
Brain Correlates of Persistent Postural-Perceptual Dizziness: A Review of Neuroimaging Studies

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
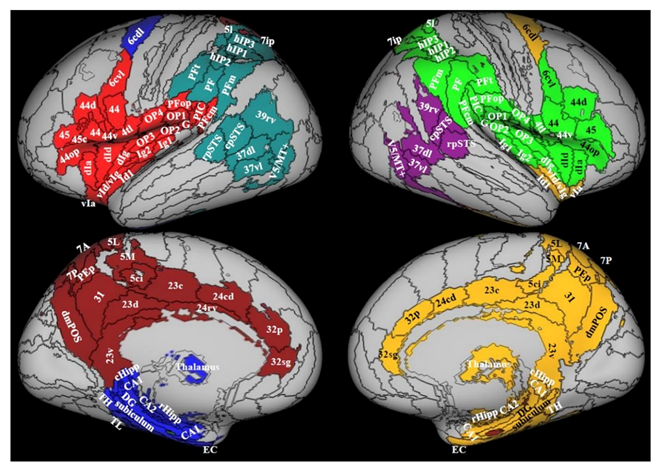
3.1. Comprehensive Screening of PPPD Neuroimaging Data
3.2. Structural Neuroimaging Studies
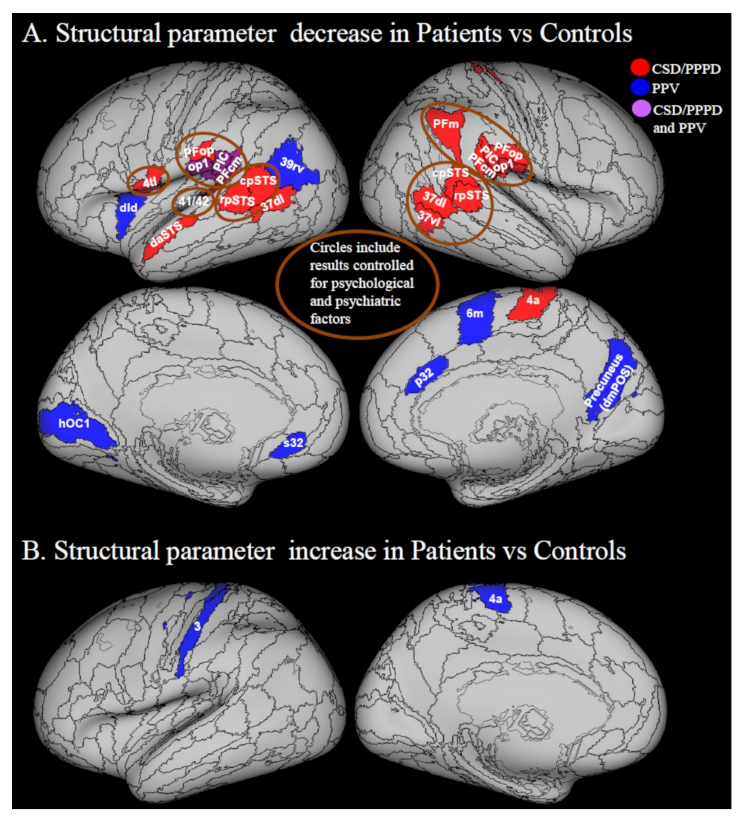
3.2.1. Decreases in Structural Neuroimaging Parameters (Grey Matter Volume, Cortical Thickness and Cortical Folding)
3.2.2. Increases in Structural Neuroimaging Parameters
3.3. Functional Neuroimaging Studies
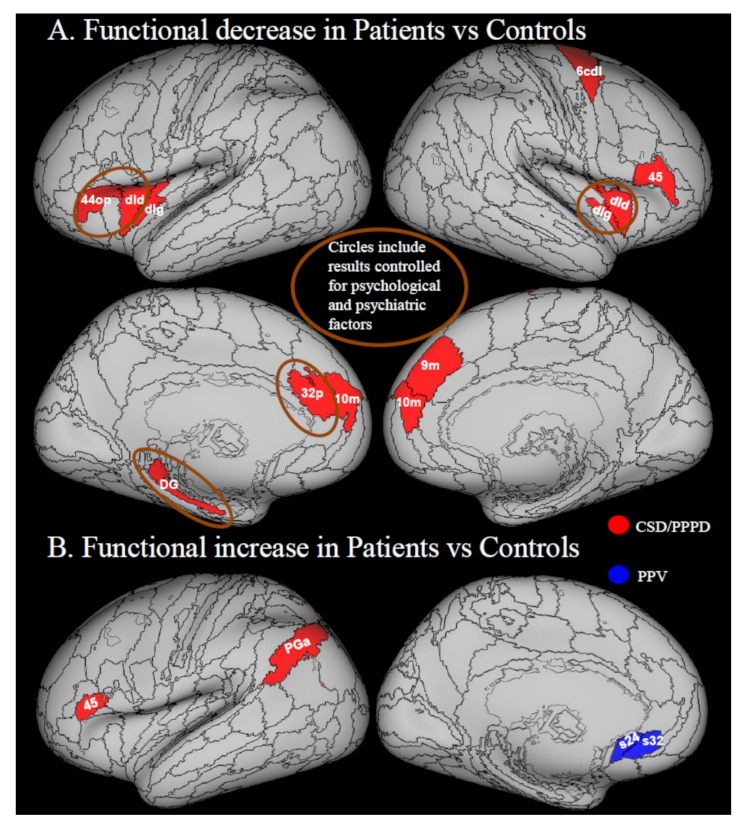
3.3.1. Decreases in Local Activity
3.3.2. Increases in Local Activity
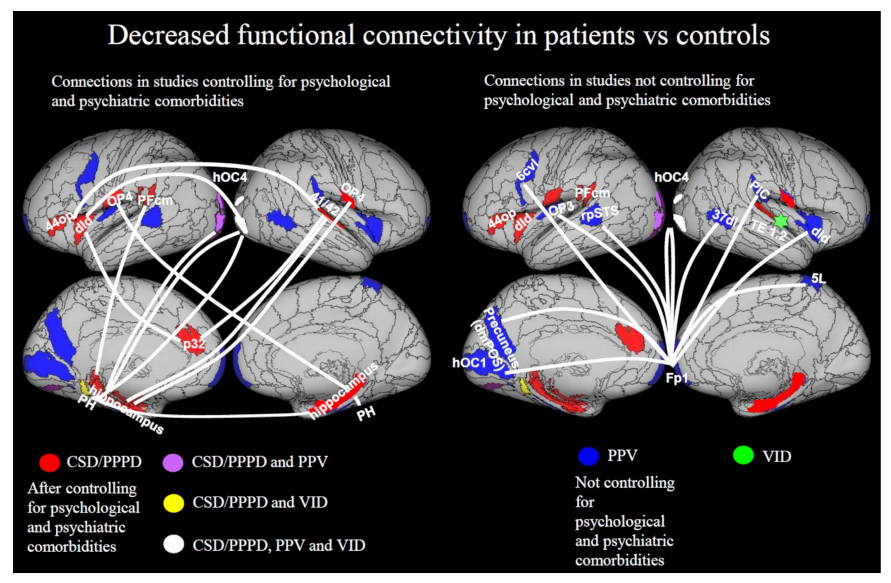
3.3.3. Decreases in Functional Connectivity
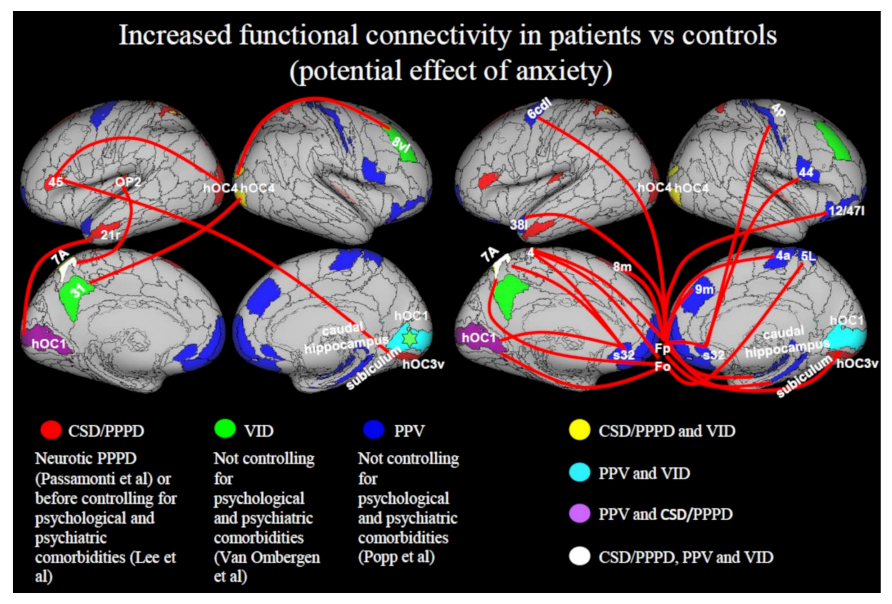
3.3.4. Increase in Functional Connectivity
4. Future Directions
4.1. The Role of Psychiatric Conditions in PPPD
4.2. Integration of Neuroimaging with Mathematical Models of Functional Disorders
4.3. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Staab, J.P.; Eckhardt-Henn, A.; Horii, A.; Jacob, R.; Strupp, M.; Brandt, T.; Bronstein, A. Diagnostic criteria for persistent postural-perceptual dizziness (PPPD): Consensus document of the committee for the Classification of Vestibular Disorders of the Bárány Society. J. Vestib. Res. 2017, 27, 191–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. ICD-11 for Mortality and Morbidity Statistics (Version 09/2020). AB32.0 Persistent Postur-al-Perceptual Dizziness. Available online: http://Id.Who.Int/Icd/Entity/2005792829 (accessed on 7 December 2020).
