1. Introduction
Technological advances in mass sequencing have enabled the identification of different microbial communities in the uterine cavity, including in the vagina and the endometrial cavity [
1]. A recent review [
2] concluded that there was not enough evidence for a “core” or bacterial resident population in the uterus, and the existence of uterine microbiota might be reflective of bacterial tourists or invaders. Nevertheless, uterine microbiota may be involved in reproductive health and disease [
1,
2]
The main bacteria at the vaginal and endometrial level belong to the genus
Lactobacillus—producers of lactic acid that maintain the acidic pH of the vagina, which acts as a barrier against pathogens [
3]. The association between vaginal flora and pregnancy outcomes has been widely studied for years. The live birth rate is correlated with the production of H
2O
2 by
Lactobacillus spp., and inversely correlated with the existence of bacterial vaginosis. Thus, alterations to the vaginal flora—for example, due to bacterial vaginosis (provoked by
Gardnerella vaginalis)—are associated with an increased risk of miscarriage [
4,
5]. Other pathogenic microorganisms—such as
Chlamydia trachomatis,
Neisseria gonorrhoeae, and
Mycoplasma tuberculosis—may cause subclinical alterations related to risk factors for subfertility [
6].
The prevalence of infertility is on the rise [
7], and assisted reproductive technologies (ART) are increasingly in demand, as well as being safer and more successful [
8]. Endometrial implantation is the single most important event determining the success of embryo transfer in ART [
9]. Other factors include the microbial colonization of the upper genital tract [
10]—and possibly the uterine–cervical canal—which has been shown to be a significant, independent determinant of the success of assisted reproductive treatments [
11]. Recently, the reproductive tract microbiota have been associated with embryo transfer failure [
12].
Until now, the reproductive microbiome has been studied by analyzing samples taken from either the endometrium or the vagina. Although the uterine microbiota appear to be a continuum from the vaginal microbiota, previous studies have found differences between the endometrial and vaginal microbiota [
13]. Currently, the role of these microbiota in embryo implantation and pregnancy outcomes is unclear. One of the main obstacles to determining the bacterial composition of the endometrium is that the small amount of the initial sample makes it vulnerable to contamination with exogenous bacterial DNA.
Recently, Di Simone et al. [
14] suggested that classification based on endometrial bacterial patterns could help prevent obstetric complications through personalized treatments. In light of scientific evidence that alterations to the reproductive microbiome reduce women’s fertility by negatively impacting on embryo implantation, determining the bacterial composition of the vagina and the endometrium may contribute to improving the prognosis of fertility treatments. Thus, the primary objective of the present study was to describe and compare the vaginal and endometrial microbiome patterns between women who became pregnant and women who did not after in vitro fertilization (IVF). The secondary objective was to compare the vaginal and endometrial microbiome patterns between women with and without a history of repeated implantation failures (RIF).
2. Materials and Methods
2.1. Design and Study Population
This pilot prospective cohort study took place in a private assisted reproduction clinic. It included women presenting to the fertility clinic for IVF from May 2017 to May 2019 who met the following inclusion criteria: aged 18–50 years; undergoing frozen embryo transfer (FET) with euploid embryos, using either their own or donated gametes; and an indication to use the intracytoplasmic sperm injection (ICSI) method to generate embryos. Exclusion criteria were: the use of antibiotics in the three months preceding the fertility treatment, uterine malformations, untreated hydrosalpinx, known implantation failure factors, or unwillingness to sign informed consent.
Participants followed the usual ART procedure with ovarian stimulation and ICSI per protocol, and underwent preimplantation genetic testing for aneuploidies (PGT-A) at the blastocyst stage using NGS (VeriSeq, Illumina,, San Diego, California, USA) with the MiSeq Sequencer (Illumina). One euploid embryo was transferred in the cycle following the ovarian stimulation via ultrasound-guided transfer, in accordance with established protocols. Endometrial preparation via estrogen–progesterone replacement therapy was performed. Human chorionic gonadotropin (β-hCG) blood testing was performed 8–9 days after the embryo transfer to test for pregnancy.
The local ethics committee approved this study in January 2017 (Reference code: 16/318), and it was conducted in accordance with the Declaration of Helsinki. All participants signed written informed consent to take part in the study.
2.2. Study Variables
The following baseline characteristics were collected on enrollment: age (years), weight (kg), tobacco use (yes/no), number of previous pregnancies, previous miscarriages (yes/no), number of miscarriages, previous fertility treatments (yes/no), RIF, i.e., failed implantation after ≥ 3 transfer cycles with good-quality embryos (yes/no), donated oocytes (yes/no), donated semen (yes/no), and normozoospermia diagnosis after semen analysis (yes/no). The date and the endometrial thickness (mm) were recorded before the embryo transfer.
The primary study outcome was clinical pregnancy (yes/no), i.e., pregnancy evidenced by ultrasound of fetal cardiac activity after positive β-hCG. Other reproductive outcomes included the result of the β-hCG blood test (positive/negative), biochemical miscarriage (yes/no), and clinical miscarriage (yes/no).
2.3. Sample Collection
Vaginal samples were collected at different stages of IVF treatment: (1) during the secretory phase of the cycle previous to FET (days 18–22 of the cycle), (2) on the day of the embryo transfer, and (3) on the day of the pregnancy test. A dry swab was used to collect vaginal fluid from the bottom of the posterior sac by direct visualization using the vaginal speculum, with the patient in the lithotomy position. In order to avoid contamination, we did not use lubricant or gel on the speculum. On the day of the embryo transfer, we took the sample before preparing the embryo transfer to prevent interference by the procedure.
