
Epidermal-Derived Hedgehog Signaling Drives Mesenchymal Proliferation during Digit Tip Regeneration

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Surgeries
2.2. Tamoxifen Injections
2.3. Digit Tip Amputation
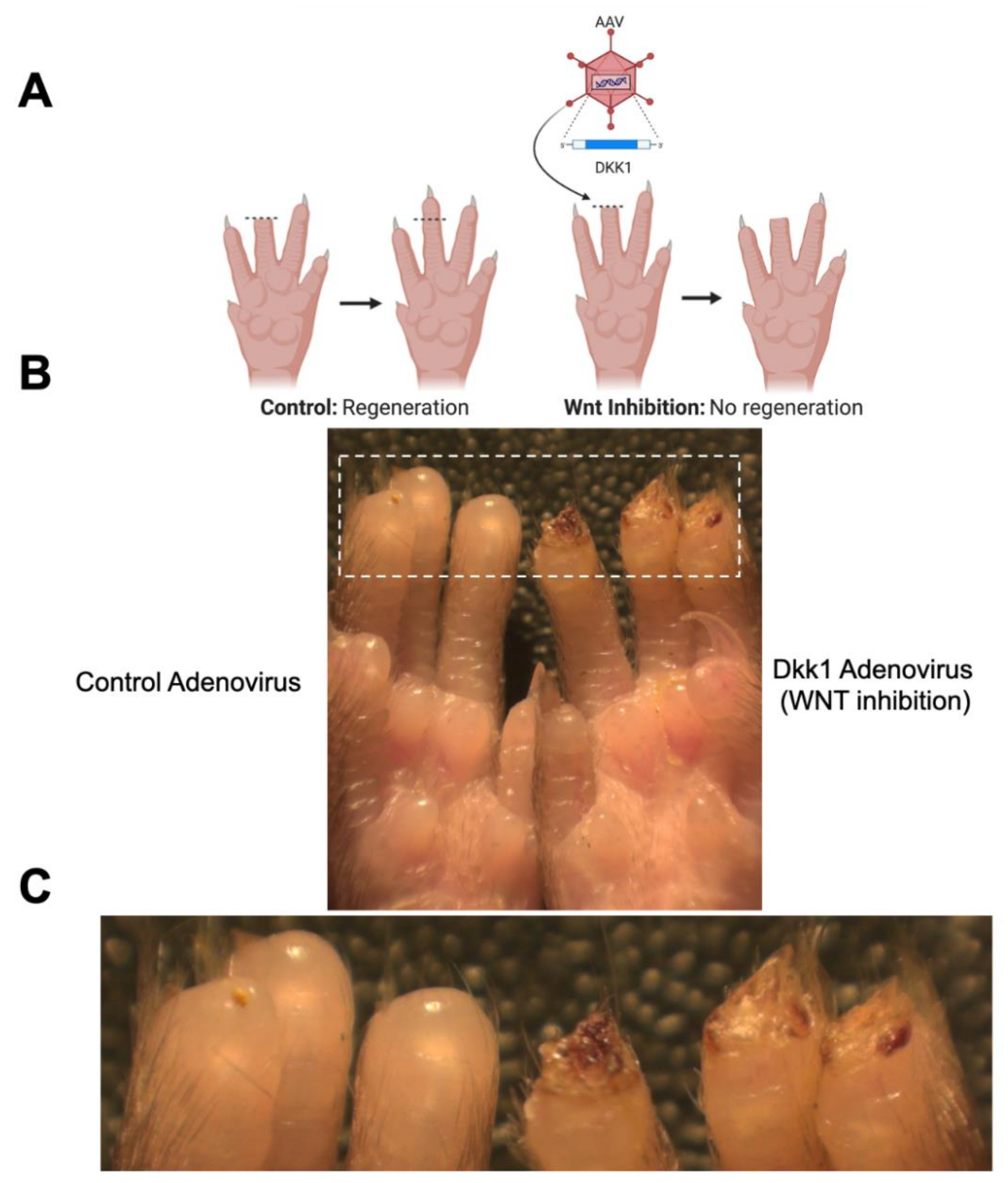
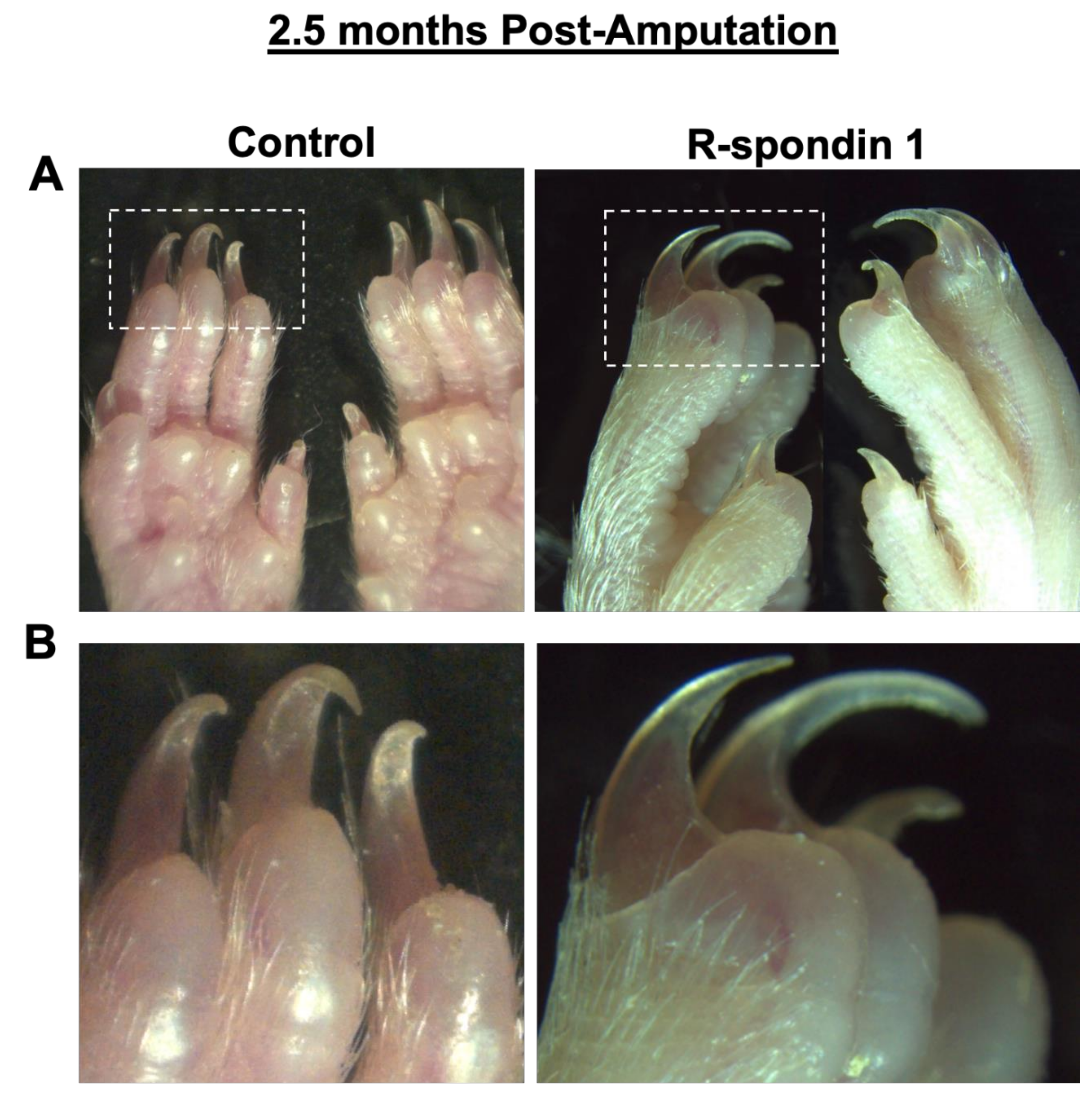
2.4. Adenovirus Treatment
2.5. Histology
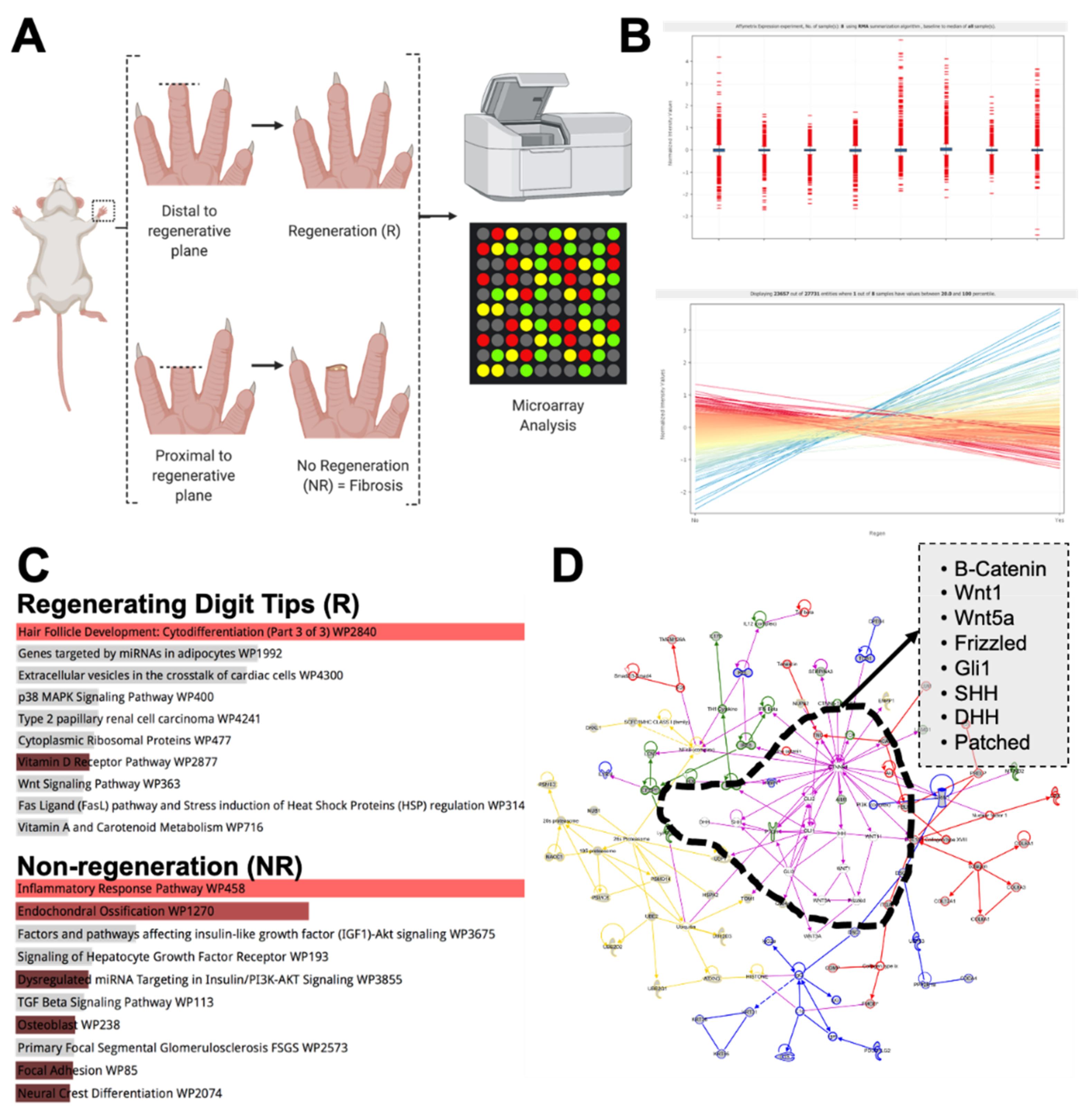
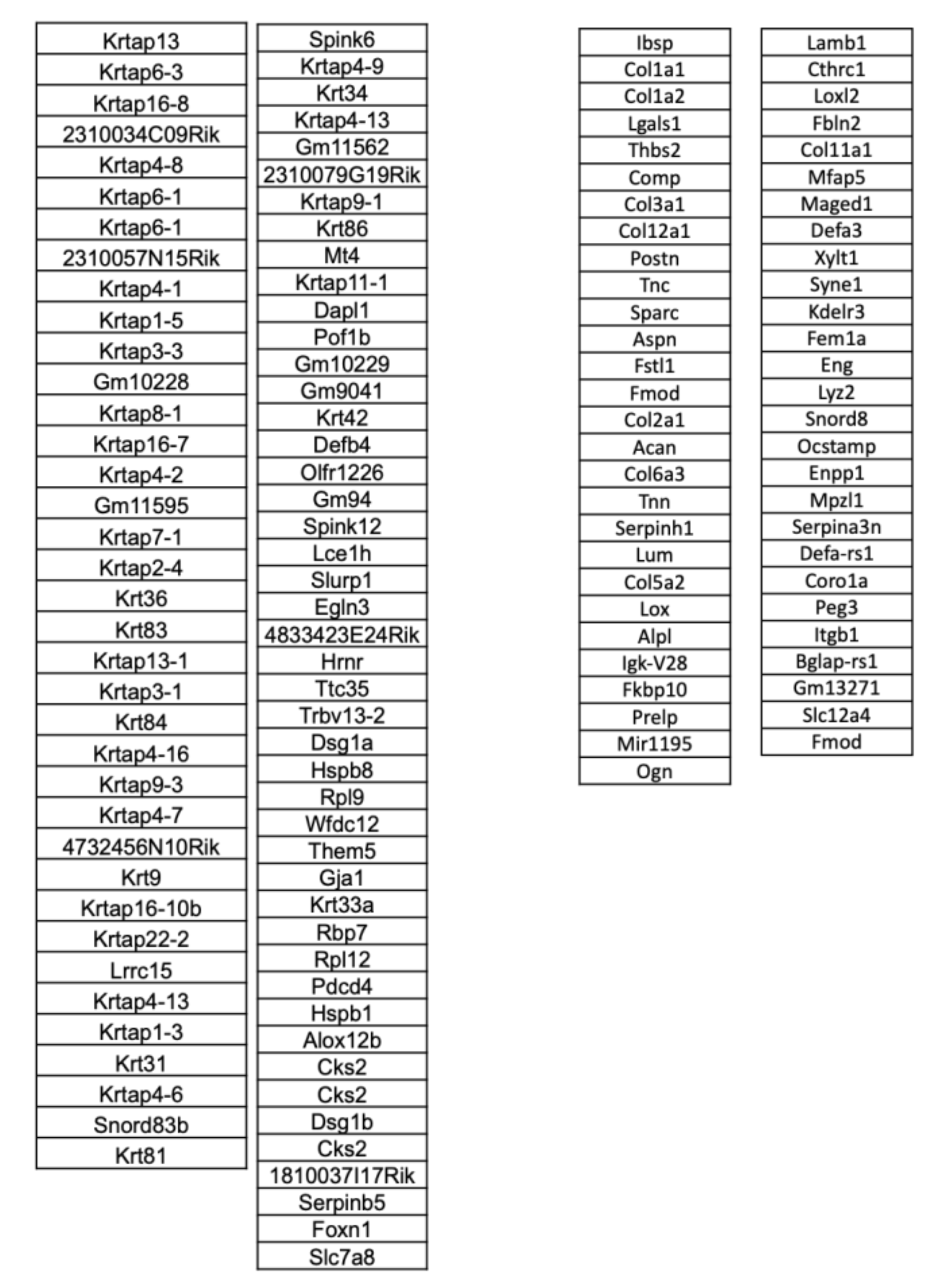
2.6. Microarray Analysis
2.7. Cell Culture
2.8. Imaging and Analysis
2.9. Statistics
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, T.J.; Deptula, P.L.; Buncke, G.M.; Maan, Z.N. Digit Tip Injuries: Current Treatment and Future Regenerative Paradigms. Stem Cells Int. 2019, 2019, 9619080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melvin, J.L. Roles and Functions of Occupational Therapy in Hand Rehabilitation. Am. J. Occup. Ther. 1985, 39, 795–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldfarb, C.A.; Lee, W.P.A.; Briskey, D.; Higgins, J.P. An American Society for Surgery of the Hand (ASSH) Task Force Report on Hand Surgery Subspecialty Certification and ASSH Membership. J. Hand Surg. 2014, 39, 330–334. [Google Scholar] [CrossRef]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nat. Cell Biol. 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Padmanabhan, J. Disease models: Method in the madness of fibrosis. Nat. Mater. 2017, 16, 1176–1177. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, K.M.C.; Barker, J.H.; Berezikov, E.; Pindur, L.; Kynigopoulos, S.; Eischen-Loges, M.; Han, Z.; Bhavsar, M.B.; Henrich, D.; Leppik, L. Electrical stimulation shifts healing/scarring towards regeneration in a rat limb amputation model. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Illingworth, C.M. Trapped fingers and amputated finger tips in children. J. Pediatr. Surg. 1974, 9, 853–858. [Google Scholar] [CrossRef]
- Douglas, B.S. Conservative Management of Guillotine Amputation of the Finger in Children. J. Paediatr. Child Health 1972, 8, 86–89. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Lindau, P.; Ueno, H.; Longaker, M.T.; Weissman, I.L. Germ-layer and lineage-restricted stem/progenitors regenerate the mouse digit tip. Nat. Cell Biol. 2011, 476, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Borgens, R. Mice regrow the tips of their foretoes. Science 1982, 217, 747–750. [Google Scholar] [CrossRef]
- Lehoczky, J.A.; Robert, B.; Tabin, C.J. Mouse digit tip regeneration is mediated by fate-restricted progenitor cells. Proc. Natl. Acad. Sci. USA 2011, 108, 20609–20614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, G.L.; Masias, E.J.; Lehoczky, J.A. Cellular Heterogeneity and Lineage Restriction during Mouse Digit Tip Regeneration at Single-Cell Resolution. Dev. Cell 2020, 52, 525–540.e5. [Google Scholar] [CrossRef]
- Storer, M.; Mahmud, N.; Karamboulas, K.; Borrett, M.J.; Yuzwa, S.A.; Gont, A.; Androschuk, A.; Sefton, M.V.; Kaplan, D.R.; Miller, F.D. Acquisition of a Unique Mesenchymal Precursor-like Blastema State Underlies Successful Adult Mammalian Digit Tip Regeneration. Dev. Cell 2020, 52, 509–524.e9. [Google Scholar] [CrossRef]