- Brandt, T.; Dieterich, M. Phobischer attacken-schwank-schwindel, ein neues syndrom? Münch Med Wochenschr 1986, 28, 247–250. [Google Scholar]
- Staab, J.P.; Ruckenstein, M.J.; Amsterdam, J.D. A prospective trial of sertraline for chronic subjective dizziness. Laryngoscope 2004, 114, 1637–1641. [Google Scholar] [CrossRef]
- Bronstein, A.M. Visual vertigo syndrome: Clinical and posturography findings. J. Neurol. Neurosurg. Psychiatry 1995, 59, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.G.; Lilienfeld, S.O.; Furman, J.M.; Durrant, J.D.; Turner, S.M. Panic disorder with vestibular dysfunction: Further clinical observations and description of space and motion phobic stimuli. J. Anxiety Disord. 1989, 3, 117–130. [Google Scholar] [CrossRef]
- Westphal, C. Die agoraphobie, eine neuropathische erscheinung. Arch. Psychiatr. 1872, 3, 138–161. [Google Scholar] [CrossRef]
- Dieterich, M.; Staab, J. Functional dizziness: From phobic postural vertigo and chronic subjective dizziness to persistent postural-perceptual dizziness. Curr. Opin. Neurol. 2017, 30, 107–113. [Google Scholar] [CrossRef]
- Staab, J. Functional and psychiatric vestibular disorders. Handb. Clin. Neurol. 2016, 137, 341–351. [Google Scholar] [CrossRef]
- Habs, M.; Strobl, R.; Grill, E.; Dieterich, M.; Becker-Bense, S. Primary or secondary chronic functional dizziness: Does it make a difference? A DizzyReg study in 356 patients. J. Neurol. 2020, 267, 212–222. [Google Scholar] [CrossRef]
- Staab, J.P.; Bronstein, A. Behavioural neuro-otology. In Oxford Textbook of Vertigo and Imbalance; Oxford University Press (OUP): Oxford, UK, 2013; pp. 333–346. [Google Scholar]
- Ishizuka, K.; Shikino, K.; Yamauchi, Y.; Yanagita, Y.; Yokokawa, D.; Ikegami, A.; Tsukamoto, T.; Noda, K.; Uehara, T.; Ikusaka, M. The clinical key features of persistent postural perceptual dizziness in the general medicine outpatient setting: A case series study of 33 patients. Intern. Med. 2020, 59, 2857–2862. [Google Scholar] [CrossRef]
- Adamec, I.; Meaški, S.J.; Skorić, M.K.; Jažić, K.; Crnošija, L.; Milivojević, I.; Habek, M. Persistent postural-perceptual dizziness: Clinical and neurophysiological study. J. Clin. Neurosci. 2020, 72, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Lee, J.-O.; Choi, J.-Y.; Kim, J.-S. Etiologic distribution of dizziness and vertigo in a referral-based dizziness clinic in South Korea. J. Neurol. 2020, 267, 2252–2259. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Chong, Y.; Jiang, Z.D.; Liu, Z.L.; Ding, L.; Yang, S.L.; Wang, L.; Xiang, W.P. Etiological analysis on patients with vertigo or dizziness. Zhonghua Yi Xue Za Zhi 2018, 98, 1227–1230. [Google Scholar]
- Staibano, P.; Lelli, D.; Tse, D. A retrospective analysis of two tertiary care dizziness clinics: A multidisciplinary chronic dizziness clinic and an acute dizziness clinic. J. Otolaryngol. Head Neck Surg. 2019, 48, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Brandt, T.; Huppert, D.; Strupp, M.; Dieterich, M. Functional dizziness: Diagnostic keys and differential diagnosis. J. Neurol. 2015, 262, 1977–1980. [Google Scholar] [CrossRef] [PubMed]
- Staab, J.P. Persistent postural-perceptual dizziness. Semin. Neurol. 2020, 40, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Chiarella, G.; Petrolo, C.; Riccelli, R.; Giofrè, L.; Olivadese, G.; Gioacchini, F.; Scarpa, A.; Cassandro, E.; Passamonti, L. Chronic subjective dizziness: Analysis of underlying personality factors. J. Vestib. Res. 2016, 26, 403–408. [Google Scholar] [CrossRef]