Endometrial samples were collected during the secretory phase of the cycle previous to FET (days 18–22 of the cycle), and we used the Tao Brush IUMC Endometrial Sampler; this device minimizes the risk of contamination during the collection of the endometrial sample via a sheath that closes prior to withdrawal from the uterus. All vaginal and endometrial samples were stored at −80 °C until further analysis.
2.4. Sample Analysis
Microbiome patterns were analyzed by estimating the prevalence and variability of types of bacteria at both the vaginal and endometrial levels. We used metagenomics for sample analysis, studying the 16S rRNA gene marker of the included samples via next-generation sequencing (NGS). Analyses took place in the molecular genetics laboratory of the fertility clinic. Rigorous controls were carried out for the reagents and all of the equipment used in all of the steps carried out during the processing and analysis of the samples.
2.5. DNA Extraction
DNA extraction was performed using the PureLink Microbiome DNA Purification Kit (Thermo Fisher, PureLinkTM Microbiome DNA Purification Kit, Darmstadt, Germany and/or its affiliates). The DNA was quantified using a Qubit 2.0 Fluorometer (Thermo Fisher). The extracted DNA was stored at −20 °C for later use.
2.6. Amplification of Region V3V4 of the 16S rRNA Gene
Polymerase chain reaction (PCR) amplification of the variable region V3V4 of the 16S rRNA gene was performed using Taq DNA polymerase (2× KAPA HiFi HotStart, Roche, Rotkreuz, Switzerland) in the presence of dNTPs, as well as oligonucleotides 357F and 806R, at a final concentration of 1 μM and an average of 100 ng of DNA, and at a final reaction volume of 25 μL, following the recommendations of Illumina (16S Metagenomic Sequencing Library Preparation). PCR was carried out in a thermal cycler (Verity, Applied Biosystems, San Francisco, CA, USA). For the validation of the PCR technique, all amplification reactions included positive and negative controls without DNA templates. The PCR products were visualized using agarose electrophoresis, verifying that the amplified DNA band was the correct size (449 base pairs). All products of amplification were stored at −20 °C for subsequent sequencing.
2.7. Sequencing of Region V3V4 of 16S rRNA Gene
Once the V3V4 amplicon was obtained and purified, we generated the library with the identifying indices of each sample using the Nextera XT sequencing kit (Illumina). After the purification of the libraries, the samples, which were previously diluted to a concentration of 4 nM before being mixed and prepared for sequencing, were quantified using a Qubit 2.0 Fluorometer (Thermo Fisher). The final concentration of the library was 15 pM. The library was sequenced using MiSeq Reagent Kit v3 (Illumina) reagents. We used MiSeq (Illumina) as the sequencing equipment and for the metagenomics of the workflow.
2.8. Bioinformatic Analysis of the Sequences
The primary analysis of the obtained sequences consisted of demultiplexing, using the MiSeq Reporter software (Illumina). The unindexed paired-end sequences of each sample were exported from the MiSeq system in FASTQ format for the analysis. The bioinformatic analysis of the sequences was carried out using the QIIME2 package. Further data analysis was also performed with MicrobiomeAnalyst software. Deblur was used to filter and denoise the sequences with QIIME2. The sequences were grouped in operational taxonomic units (OTUs) with a similarity percentage of 97%.
