, 369–376. [Google Scholar] [CrossRef]
- Urbin, M.A.; Ozdemir, R.A.; Tazoe, T.; Perez, M.A. Spike-timing-dependent plasticity in lower-limb motoneurons after human spinal cord injury. J. Neurophysiol. 2017, 118, 2171–2180. [Google Scholar] [CrossRef]
- Shulga, A.; Lioumis, P.; Kirveskari, E.; Savolainen, S.; Mäkelä, J.P.; Ylinen, A. The use of F-response in defining interstimulus intervals appropriate for LTP-like plasticity induction in lower limb spinal paired associative stimulation. J. Neurosci. Methods 2015, 242, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, A.N.; Jackson, A. Upper-limb muscle responses to epidural, subdural and intraspinal stimulation of the cervical spinal cord. J. Neural Eng. 2014, 11, 016005. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.; Zimmermann, J. Neural interfaces for the brain and spinal cord—Restoring motor function. Nat. Rev. Neurol. 2012, 8, 690–699. [Google Scholar] [CrossRef]
- Mercier, C.; Roosink, M.; Bouffard, J.; Bouyer, L. Promoting Gait Recovery and Limiting Neuropathic Pain After Spinal Cord Injury. Neurorehabilit. Neural Repair 2017, 31, 315–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Ramamurthy, A.; Lall, S.; Santos, J.; Ratnadurai-Giridharan, S.; Lopane, M.; Zareen, N.; Alexander, H.; Ryan, D.; Martin, J.H.; et al. Independent replication of motor cortex and cervical spinal cord electrical stimulation to promote forelimb motor function after spinal cord injury in rats. Exp. Neurol. 2019, 320, 112962. [Google Scholar] [CrossRef]
- Zareen, N.; Shinozaki, M.; Ryan, D.; Alexander, H.; Amer, A.; Truong, D.; Khadka, N.; Sarkar, A.; Naeem, S.; Bikson, M.; et al. Motor cortex and spinal cord neuromodulation promote corticospinal tract axonal outgrowth and motor recovery after cervical contusion spinal cord injury. Exp. Neurol. 2017, 297, 179–189. [Google Scholar] [CrossRef]
- Pal, A.; Park, H.; Ramamurthy, A.; Asan, S.; Bethea, T.; Johnkutty, M.; Carmel, J. Paired motor cortex and spinal cord epidural stimulation strengthens sensorimotor connections and improves forelimb function after cervical spinal cord injury in rats. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ting, W.K.-C.; Huot-Lavoie, M.; Ethier, C. Paired Associative Stimulation Fails to Induce Plasticity in Freely Behaving Intact Rats. eNeuro 2020, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumru, H.; Rodríguez-Cañón, M.; Edgerton, V.; García, L.; Flores, Á.; Soriano, I.; Opisso, E.; Gerasimenko, Y.; Navarro, X.; García-Alías, G.; et al. Transcutaneous Electrical Neuromodulation of the Cervical Spinal Cord Depends Both on the Stimulation Intensity and the Degree of Voluntary Activity for Training. A Pilot Study. J. Clin. Med. 2021, 10, 3278. [Google Scholar] [CrossRef]
- Thickbroom, G.W.; Byrnes, M.L.; Edwards, D.; Mastaglia, F.L. Repetitive paired-pulse TMS at I-wave periodicity markedly increases corticospinal excitability: A new technique for modulating synaptic plasticity. Clin. Neurophysiol. 2006, 117, 61–66. [Google Scholar] [CrossRef]