- Lehoczky, J.A.; Tabin, C.J. Lgr6 marks nail stem cells and is required for digit tip regeneration. Proc. Natl. Acad. Sci. USA 2015, 112, 13249–13254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeo, M.; Chou, W.C.; Sun, Q.; Lee, W.; Rabbani, P.; Loomis, C.; Taketo, M.M.; Ito, M. Wnt activation in nail epithelium couples nail growth to digit regeneration. Nat. Cell Biol. 2013, 499, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, Y.; Kandyba, E.; Chen, Y.-B.; Ruffins, S.; Chuong, C.-M.; Kobielak, K. Bifunctional ectodermal stem cells around the nail display dual fate homeostasis and adaptive wounding response toward nail regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 15114–15119. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, D.A.; Zhao, W. Bone regrowth after digit tip amputation in mice is equivalent in adults and neonates. Wound Repair Regen. 1995, 3, 461–466. [Google Scholar] [CrossRef]
- Chamberlain, C.S.; Jeffery, J.J.; Leiferman, E.M.; Yildirim, T.; Sun, X.; Baer, G.S.; Murphy, W.L.; Vanderby, R. Level-specific amputations and resulting regenerative outcomes in the mouse distal phalanx. Wound Repair Regen. 2017, 25, 443–453. [Google Scholar] [CrossRef]
- Dawson, L.A.; Schanes, P.P.; Marrero, L.; Jordan, K.; Brunauer, R.; Zimmel, K.N.; Qureshi, O.; Imholt, F.M.; Falck, A.R.; Yan, M.; et al. Proximal digit tip amputation initiates simultaneous blastema and transient fibrosis formation and results in partial regeneration. Wound Repair Regen. 2021, 29, 196–205. [Google Scholar] [CrossRef]