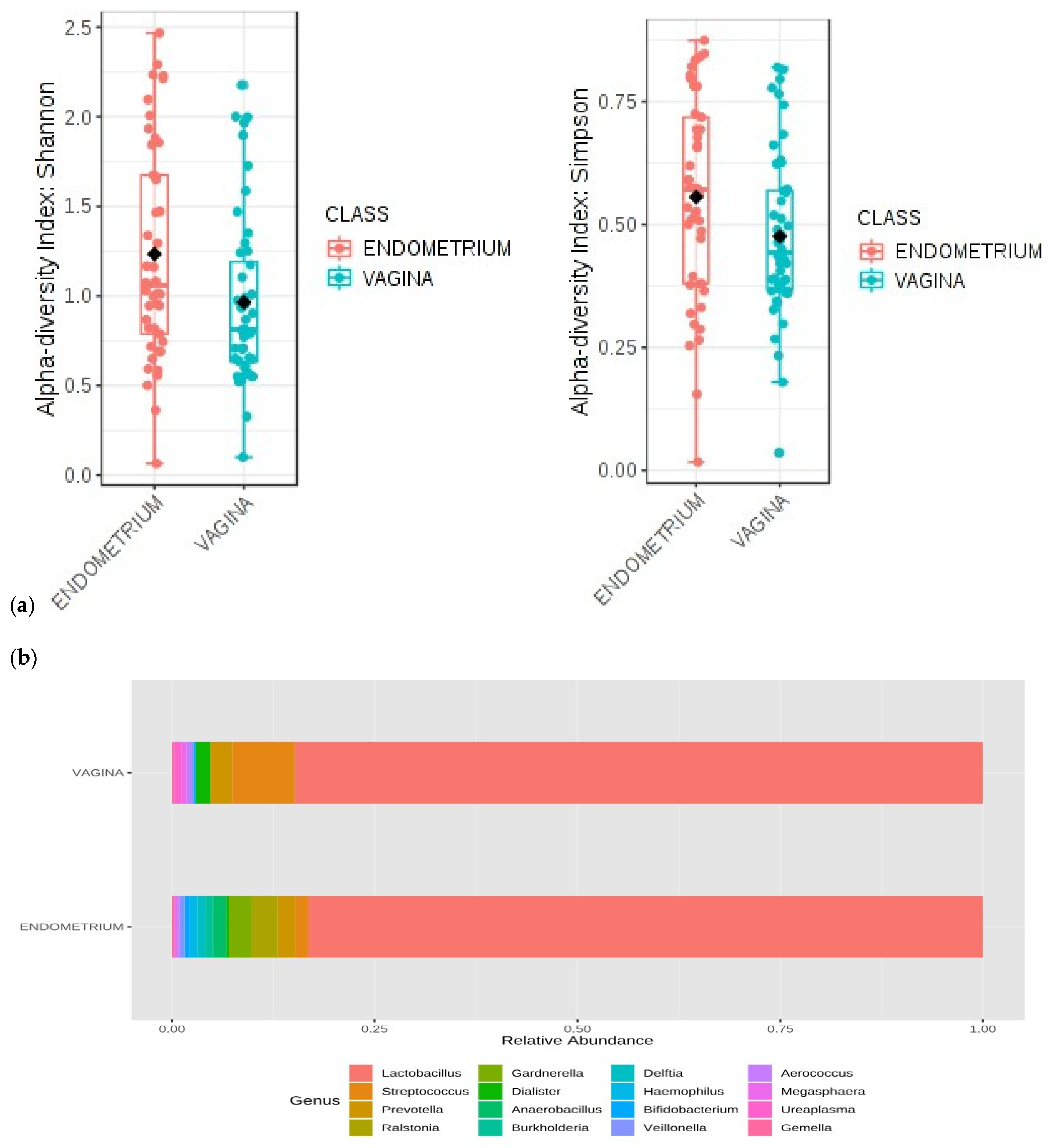
In order to estimate alpha diversity—i.e., the number of different species present in the sample—a rarefaction analysis was performed on 1000 sequences per sample, followed by an alpha diversity analysis. To aid in comparisons of alpha diversity, differences in library sizes across samples were adjusted by the rarefaction method. Three different indices were used: Shannon, Simpson, and Faith. Since these indices did not follow a normal distribution, the non-parametric Mann–Whitney U method was used. The Shannon index quantifies the different types of taxa present in the community, and considers the richness and equitability of species. If two sites are equally rich in species, however, with one site dominated by a single species and the other showing more evenness, the second would clearly be considered more diverse. The Simpson index expresses the probability that two microorganisms randomly selected from an infinitely large community are of different species.
The results for beta diversity were visualized with QIIME2 using the graphics generated by principal coordinates analysis (PCoA), obtained with EMPeror. We carried out the analysis of beta diversity using the unweighted UniFrac index. Beta diversity expresses composition in terms of the abundance of different taxa among the samples. UniFrac is a measure of beta diversity that uses phylogenetic information to compare samples belonging to the interest groups—in this case, four; the unweighted version is qualitative. Therefore, UniFrac measures concordance based on the abundance of OTUs in each sample, including phylogenetic distances. The matrices with beta diversity measurements were analyzed using PERMANOVA for differences in composition according to the group they belonged to (type of sample).
Taxonomic assignment was performed using a classification based on a filtering of the 99_otus sequence from the Greengenes database to the V3V4 region. Finally, we performed the univariate analysis of each specified taxon or group according to the results we obtained, using the correction for multiple testing.
2.9. Statistical Analysis
A descriptive analysis of all variables was performed by calculating frequencies for the qualitative parameters and minima, maxima, means, and standard deviations (SD) for the quantitative variables. To compare them, we used the parametric Student’s t-test or the non-parametric Mann–Whitney U test, as appropriate. Microbiome patterns as well as vaginal and endometrial samples were compared between groups according to outcome variables, using double-entry tables for qualitative variables and the chi-squared test. For quantitative variables, Student’s t-test was used to assess the association between microbiome patterns and clinical pregnancy outcomes. A linear mixed-effects model was constructed to determine the evolution of the vaginal microbiome pattern and its association with clinical pregnancy rate and RIF diagnosis. Analyses were performed using R v.3.5.1.