- Staab, J.P.; Rohe, D.E.; Eggers, S.D.; Shepard, N.T. Anxious, introverted personality traits in patients with chronic subjective dizziness. J. Psychosom. Res. 2014, 76, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Cui, L.; Yu, T.; Liang, H.; Wang, Y.; Chen, C. Analysis of the characteristics of persistent postural-perceptual dizziness: A clinical-based study in China. Int. J. Audiol. 2016, 56, 33–37. [Google Scholar] [CrossRef]
- Staab, J.P.; Ruckenstein, M.J. Which comes first? Psychogenic dizziness versus otogenic anxiety. Laryngoscope 2010, 113, 1714–1718. [Google Scholar] [CrossRef]
- Cousins, S.; Kaski, D.; Cutfield, N.; Arshad, Q.; Ahmad, H.; Gresty, M.A.; Seemungal, B.; Golding, J.; Bronstein, A.M. Predictors of clinical recovery from vestibular neuritis: A prospective study. Ann. Clin. Transl. Neurol. 2017, 4, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Godemann, F.; Siefert, K.; Hantschke-Brüggemann, M.; Neu, P.; Seidl, R.; Strohle, A. What accounts for vertigo one year after neuritis vestibularis—Anxiety or a dysfunctional vestibular organ? J. Psychiatr. Res. 2005, 39, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, N.; Edler, C.; Eskens, S.; Mielczarek, M.M.; Moschner, C. Predicting continued dizziness after an acute peripheral vestibular disorder. Psychosom. Med. 2007, 69, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Trinidade, A.; Harman, P.; Stone, J.; Staab, J.P.; Goebel, J.A. Assessment of potential risk factors for the development of persistent postural-perceptual dizziness: A case-control pilot study. Front. Neurol. 2021, 11, 1955. [Google Scholar] [CrossRef] [PubMed]
- Ödman, M.; Maire, R. Chronic subjective dizziness. Acta Oto-Laryngol. 2008, 128, 1085–1088. [Google Scholar] [CrossRef]
- Schniepp, R.; Wuehr, M.; Huth, S.; Pradhan, C.; Brandt, T.; Jahn, K. Gait characteristics of patients with phobic postural vertigo: Effects of fear of falling, attention, and visual input. J. Neurol. 2014, 261, 738–746. [Google Scholar] [CrossRef]
- Wuehr, M.; Brandt, T.; Schniepp, R. Distracting attention in phobic postural vertigo normalizes leg muscle activity and balance. Neurology 2016, 88, 284–288. [Google Scholar] [CrossRef]
- Wuehr, M.; Kugler, G.; Schniepp, R.; Eckl, M.; Pradhan, C.; Jahn, K.; Huppert, D.; Brandt, T. Balance control and anti-gravity muscle activity during the experience of fear at heights. Physiol. Rep. 2014, 2, e00232. [Google Scholar] [CrossRef]
- Wuehr, M.; Pradhan, C.; Novozhilov, S.; Krafczyk, S.; Brandt, T.; Jahn, K.; Schniepp, R. Inadequate interaction between open- and closed-loop postural control in phobic postural vertigo. J. Neurol. 2013, 260, 1314–1323. [Google Scholar] [CrossRef]
- Cousins, S.; Cutfield, N.J.; Kaski, D.; Palla, A.; Seemungal, B.; Golding, J.F.; Staab, J.; Bronstein, A.M. Visual dependency and dizziness after vestibular neuritis. PLoS ONE 2014, 9, e105426. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Arshad, Q.; Roberts, E.; Ahmad, H.; Bronstein, A.M. Chronic symptoms after vestibular neuritis and the high-velocity vestibulo-ocular reflex. Otol. Neurotol. 2016, 37, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Pavlou, M.; Kanegaonkar, R.; Swapp, D.; Bamiou, D.; Slater, M.; Luxon, L. The effect of virtual reality on visual vertigo symptoms in patients with peripheral vestibular dysfunction: A pilot study. J. Vestib. Res. 2012, 22, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Teggi, R.; Gatti, O.; Cangiano, J.; Fornasari, F.; Bussi, M. Functional head impulse test with and without optokinetic stimulation in subjects with persistent postural perceptual dizziness (PPPD): Preliminary report. Otol. Neurotol. 2020, 41, e70–e75. [Google Scholar] [CrossRef] [PubMed]