- Quijano, L.M.; Lynch, K.M.; Allan, C.H.; Badylak, S.F.; Ahsan, T. Looking Ahead to Engineering Epimorphic Regeneration of a Human Digit or Limb. Tissue Eng. Part B Rev. 2016, 22, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Wong, V.W.; Paterno, J.; Sorkin, M.; Glotzbach, J.P.; Levi, K.; Januszyk, M.; Rustad, K.C.; Longaker, M.T.; Gurtner, G.C. Mechanical force prolongs acute inflammation via T-cell-dependent pathways during scar formation. FASEB J. 2011, 25, 4498–4510. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, G.G.; Maan, Z.N.; Hu, M.S.; Atashroo, D.A.; Whittam, A.J.; Duscher, D.; Tevlin, R.; Marecic, O.; Lorenz, H.P.; Gurtner, G.C.; et al. Murine Dermal Fibroblast Isolation by FACS. J. Vis. Exp. 2016, e53430. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Maldonado, E.; Vasconcelos, V.; O’Brien, S.J.; Johnson, W.; Antunes, A. Mammalian keratin associated proteins (KRTAPs) subgenomes: Disentangling hair diversity and adaptation to terrestrial and aquatic environments. BMC Genom. 2014, 15, 779. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, H.; Fujimoto, A.; Farooq, M.; Ito, M.; Shimomura, Y. Characterization of the Human Hair Keratin–Associated Protein 2 (KRTAP2) Gene Family. J. Investig. Dermatol. 2012, 132, 1806–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.-Y.; Klar, J.; Georgii-Heming, P.; Fröjmark, A.-S.; Baig, S.M.; Schlessinger, D.; Dahl, N. Frizzled6 deficiency disrupts the differentiation process of nail development. J. Investig. Dermatol. 2013, 133, 1990–1997. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Takeo, M.; Hale, C.S.; Ito, M. Epithelium-Derived Wnt Ligands Are Essential for Maintenance of Underlying Digit Bone. J. Investig. Dermatol. 