4. Discussion
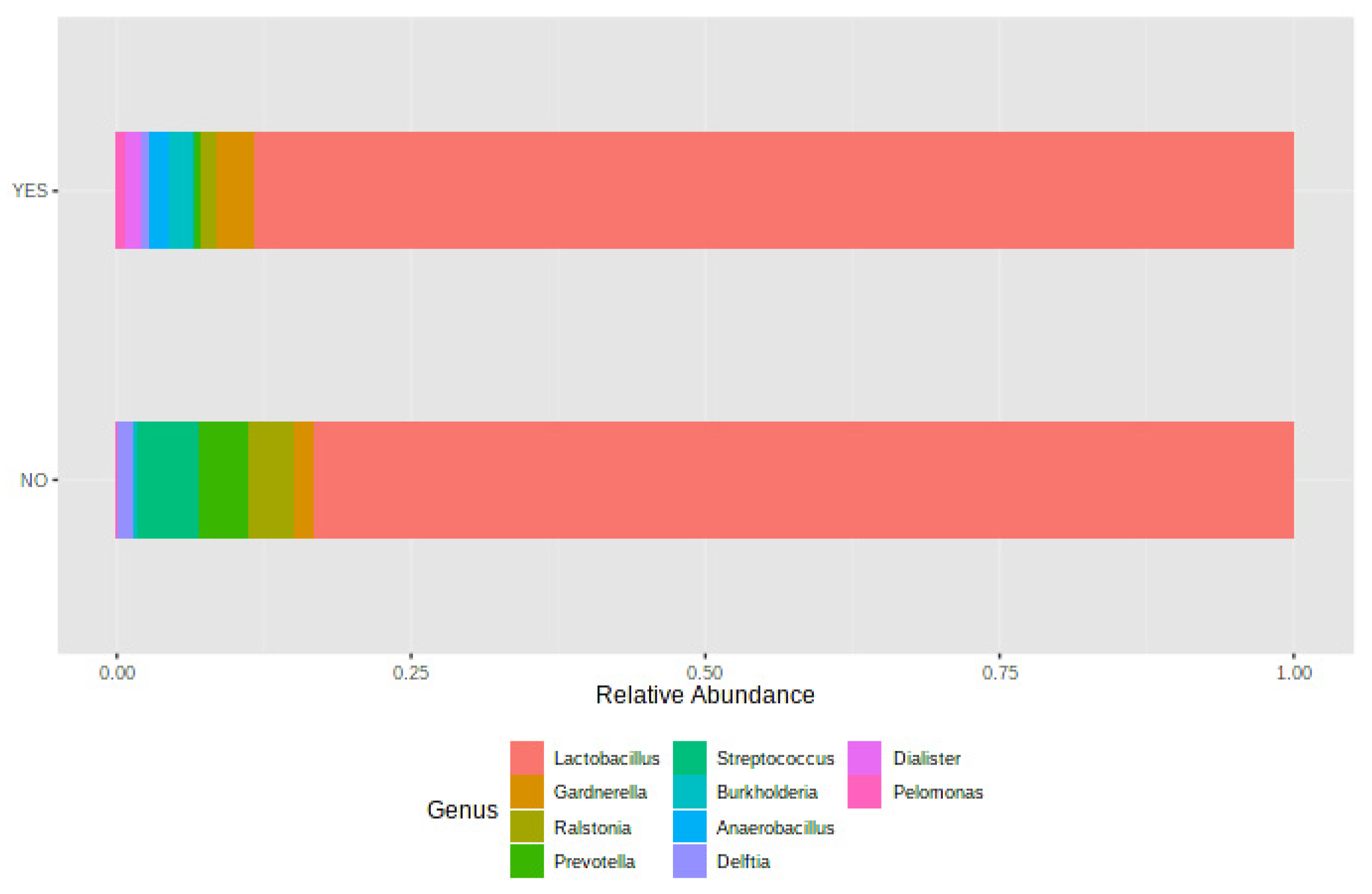
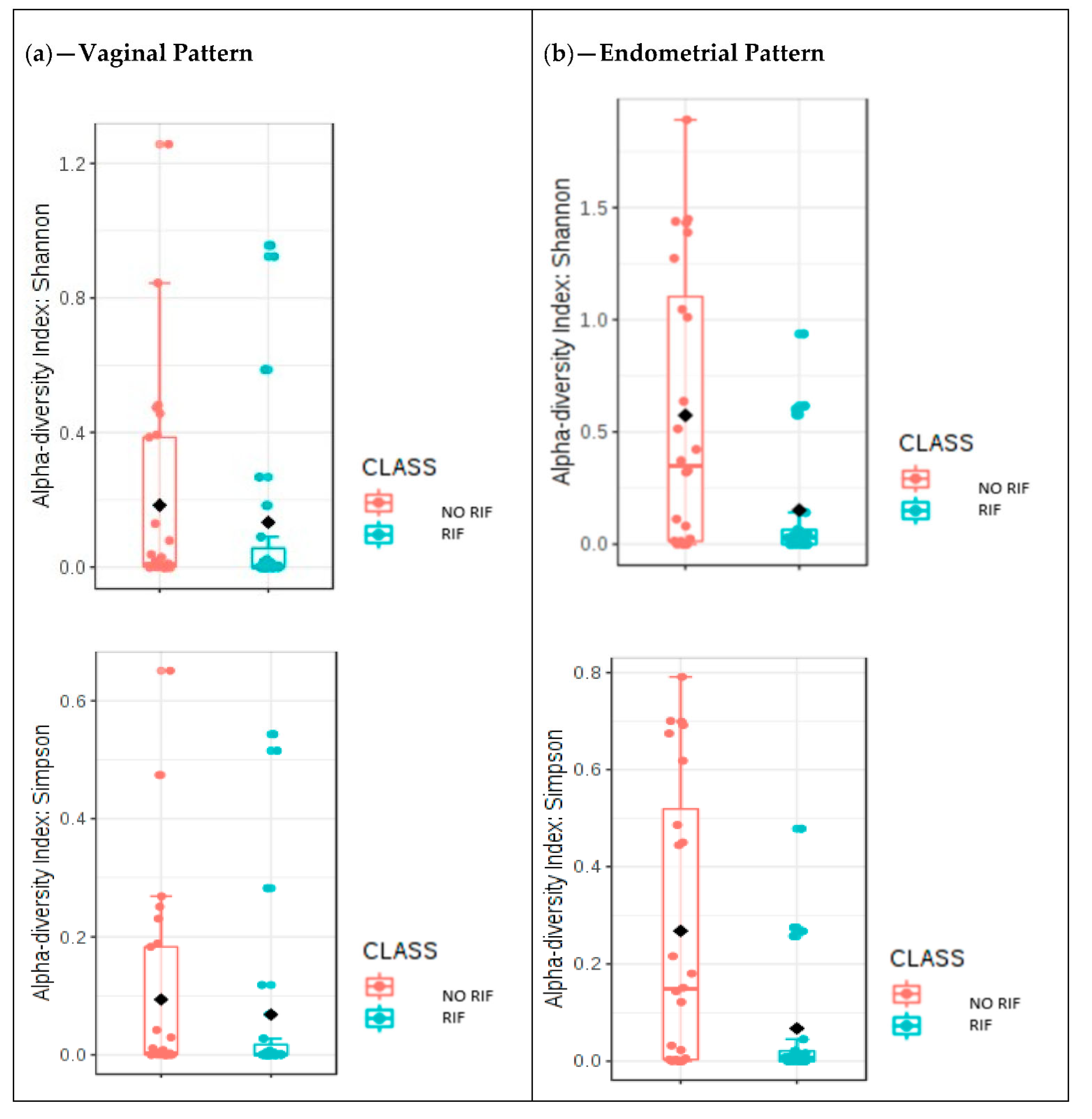
The results of this study show that there were no differences in the diversity of the vaginal microbiome between women who got pregnant and those who did not, but there were differences in its taxonomic characterization, and while statistically significant differences were not observed in the endometrial microbiome patterns, there were statistical differences in the composition of the vaginal versus endometrial microbiomes. Our results also suggest stability of the vaginal microbiome throughout different timepoints of the cycle. On the other hand, the vaginal samples collected during the secretory phase of the cycle previous to FET showed differences only for the Faith index between the women with and without RIF diagnosis, with a higher alpha diversity in women with a history of RIF. At the taxonomic level, the relative abundance of the genera in women with RIF was different from those without RIF. With regard to endometrial microbiome patterns, statistically significant differences were found between women with and without RIF.
Franasiak et al. [