- Van Ombergen, A.; Heine, L.; Jillings, S.; Roberts, R.E.; Jeurissen, B.; van Rompaey, V.; Mucci, V.; Vanhecke, S.; Sijbers, J.; Vanhevel, F.; et al. Altered functional brain connectivity in patients with visually induced dizziness. NeuroImage Clin. 2017, 14, 538–545. [Google Scholar] [CrossRef]
- Van Ombergen, A.; Lubeck, A.J.; van Rompaey, V.; Maes, L.K.; Stins, J.F.; van de Heyning, P.H.; Wuyts, F.L.; Bos, J.E. The effect of optokinetic stimulation on perceptual and postural symptoms in visual vestibular mismatch patients. PLoS ONE 2016, 11, e0154528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaban, C.D.; Thayer, J.F. Neurological bases for balance-anxiety links. J. Anxiety Disord. 2001, 15, 53–79. [Google Scholar] [CrossRef]
- Indovina, I.; Riccelli, R.; Chiarella, G.; Petrolo, C.; Augimeri, A.; Giofrè, L.; Lacquaniti, F.; Staab, J.; Passamonti, L. Role of the insula and vestibular system in patients with chronic subjective dizziness: An fMRI study using sound-evoked vestibular stimulation. Front. Behav. Neurosci. 2015, 9, 334. [Google Scholar] [CrossRef] [PubMed]
- Söhsten, E.; Bittar, R.S.; Staab, J.P. Posturographic profile of patients with persistent postural-perceptual dizziness on the sensory organization test. J. Vestib. Res. 2016, 26, 319–326. [Google Scholar] [CrossRef]
- Sun, L.; Xiang, K. A review on the alterations in the brain of persistent postural-perceptual dizziness patients and non-pharmacological interventions for its management. Rev. Neurosci. 2020, 31, 675–680. [Google Scholar] [CrossRef]
- Lee, J.-O.; Lee, E.-S.; Kim, J.-S.; Lee, Y.-B.; Jeong, Y.; Choi, B.S.; Kim, J.-H.; Staab, J.P. Altered brain function in persistent postural perceptual dizziness: A study on resting state functional connectivity. Hum. Brain Mapp. 2018, 39, 3340–3353. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Si, L.; Cui, B.; Ling, X.; Shen, B.; Yang, X. Altered spontaneous functional activity of the right precuneus and cuneus in patients with persistent postural-perceptual dizziness. Brain Imaging Behav. 2020, 14, 2176–2186. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Si, L.; Cui, B.; Ling, X.; Shen, B.; Yang, X. Altered intra- and inter-network functional connectivity in patients with persistent postural-perceptual dizziness. NeuroImage Clin. 2020, 26, 102216. [Google Scholar] [CrossRef]
- Na, S.; Im, J.J.; Jeong, H.; Lee, E.-S.; Lee, T.-K.; Chung, Y.-A.; Song, I.-U. Cerebral perfusion abnormalities in patients with persistent postural-perceptual dizziness (PPPD): A SPECT study. J. Neural Transm. 2019, 126, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Nigro, S.; Indovina, I.; Riccelli, R.; Chiarella, G.; Petrolo, C.; Lacquaniti, F.; Staab, J.P.; Passamonti, L. Reduced cortical folding in multi-modal vestibular regions in persistent postural perceptual dizziness. Brain Imaging Behav. 2019, 13, 798–809. [Google Scholar] [CrossRef] [Green Version]
- Passamonti, L.; Riccelli, R.; Lacquaniti, F.; Staab, J.P.; Indovina, I. Brain responses to virtual reality visual motion stimulation are affected by neurotic personality traits in patients with persistent postural-perceptual dizziness. J. Vestib. Res. 2019, 28, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Popp, P.; Zu Eulenburg, P.; Stephan, T.; Bögle, R.; Habs, M.; Henningsen, P.; Feuerecker, R.; Dieterich, M. Cortical alterations in phobic postural vertigo—A multimodal imaging approach. Ann. Clin. Transl. Neurol. 2018, 5, 717–729. [Google Scholar] [CrossRef]
- Riccelli, R.; Passamonti, L.; Toschi, N.; Nigro, S.; Chiarella, G.; Petrolo, C.; Lacquaniti, F.; Staab, J.; Indovina, I. Altered insular and occipital responses to simulated vertical self-motion in patients with persistent postural-perceptual dizziness. Front. Neurol. 2017, 8, 529. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.; Ahmad, H.; Patel, M.; Dima, D.; Ibitoye, R.; Sharif, M.; Leech, R.; Arshad, Q.; Bronstein, A. An fMRI study of visuo-vestibular interactions following vestibular neuritis. NeuroImage Clin. 2018, 20, 1010–1017. [Google Scholar] [CrossRef]