2016, 136, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.H.; Sun, Q.; Ratti, K.; Lee, S.-H.; Zheng, Y.; Takeo, M.; Lee, W.; Rabbani, P.; Plikus, M.V.; Cain, J.E.; et al. Hedgehog stimulates hair follicle neogenesis by creating inductive dermis during murine skin wound healing. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Soikkeli, J.; Podlasz, P.; Yin, M.; Nummela, P.; Jahkola, T.; Virolainen, S.; Krogerus, L.; Heikkilä, P.; von Smitten, K.; Saksela, O.; et al. Metastatic Outgrowth Encompasses COL-I, FN1, and POSTN Up-Regulation and Assembly to Fibrillar Networks Regulating Cell Adhesion, Migration, and Growth. Am. J. Pathol. 2010, 177, 387–403. [Google Scholar] [CrossRef]
- Karlsen, T.A.; Jakobsen, R.B.; Mikkelsen, T.S.; Brinchmann, J.E. microRNA-140 Targets RALA and Regulates Chondrogenic Differentiation of Human Mesenchymal Stem Cells by Translational Enhancement of SOX9 and ACAN. Stem Cells Dev. 2014, 23, 290–304. [Google Scholar] [CrossRef] [Green Version]
- Nomura, M.; Liu, J.; Rovira, I.; Gonzalez-Hurtado, E.; Lee, J.; Wolfgang, M.J.; Finkel, T. Fatty acid oxidation in macrophage polarization. Nat. Immunol. 2016, 17, 216–217. [Google Scholar] [CrossRef]
- Wong, V.W.; Rustad, K.C.; Akaishi, S.; Sorkin, M.; Glotzbach, J.; Januszyk, M.; Nelson, E.R.; Levi, K.; Paterno, J.; Vial, I.N.; et al. Focal adhesion kinase links mechanical force to skin fibrosis via inflammatory signaling. Nat. Med. 2012, 18, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Carver, W.; Goldsmith, E.C. Regulation of Tissue Fibrosis by the Biomechanical Environment. BioMed Res. Int. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarabi, S.; Bhatt, K.A.; Shi, Y.; Paterno, J.; Chang, E.I.; Loh, S.A.; Holmes, J.; Longaker, M.T.; Yee, H.; Gurtner, G.C. Mechanical load initiates hypertrophic scar formation through decreased cellular apoptosis. FASEB J. 2007, 21, 3250–3261. [Google Scholar] [CrossRef] [Green Version]
- Scali, C.; Caraci, F.; Gianfriddo, M.; Diodato, E.; Roncarati, R.; Pollio, G.; Gaviraghi, G.; Copani, A.; Nicoletti, F.; Terstappen, G.C.; et al. Inhibition of Wnt signaling, modulation of Tau phosphorylation and induction of neuronal cell death by DKK1. Neurobiol. Dis. 2006, 24, 254–265. [Google Scholar] [CrossRef] [PubMed]
- De Lau, W.; Peng, W.C.; Gros, P.; Clevers, H. The R-spondin/Lgr5/Rnf43 module: Regulator of Wnt signal strength. Genes Dev. 2014, 28, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepers, A.G.; Snippert, H.J.; Stange, D.E.; Born, M.V.D.; van Es, J.H.; van de Wetering, M.; Clevers, H. Lineage Tracing Reveals Lgr5+ Stem Cell Activity in Mouse Intestinal Adenomas. Science 2012, 337, 730–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Poss, K.D. Clonally dominant cardiomyocytes direct heart morphogenesis. Nat. Cell Biol. 2012, 484, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Li, Y.; Liu, Z.; Zhang, J.; Cui, Y.; Chen, X.; Chopp, M. The Sonic Hedgehog Pathway Mediates Brain Plasticity and Subsequent Functional Recovery After Bone Marrow Stromal Cell Treatment of Stroke in Mice. J. Cereb. Blood Flow Metab. 2013, 33, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Galiano, R.D.; Michaels, V.J.; Dobryansky, M.; Levine, J.P.; Gurtner, G.C. Quantitative and reproducible murine model of excisional wound healing. Wound Repair Regen. 