- Vastano, R.; Perez, M.A. Changes in motoneuron excitability during voluntary muscle activity in humans with spinal cord injury. J. Neurophysiol. 2020, 123, 454–461. [Google Scholar] [CrossRef]
- Thomas, C.K.; Häger, C.K.; Klein, C.S. Increases in human motoneuron excitability after cervical spinal cord injury depend on the level of injury. J. Neurophysiol. 2017, 117, 684–691. [Google Scholar] [CrossRef]
- Espiritu, M.G.; Lin, C.S.-Y.; Burke, D. Motoneuron excitability and the F wave. Muscle Nerve 2003, 27, 720–727. [Google Scholar] [CrossRef]
- Minassian, K.; Persy, I.; Rattay, F.; Dimitrijevic, M.R.; Hofer, C.; Kern, H. Posterior root–muscle reflexes elicited by transcutaneous stimulation of the human lumbosacral cord. Muscle Nerve 2007, 35, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Murray, L.M.; Knikou, M. Repeated cathodal transspinal pulse and direct current stimulation modulate cortical and corticospinal excitability differently in healthy humans. Exp. Brain Res. 2019, 237, 1841–1852. [Google Scholar] [CrossRef]
- Holtz, K.A.; Lipson, R.; Noonan, V.K.; Kwon, B.K.; Mills, P.B. Prevalence and Effect of Problematic Spasticity after Traumatic Spinal Cord Injury. Arch. Phys. Med. Rehabil. 2017, 98, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Sköld, C.; Levi, R.; Seiger, Å. Spasticity after traumatic spinal cord injury: Nature, severity, and location. Arch. Phys. Med. Rehabil. 1999, 80, 1548–1557. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; McKay, W.B.; Tansey, K.E.; Mayr, W.; Kern, H.; Minassian, K. Modification of spasticity by transcutaneous spinal cord stimulation in individuals with incomplete spinal cord injury. J. Spinal Cord Med. 2014, 37, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Hofstoetter, U.S.; Freundl, B.; Danner, S.M.; Krenn, M.J.; Mayr, W.; Binder, H.; Minassian, K. Transcutaneous Spinal Cord Stimulation Induces Temporary Attenuation of Spasticity in Individuals with Spinal Cord Injury. J. Neurotrauma 2020, 37, 481–493. [Google Scholar] [CrossRef]
- Goldsworthy, M.; Hordacre, B.; Ridding, M. Minimum number of trials required for within- and between-session reliability of TMS measures of corticospinal excitability. Neuroscience 2016, 320, 205–209. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Synaptic Delay (ms) | TSCS Intensity (% RMT) | ||
|---|---|---|---|
| TMS | F | H | |
| −200 | 50 | ||
| −100 | 50 | ||
| −50 | 50 | 50 | |
| −25 | 50, 70, 90 | 50 | 50 |
| −10 | 50, 70, 90 | 50, 70, 175 | 50 |
| −5 | 50, 70 | 50 | |
| −2 | 50, 70, 90 | 50, 70 | 50 |
| 0 | 50, 70, 90 | 50, 175 | 50 |
| 2 | 50, 70, 90 | ||
| 5 | 50, 70, 90 | ||
| 10 | 50, 70, 90 | ||
| SCI ID | Gender | Age | Trauma/NT | DOI (Years) | Level | Grade | Baclofen Use (Oral) |
|---|---|---|---|---|---|---|---|
| 1 | M | 64 | T | 35 | C4 | D | No |