- Wurthmann, S.; Naegel, S.; Steinberg, B.S.; Theysohn, N.; Diener, H.-C.; Kleinschnitz, C.; Obermann, M.; Holle, D. Cerebral gray matter changes in persistent postural perceptual dizziness. J. Psychosom. Res. 2017, 103, 95–101. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Stephan, K.E.; Mohlberg, H.; Grefkes, C.; Fink, G.R.; Amunts, K.; Zilles, K. A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. NeuroImage 2005, 25, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Lingzhong, F.; Zhuo, J.; Zhang, Y.; Wang, J.; Chen, L.; Yang, Z.; Chu, C.; Xie, S.; Laird, A.; et al. The human brainnetome atlas: A new brain atlas based on connectional architecture. Cereb. Cortex 2016, 26, 3508–3526. [Google Scholar] [CrossRef] [PubMed]
- von Söhsten Lins, E.M.D.; Bittar, R.S.M.; Bazán, P.R.; Júnior, E.A.; Staab, J.P. Cerebral responses to stationary emotional stimuli measured by fMRI in women with persistent postural-perceptual dizziness. Int. Arch. Otorhinolaryngol. 2021, 25, e355–e364. [Google Scholar] [CrossRef] [PubMed]
- Indovina, I.; Bosco, G.; Riccelli, R.; Maffei, V.; Lacquaniti, F.; Passamonti, L.; Toschi, N. Structural connectome and connectivity lateralization of the multimodal vestibular cortical network. NeuroImage 2020, 222, 117247. [Google Scholar] [CrossRef]
- Indovina, I.; Maffei, V.; Mazzarella, E.; Sulpizio, V.; Galati, G.; Lacquaniti, F. Path integration in 3D from visual motion cues: A human fMRI study. NeuroImage 2016, 142, 512–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indovina, I.; Maffei, V.; Pauwels, K.; Macaluso, E.; Orban, G.A.; Lacquaniti, F. Simulated self-motion in a visual gravity field: Sensitivity to vertical and horizontal heading in the human brain. NeuroImage 2013, 71, 114–124. [Google Scholar] [CrossRef]
- Indovina, I.; Maffei, V.; Bosco, G.; Zago, M.; Macaluso, E.; Lacquaniti, F. Representation of visual gravitational motion in the human vestibular cortex. Science 2005, 308, 416–419. [Google Scholar] [CrossRef] [Green Version]
- Maffei, V.; Mazzarella, E.; Piras, F.; Spalletta, G.; Caltagirone, C.; Lacquaniti, F.; Daprati, E. Processing of visual gravitational motion in the peri-sylvian cortex: Evidence from brain-damaged patients. Cortex 2016, 78, 55–69. [Google Scholar] [CrossRef]
- Maffei, V.; Macaluso, E.; Indovina, I.; Orban, G.; Lacquaniti, F. Processing of targets in smooth or apparent motion along the vertical in the human brain: An fMRI study. J. Neurophysiol. 2010, 103, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.L.; Maffei, V.; Bosco, G.; Iosa, M.; Zago, M.; Macaluso, E.; Lacquaniti, F. Vestibular nuclei and cerebellum put visual gravitational motion in context. J. Neurophysiol. 2008, 99, 1969–1982. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, C.; Fautrelle, L.; Papaxanthis, C.; Fadiga, L.; Pozzo, T.; White, O. Direction-dependent activation of the insular cortex during vertical and horizontal hand movements. Neuroscience 2016, 325, 10–19. [Google Scholar] [CrossRef]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, E.Y.K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A multi-modal parcellation of human cerebral cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Van Essen, D.C. A tension-based theory of morphogenesis and compact wiring in the central nervous system. Nature 1997, 385, 313–318. [Google Scholar] [CrossRef]
- Von Economo, C.; Koskinas, G. Atlas of Cytoarchitectonics of the Adult Human Cerebral Cortex; Karger: Basel, Switzerland, 2008. [Google Scholar]