15] showed that the microbiome at the time of embryo transfer could be successfully characterized without altering standard clinical practice. However, our vaginal samples were taken with a dry swab, and endometrial samples by Tao Brush, in order to discern the microbiome from both locations and avoid possible contamination. With reference to the taxonomic characterization in our study, similarly to Franasiak et al. [
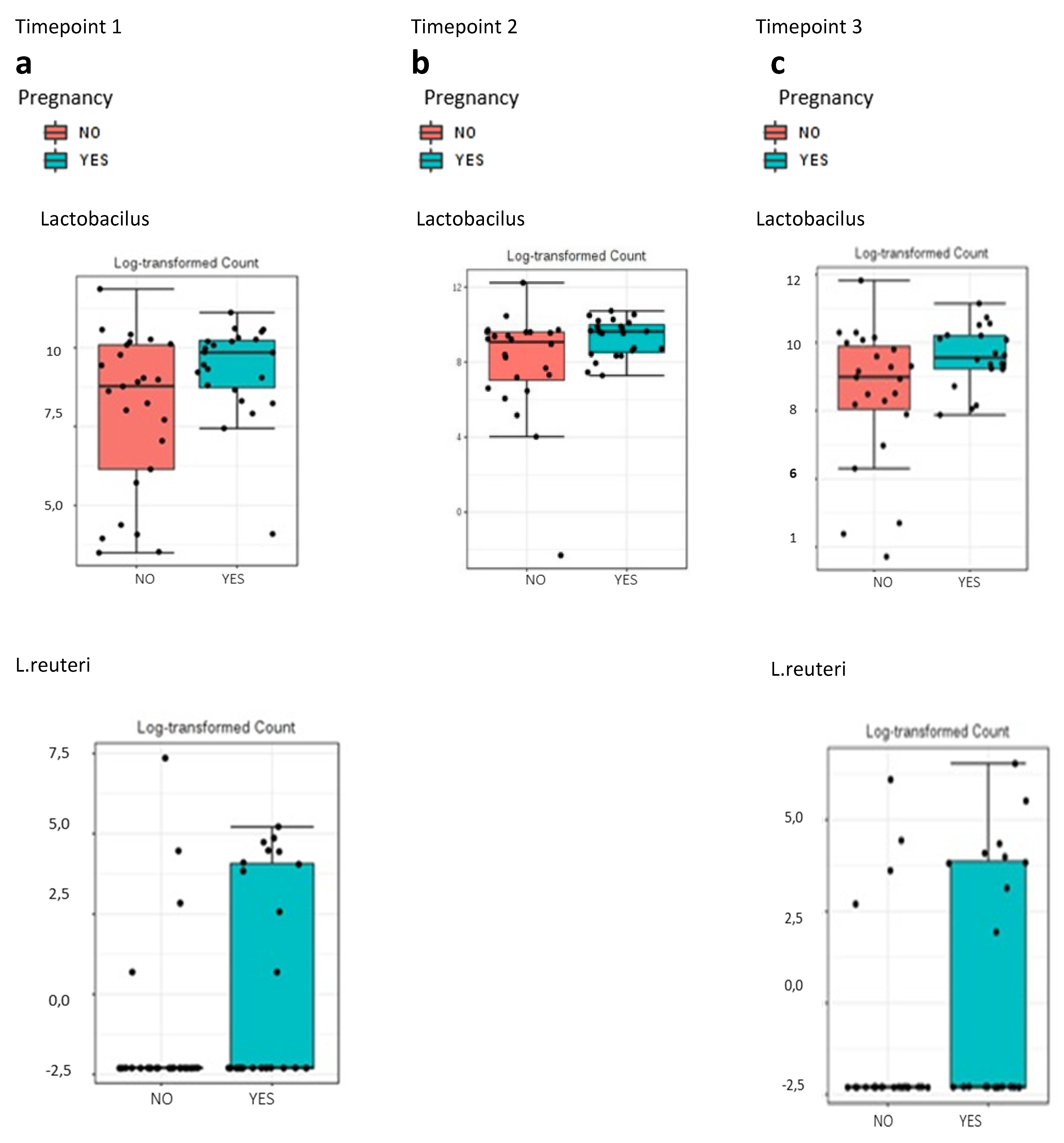
15], we also found that the relative abundance of bacteria of the genus
Lactobacillus was higher in vaginal samples of women who achieved clinical pregnancy after FET than of those who did not. However, Franasiak et al. [
15] did not obtain statistically significant values as we did. We also observed that
Streptococcus and
Prevotella may be associated with a poor prognosis with regard to gestation, while an abundance of
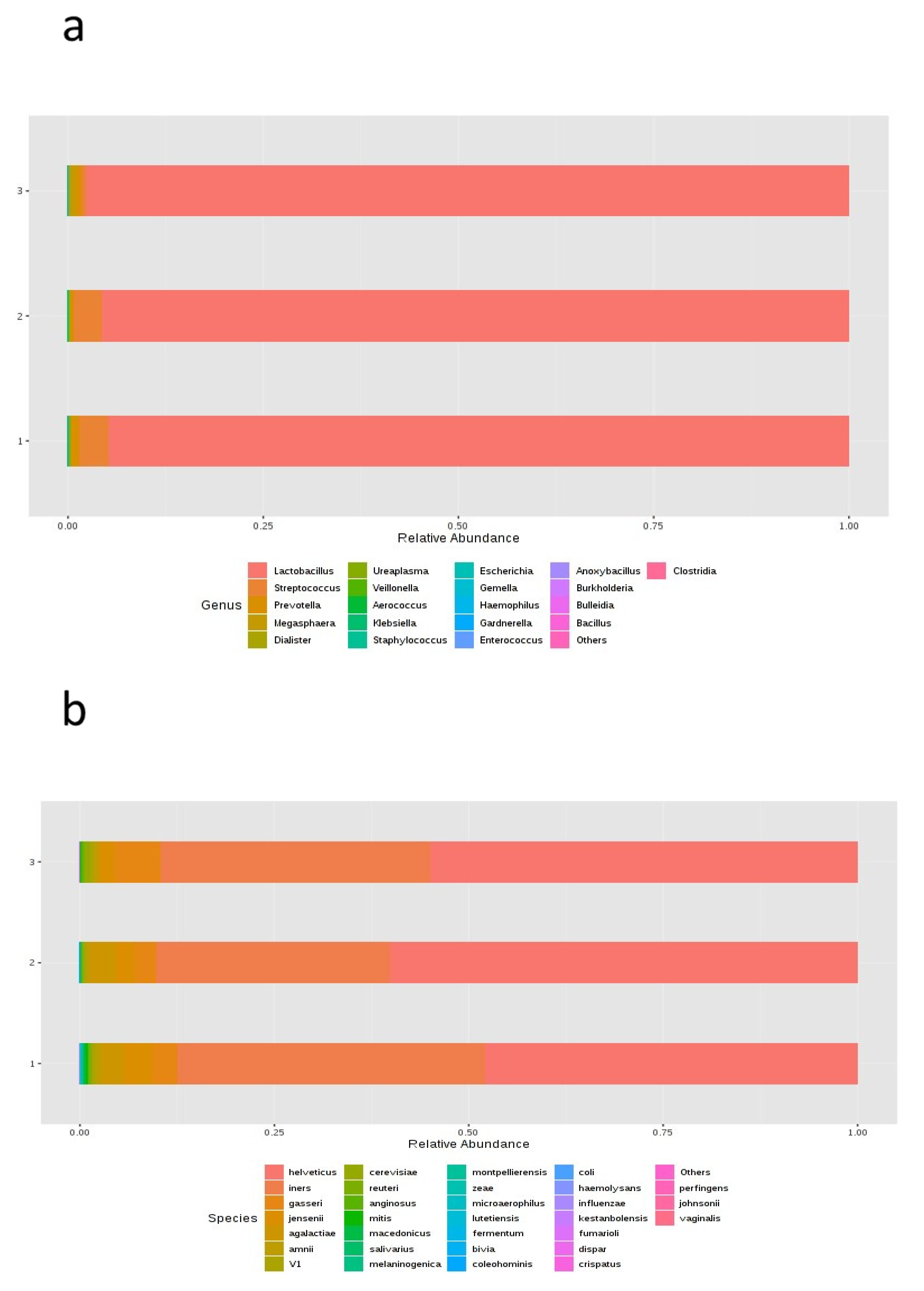
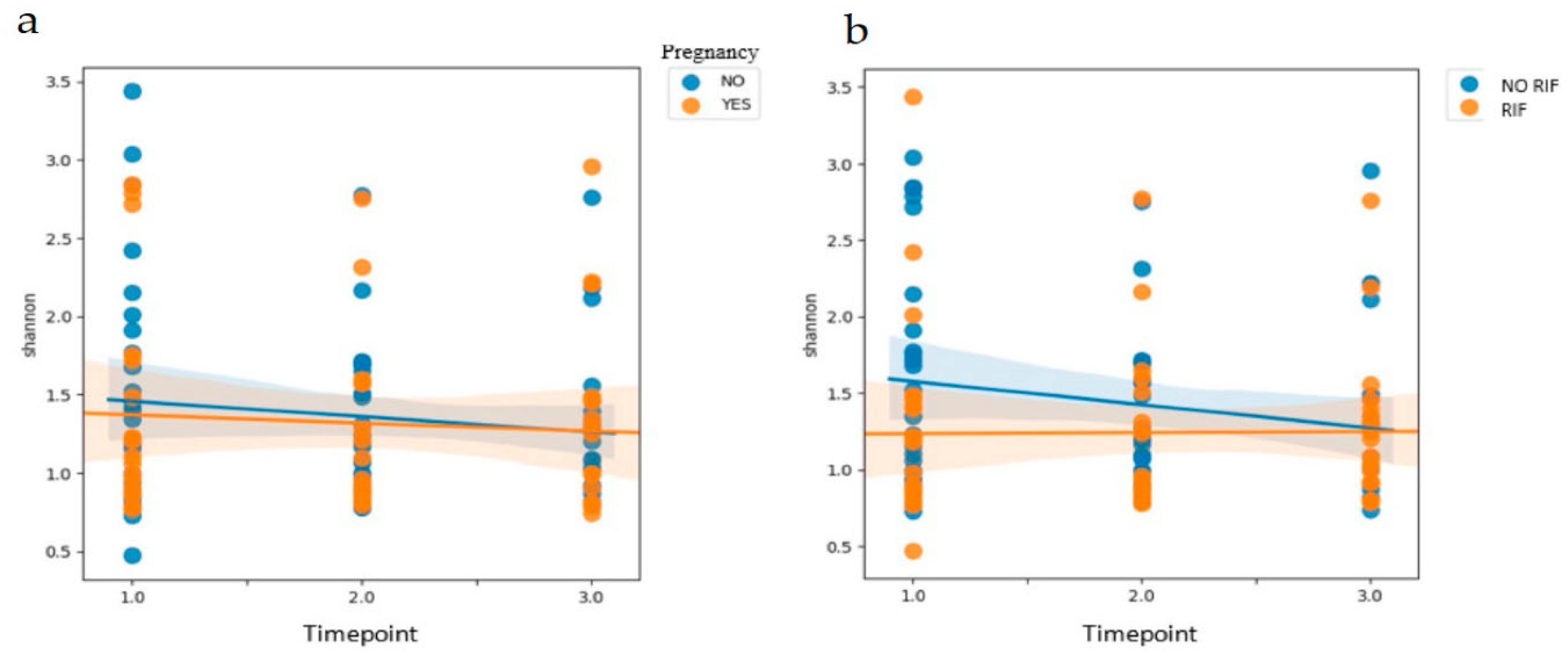
Lactobacillus spp. could be indicative of more favorable conditions. Since no statistically significant differences were obtained when analyzing the differences in relative abundance between samples taken at three different timepoints of the cycle, we concluded that the vaginal sample could be taken