| 3 | M | 54 | T | 13 | C5 | C | No |
| 5 | F | 22 | NT | 1.5 | C5 | C | Yes |
| 12 | M | 43 | T | 2 | C4 | D | Yes |
| 15 | M | 56 | T | 20 | C7 | D | No |
| 16 | M | 71 | T | 1.5 | C3 | D | Yes |
| 17 | M | 54 | T | 3 | C5 | D | No |
| 18 | M | 38 | T | 13 | C3 | C | No |
| 19 | F | 62 | T | 4 | C3 | D | No |
| 23 | M | 32 | T | 2 | C5 | C | No |
| 25 | M | 26 | T | 3 | C3 | B | No |
| 27 | F | 34 | T | 2 | C3 | A | Yes |
| 28 | M | 63 | T | 4 | C3 | C | Yes |
| AB ID | |||||||
| 2 | M | 46 | |||||
| 6 | M | 22 | |||||
| 7 | M | 55 | |||||
| 8 | M | 58 | |||||
| 9 | F | 52 | |||||
| 10 | M | 47 | |||||
| 11 | M | 60 | |||||
| 13 | F | 22 | |||||
| 14 | M | 22 | |||||
| 20 | M | 24 | |||||
| 21 | M | 45 | |||||
| 22 | M | 26 | |||||
| 24 | M | 24 | |||||
| 26 | M | 51 | |||||
| 29 | F | 27 |
| Participant | Group | TMS RMT | TMS120 Ampl | TSCS RMT |
|---|---|---|---|---|
| 2 | AB | 32.0 | 0.848 | 27.0 |
| 6 | AB | 38.0 | 0.714 | 26.3 |
| 7 | AB | 35.0 | 0.696 | 24.3 |
| 8 | AB | 46.0 | 0.581 | 45.0 |
| 9 | AB | 35.5 | 0.289 | 26.2 |
| 10 | AB | 30.0 | 0.665 | 30.8 |
| 11 | AB | 43.5 | 0.383 | 45.0 |
| 13 | AB | 35.0 | 0.511 | 4.0 |
| 14 | AB | 49.5 | 0.413 | 22.3 |
| 20 | AB | 41.0 | 0.176 | 16.7 |
| 21 | AB | 40.5 | 0.466 | 10.2 |
| 22 | AB | 41.5 | 0.518 | 26.7 |
| 24 | AB | 46.0 | 0.801 | 21.7 |
| 26 | AB | 52.0 | 0.758 | 24.5 |
| 29 | AB | 44.5 | 0.415 | 25.7 |
| AB | Mean | 40.7 | 0.549 | 25.1 |
| SEM | 1.7 | 0.051 | 2.8 | |
| 1 | SCI | 53.0 | 0.182 | 15.0 |
| 3 | SCI | 53.5 | 0.093 | 33.3 |
| 5 | SCI | 30.5 | 0.261 | 5.8 |
| 12 | SCI | 62.0 | 0.058 | 27.8 |
| 15 | SCI | 34.0 | 0.713 | 38.0 |
| 16 | SCI | 72.0 | 0.065 | 38.7 |
| 17 | SCI | 51.0 | 0.182 | 42.5 |
| 18 | SCI | 36.5 | 0.074 | 23.0 |
| 19 | SCI | 61.0 | 0.186 | 4.8 |
| 23 | SCI | 40.1 | ||
| 25 | SCI | 71.0 | 0.058 | 33.7 |
| 27 | SCI | 18.7 | ||
| 28 | SCI | 50.0 | 0.142 | 23.2 |
| SCI | Mean | 52.2 | 0.183 | 26.5 |
| SEM | 4.2 | 0.057 | 3.5 |
| Group | TSCS Intensity (% RMT) | Synapse Delay | Compared to TMS Alone (%) | SEM |
|---|---|---|---|---|
| AB | 50% | −25 | 0.2% | 9.4 |
| −10 | 15.4% | 16.7 | ||
| −2 | 0.6% | 8.9 | ||
| 0 | −11.4% | 8.9 | ||
| 2 | −0.5% | 7.1 | ||
| 5 | 8.1% | 16.2 | ||
| 10 | 8.7% | 9.1 | ||
| n/a | −98.9% | 0.6 | ||
| 70% | −25 | −2.7% | 4.8 | |
| −10 | 2.9% | 11.0 | ||
| −2 | 0.7% | 6.3 | ||
| 0 | 4.4% | 7.8 | ||
| 2 | 12.6% | 11.2 | ||
| 5 | 9.4% | 12.2 | ||
| 10 | 20.4% | 19.6 | ||
| n/a | −97.4% | 1.1 | ||
| 90% | −25 | −1.8% | 13.3 | |
| −10 | 4.7% | 9.6 | ||
| −2 | 10.6% | 10.5 | ||
| 0 | 15.5% | 13.9 | ||
| 2 | 6.1% | 8.9 | ||
| 5 | 3.1% | 9.8 | ||
| 10 | 3.7% | 6.6 | ||
| n/a | −93.1% | 1.9 | ||
| n/a | n/a | 0.0% | 0.0 | |
| SCI | 50% | −25 | 2.5% | 6.4 |
| −10 | 4.7% | 8.7 | ||
| −2 | −1.1% | 6.4 | ||
| 0 | −4.5% | 4.4 | ||
| 2 | 13.3% | 12.9 | ||
| 5 | −9.1% | 6.1 | ||
| 10 | 0.6% | 4.7 | ||
| n/a | −94.5% | 2.1 | ||
| 70% | −25 | −10.1% | 6.6 | |
| −10 | 1.5% | 7.3 | ||