- Frank, S.M.; Greenlee, M.W. The parieto-insular vestibular cortex in humans: More than a single area? J. Neurophysiol. 2018, 120, 1438–1450. [Google Scholar] [CrossRef] [Green Version]
- Zu Eulenburg, P.; Stoeter, P.; Dieterich, M. Voxel-based morphometry depicts central compensation after vestibular neuritis. Ann. Neurol. 2010, 68, 241–249. [Google Scholar] [CrossRef]
- Christoffel, D.; Golden, S.; Russo, S.J. Structural and synaptic plasticity in stress-related disorders. Rev. Neurosci. 2011, 22, 535–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S.; Eiland, L.; Hunter, R.; Miller, M.M. Stress and anxiety: Structural plasticity and epigenetic regulation as a consequence of stress. Neuropharmacology 2012, 62, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Dusi, N.; Barlati, S.; Vita, A.; Brambilla, P. Brain structural effects of antidepressant treatment in major depression. Curr. Neuropharmacol. 2015, 13, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Indovina, I.; Maffei, V.; Lacquaniti, F. Anticipating the effects of visual gravity during simulated self-motion: Estimates of time-to-passage along vertical and horizontal paths. Exp. Brain Res. 2013, 229, 579–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestrucci, P.; Daprati, E.; Lacquaniti, F.; Maffei, V. Effects of visual motion consistent or inconsistent with gravity on postural sway. Exp. Brain Res. 2017, 235, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Palomero-Gallagher, N.; Eickhoff, S.B.; Hoffstaedter, F.; Schleicher, A.; Mohlberg, H.; Vogt, B.A.; Amunts, K.; Zilles, K. Functional organization of human subgenual cortical areas: Relationship between architectonical segregation and connectional heterogeneity. NeuroImage 2015, 115, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drevets, W.C.; Savitz, J.; Trimble, M. The subgenual anterior cingulate cortex in mood disorders. CNS Spectrums 2008, 13, 663–681. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.; Morrison, I.; Roberts, N. Role for human posterior parietal cortex in visual processing of aversive objects in peripersonal space. J. Neurophysiol. 2006, 95, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Indovina, I.; Macaluso, E. Dissociation of stimulus relevance and saliency factors during shifts of visuospatial attention. Cereb. Cortex 2007, 17, 1701–1711. [Google Scholar] [CrossRef] [Green Version]
- Indovina, I.; Macaluso, E. Occipital–parietal interactions during shifts of exogenous visuospatial attention: Trial-dependent changes of effective connectivity. Magn. Reson. Imaging 2004, 22, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Downar, J.; Daskalakis, Z.J. New targets for rTMS in depression: A review of convergent evidence. Brain Stimul. 2013, 6, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Fox, P.T.; Miller, K.L.; Glahn, D.C.; Mackay, C.E.; Filippini, N.; Watkins, K.; Toro, R.; Laird, A.; Beckmann, C.F. Correspondence of the brain’s functional architecture during activation and rest. Proc. Natl. Acad. Sci. USA 2009, 106, 13040–13045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redfern, M.S.; Furman, J.M.; Jacob, R.G. Visually induced postural sway in anxiety disorders. J. Anxiety Disord. 2007, 21, 704–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, B.; Milad, M.R. The study of fear extinction: Implications for anxiety disorders. Am. J. Psychiatry 2011, 168, 1255–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holle, D.; Schulte-Steinberg, B.; Wurthmann, S.; Naegel, S.; Ayzenberg, I.; Diener, H.-C.; Katsarava, Z.; Obermann, M. Persistent postural-perceptual dizziness: A matter of higher, central dysfunction? PLoS ONE 2015, 10, e0142468. [Google Scholar] [CrossRef]