at any timepoint to obtain similar findings. The microbiome is usually affected by ovarian steroids, and the FET cycle may be related to higher local serum progesterone (P4) levels than in the previous cycle (timepoint 1). However, since all patients had one endometrial preparation per hormonal cycle substituted, we considered that this factor was homogeneous for all study participants.
Lactobacillus spp. are the most abundant bacteria in the vaginal samples. The ability of lactobacilli to inhibit infection without inducing inflammation can maximize fertility and favor pregnancy outcomes [
16]. In relation to the endometrium microbiome, we did not find differences between women who got pregnant and those who did not. The presence of a non-
Lactobacillus-dominated microbiome (NLDM)—i.e., < 90%
Lactobacillus spp. with > 10% of other bacteria—was associated with lower rates of implantation, pregnancy rate, pregnancy progression, and live birth [
17]. In this study, women classified as having an NLDM and showing a relative abundance of more than 80%
Lactobacillus spp. in the endometrium showed good pregnancy outcomes, suggesting that this threshold could be considered sufficient for embryonic implantation [
17]. In addition, even if classified as having an NLDM, the endometrium with a dominant quantity of
Bifidobacteria could also be an acceptable environment for implantation [
18]. Kyono et al. [
19] analyzed both the endometrial and vaginal microbiomes in the infertile Japanese population and assessed their impact on embryonic implantation. The bacterial status of the endometrium and vagina showed
Lactobacillus-dominated microbiota (> 90%
Lactobacillus spp.) in the endometria of 38% of IVF patients, and in the vaginas of 44.3%. The mean percentage of
Lactobacillus spp. in pregnant women was 96.45% (SD 33.61%). A considerably high proportion of NLDMs was found in the endometria of infertile Japanese women. A subsequent pilot study by Kyono et al. [
18] did show that the predominance of
Lactobacillus in the endometrium was favorable in terms of the pregnancy rate, but the results were not conclusive. A previous meta-analysis [
20] concluded that women with abnormal vaginal microbiota are approximately 1.4 times less likely to achieve first-trimester pregnancy after IVF treatment compared to women with normal microbiota; however, the methodologies of the included studies were heterogeneous.