2004, 12, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Ang, S.; Conlon, R.; Jin, O.; Rossant, J. Positive and negative signals from mesoderm regulate the expression of mouse Otx2 in ectoderm explants. Development 1994, 120, 2979–2989. [Google Scholar] [CrossRef] [PubMed]
- Mariani, F.V.; Fernandez-Teran, M.; Ros, M.A. Ectoderm-mesoderm crosstalk in the embryonic limb: The role of fibroblast growth factor signaling. Dev. Dyn. 2017, 246, 208–216. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maan, Z.N.; Rinkevich, Y.; Barrera, J.; Chen, K.; Henn, D.; Foster, D.; Bonham, C.A.; Padmanabhan, J.; Sivaraj, D.; Duscher, D.; et al. Epidermal-Derived Hedgehog Signaling Drives Mesenchymal Proliferation during Digit Tip Regeneration. J. Clin. Med. 2021, 10, 4261. https://doi.org/10.3390/jcm10184261
Maan ZN, Rinkevich Y, Barrera J, Chen K, Henn D, Foster D, Bonham CA, Padmanabhan J, Sivaraj D, Duscher D, et al. Epidermal-Derived Hedgehog Signaling Drives Mesenchymal Proliferation during Digit Tip Regeneration. Journal of Clinical Medicine. 2021; 10(18):4261. https://doi.org/10.3390/jcm10184261
Chicago/Turabian StyleMaan, Zeshaan N., Yuval Rinkevich, Janos Barrera, Kellen Chen, Dominic Henn, Deshka Foster, Clark Andrew Bonham, Jagannath Padmanabhan, Dharshan Sivaraj, Dominik Duscher, and et al. 2021. "Epidermal-Derived Hedgehog Signaling Drives Mesenchymal Proliferation during Digit Tip Regeneration" Journal of Clinical Medicine 10, no. 18: 4261. https://doi.org/10.3390/jcm10184261