- Gold, A.; Morey, R.A.; McCarthy, G. Amygdala—Prefrontal cortex functional connectivity during threat-induced anxiety and goal distraction. Biol. Psychiatry 2015, 77, 394–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indovina, I.; Riccelli, R.; Staab, J.; Lacquaniti, F.; Passamonti, L. Personality traits modulate subcortical and cortical vestibular and anxiety responses to sound-evoked otolithic receptor stimulation. J. Psychosom. Res. 2014, 77, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Riccelli, R.; Indovina, I.; Staab, J.; Nigro, S.; Augimeri, A.; Lacquaniti, F.; Passamonti, L. Neuroticism modulates brain visuo-vestibular and anxiety systems during a virtual rollercoaster task. Hum. Brain Mapp. 2016, 38, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.J.; Adams, R.; Brown, H.; Pareés, I.; Friston, K. A Bayesian account of ‘hysteria’. Brain 2012, 135, 3495–3512. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Patients | Diagnosis | Trigger a | Depression Accounted for | Anxiety Accounted for | Protocol b | |
|---|---|---|---|---|---|---|
| Wurthmann et al. [51] | 42 | PPPD | 13 vestibular 29 non-vestibular (unknown) | no | no | grey matter volume |
| Popp et al. [48] | 37 | PPV | non-vestibular (unknown) | no | no | grey matter volume and cortical thickness, visual motion stimulation during fMRI |
| Nigro et al., 2019 [46] Indovina et al., 2015 [39] Riccelli et al. [49] Passamonti et al. [47] | 18 | CSD/PPPD | vestibular | yes | yes | cortical folding sound evoked vestibular stimulation during fMRI visually evoked self-motion simulation during fMRI |
| Van Ombergen et al. [36] | 10 | VID | 9 vestibular 1 non-vestibular (unknown) | no | no | Resting state fMRI |
| Lee et al. [42] | 38 | PPPD | 9 vestibular 4 migraine 1 anxiety 14 non-vestibular (unknown) | yes | yes | Resting state fMRI |
| Li et al., 2020 [43,44] | 12 | PPPD | not available | no | no | Resting state fMRI |
| Na et al. [45] | 25 | PPPD | 6 vestibular 6 emotional stress 2 presyncope 2 sleep deprivation 1 mild head trauma | no | no | Resting state during SPECT |
| Roberts et al. [50] | 17 | VID | vestibular | no | no | Caloric vestibular and visual motion stimulation during fMRI |
| von Söhsten Lins, et al. [54] | 16 | PPPD | Patients with PPPD (N = 16) 5 psychiatric disorder 4 metabolic disorder 4 peripheral vestibular 2 dysautonomia 1 other Patients recovered from acute vestibular syndromes (N = 16) 8 metabolic disorder 5 peripheral vestibular 2 substance-related 1 other | no | no | Stationary images with positive, neutral, and negative emotional valence during fMRI. Note. Comparison patients had recovered fully from their acute vestibular syndromes by the time they underwent neuroimaging. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Indovina, I.; Passamonti, L.; Mucci, V.; Chiarella, G.; Lacquaniti, F.; Staab, J.P. Brain Correlates of Persistent Postural-Perceptual Dizziness: A Review of Neuroimaging Studies. J. Clin. Med. 2021, 10, 4274. https://doi.org/10.3390/jcm10184274
Indovina I, Passamonti L, Mucci V, Chiarella G, Lacquaniti F, Staab JP. Brain Correlates of Persistent Postural-Perceptual Dizziness: A Review of Neuroimaging Studies. Journal of Clinical Medicine. 2021; 10(18):4274. https://doi.org/10.3390/jcm10184274
Chicago/Turabian StyleIndovina, Iole, Luca Passamonti, Viviana Mucci, Giuseppe Chiarella, Francesco Lacquaniti, and Jeffrey P. Staab. 2021. "Brain Correlates of Persistent Postural-Perceptual Dizziness: A Review of Neuroimaging Studies" Journal of Clinical Medicine 10, no. 18: 4274. https://doi.org/10.3390/jcm10184274