In the present study, the taxonomic characterization showed differences in the microbial profiles between the vaginal and endometrial samples in terms of genera and species. The univariate analysis performed on the relative abundance of the different genera by sample type reached statistical significance for
Lactobacillus spp.,
Streptococcus spp.,
Ureaplasma spp.,
Delftia spp.,
Anaerobacillus spp.,
Ralstonia spp., and the species
L. helveticus. These results suggest that vaginal and endometrial samples are different in terms of both microbial diversity and the composition of the taxa. In a previous pilot study [
21], the dominant members of the microbial community were constant in the vagina and the cervix, and generally consistent in the endometrium; however, the relative proportions varied. In addition, infertile women tended to have
Ureaplasma spp. in the vagina and
Gardnerella spp. in the cervix more frequently than fertile women. In relation to the above-mentioned study, our analysis of alpha and beta indices for the microbial diversity of vaginal and endometrial samples revealed significant differences. Although the diversity of the vaginal microbiome could be expected a priori to be higher than that of the endometrial one due to factors such as more direct exposure to the outside of the body and direct contact with the seminal sample during sexual intercourse, the present study found a more diverse microbiome in the endometrium than in the vagina. This fact could be due to the non-dominance of a genus (more than 90% of relative abundance according to current scientific evidence), which can lead to colonization by other types of microorganisms and, thus, generate a more diverse environment, which might be related to infertility.
Another important aspect of the present study is the assessment of both diversity and taxonomic characterization according to participants’ history of RIF. Previously, the endometrial and vaginal microbiota were characterized by Kitaya et al. [
22] in infertile women with and without history of RIF. The microbiota detected in the endometrium showed significant variation in the composition of the bacterial community between the RIF and non-RIF groups, which was not observed in the vagina.
Burkholderia spp. were not detected in the microbiota of the endometrium in any sample women without RIF, but they were present in a quarter of the RIF patients. In our study, alpha diversity was higher at the endometrial level in women without RIF. This did not occur in vaginal samples, where no differences were observed. In addition, according to our findings and the available literature, a greater microbial diversity in the case of the vaginal and endometrial microbiota seems to be a scenario most unfavorable for the success of IVF treatments. The results of alpha diversity should be interpreted with caution, especially in patients with RIF. A detailed knowledge of the composition of this microbial pattern is necessary in order to know the % relative abundance of the genera, species, and subspecies present in the samples, and even to examine in depth the presence of pathogens that may have an effect on the vagina and/or endometrium. In the above-mentioned study, the authors found differences in the endometrial microbiota in both groups. When we analyzed the taxonomic characterization, we also observed clear differences in the relative abundances of the different genera and species in the endometrial microbiome between women with and without RIF. Unlike Kitaya et al. [
19], we found no differences with respect to the genus
Burkholderia. The generalized linear model showed that the vaginal microbiome pattern in women without RIF changed between different timepoints of the cycle, while in women with RIF it remained stable. This could be due to a possible adaptation of the microbiome pattern as a result of physiological changes occurring during the cycle. The possible causes and mechanisms involved in these modifications are still unknown and unstudied, but this could be a valuable line of future research.
In view of our study’s findings, before infertility treatment, both the vaginal and endometrial microbiomes can be examined as targets to improve pregnancy rates. A detailed knowledge of the composition of the microbiome pattern, together with the alpha and beta diversity analysis, might be helpful to guide the treatments.
Strengths and Limitations
Strengths of this study include the use of NGS to sequence the hypervariable regions of the 16S ribosome subunit. Another strength was the use of the Tao Brush to obtain the endometrial samples, as this device has a sheath that covers the specimen and closes before removal from the uterus and passage through the vagina; its design thus prevents any possible contamination by microorganisms from the vagina, and avoids the extraction of samples with patterns that are not directly associated with the endometrial microbiome. Moreover, all of the transferred embryos in the participants were previously analyzed via NGS to confirm chromosome normality and homogeneity, thus avoiding any bias due to embryological factors.
The main limitation was the sample size, which was small due to the pilot study design. Other limitations could be potential confounding stemming from recent sexual relations in some participants, which could have influenced the microbial compositions obtained. Finally, we only extracted endometrial samples at one timepoint in the cycle, in order to avoid compromising the treatment outcome and embryo implantation. This decision limited the amount of information we could obtain on the endometrial pattern, as no samples were taken for the other two timepoints. However, this limitation was partially overcome, as the vaginal samples were taken at three timepoints.
Further studies are needed in order to confirm our findings and to clarify the role of antibiotic and/or probiotic treatment in the normalization of the microbiome pattern and its consequences on clinical outcomes. Future research on the endometrial and vaginal microbiomes, along with their effects on and associations with reproductive health and infertility, should take a comprehensive approach, including an analysis of any interactions with the immune system and between different strains, metabolic and transcriptomic variables, and a study of biofilms—all of which are related to implantation and clinical pregnancy rates.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}