| −2 | −2.9% | 6.5 | ||
| 0 | 5.8% | 5.4 | ||
| 2 | 7.6% | 12.8 | ||
| 5 | 9.6% | 8.1 | ||
| 10 | 17.8% | 8.2 | ||
| n/a | −92.0% | 3.1 | ||
| 90% | −25 | −15.3% | 4.5 | |
| −10 | 12.1% | 9.0 | ||
| −2 | 16.2% | 21.8 | ||
| 0 | 5.1% | 7.6 | ||
| 2 | 32.0% | 27.8 | ||
| 5 | 36.4% | 24.3 | ||
| 10 | 28.8% | 21.9 | ||
| n/a | −88.2% | 3.1 | ||
| n/a | n/a | 0.0% | 0.0 |
| Group | TSCS Intensity (% RMT) | Synapse Delay | Ampl Compared to F Alone (%) | SEM | Persistence Compared to F Alone (%) | SEM |
|---|---|---|---|---|---|---|
| AB | 50% | −200 | 23.1% | 11.56 | 8.9% | 6.0 |
| −50 | 6.7% | 8.6 | 5.7% | 6.3 | ||
| −25 | 1.7% | 8.3 | −2.8% | 6.6 | ||
| −10 | 7.2% | 7.2 | −4.3% | 5.5 | ||
| −5 | −3.8% | 11.6 | −5.8% | 5.5 | ||
| −2 | 12.0% | 10.8 | 0.3% | 5.0 | ||
| 0 | 0.8% | 7.3 | 1.8% | 6.0 | ||
| n/a | −92.8% | 2.1 | n/a | n/a | ||
| 70% | −10 | 4.3% | 7.7 | −2.6% | 8.0 | |
| −5 | 16.8% | 12.4 | 9.6% | 10.1 | ||
| −2 | 5.8% | 9.9 | 0.2% | 5.9 | ||
| n/a | −77.4% | 6.5 | n/a | n/a | ||
| 175% | −10 | −18.5% | 14.6 | −27.0% | 9.3 | |
| 0 | 328.5% | 136.1 | 39.5% | 18.6 | ||
| n/a | 1229.1% | 340.4 | n/a | n/a | ||
| n/a | n/a | 0.0% | 0.00 | 0.0% | 0.00 | |
| SCI | 50% | −200 | 9.5% | 7.7 | 6.0% | 2.9 |
| −50 | −1.5% | 11.6 | 11.2% | 8.9 | ||
| −25 | −11.0% | 9.0 | −4.5% | 9.4 | ||
| −10 | −11.5% | 8.9 | −7.2% | 8.0 | ||
| −5 | −1.9% | 6.4 | 5.7% | 3.1 | ||
| −2 | 17.2% | 16.9 | 9.2% | 7.2 | ||
| 0 | −9.9% | 11.3 | −9.9% | 9.8 | ||
| n/a | −77.7% | 9.5 | n/a | n/a | ||
| 70% | −10 | −17.6% | 6.5 | −5.8% | 4.0 | |
| −5 | −11.2% | 4.5 | −5.2% | 2.9 | ||
| −2 | −6.7% | 6.2 | 3.1% | 4.9 | ||
| n/a | −81.8% | 7.7 | n/a | n/a | ||
| 175% | −10 | −27.7% | 22.1 | −7.0% | 37.0 | |
| 0 | 371.4% | 189.1 | 75.4% | 35.8 | ||
| n/a | 1649.4% | 604.7 | n/a | n/a | ||
| n/a | n/a | 0.0% | 0.00 | 0.0% | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wecht, J.R.; Savage, W.M.; Famodimu, G.O.; Mendez, G.A.; Levine, J.M.; Maher, M.T.; Weir, J.P.; Wecht, J.M.; Carmel, J.B.; Wu, Y.-K.; et al. Posteroanterior Cervical Transcutaneous Spinal Cord Stimulation: Interactions with Cortical and Peripheral Nerve Stimulation. J. Clin. Med. 2021, 10, 5304. https://doi.org/10.3390/jcm10225304
Wecht JR, Savage WM, Famodimu GO, Mendez GA, Levine JM, Maher MT, Weir JP, Wecht JM, Carmel JB, Wu Y-K, et al. Posteroanterior Cervical Transcutaneous Spinal Cord Stimulation: Interactions with Cortical and Peripheral Nerve Stimulation. Journal of Clinical Medicine. 2021; 10(22):5304. https://doi.org/10.3390/jcm10225304
Chicago/Turabian StyleWecht, Jaclyn R., William M. Savage, Grace O. Famodimu, Gregory A. Mendez, Jonah M. Levine, Matthew T. Maher, Joseph P. Weir, Jill M. Wecht, Jason B. Carmel, Yu-Kuang Wu, and et al. 2021. "Posteroanterior Cervical Transcutaneous Spinal Cord Stimulation: Interactions with Cortical and Peripheral Nerve Stimulation" Journal of Clinical Medicine 10, no. 22: 5304. https://doi.org/10.3390/jcm10225304