
The Time-Dependent Role of Bisphosphonates on Atherosclerotic Plaque Calcification

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vivo Study
2.2. Calcification Morphological Quantification
2.3. Serum Alkaline Phosphatase Activity and Total Cholesterol Assessment
2.4. X-ray Computed Tomography (X-ray CT)
2.5. Statistics
3. Results
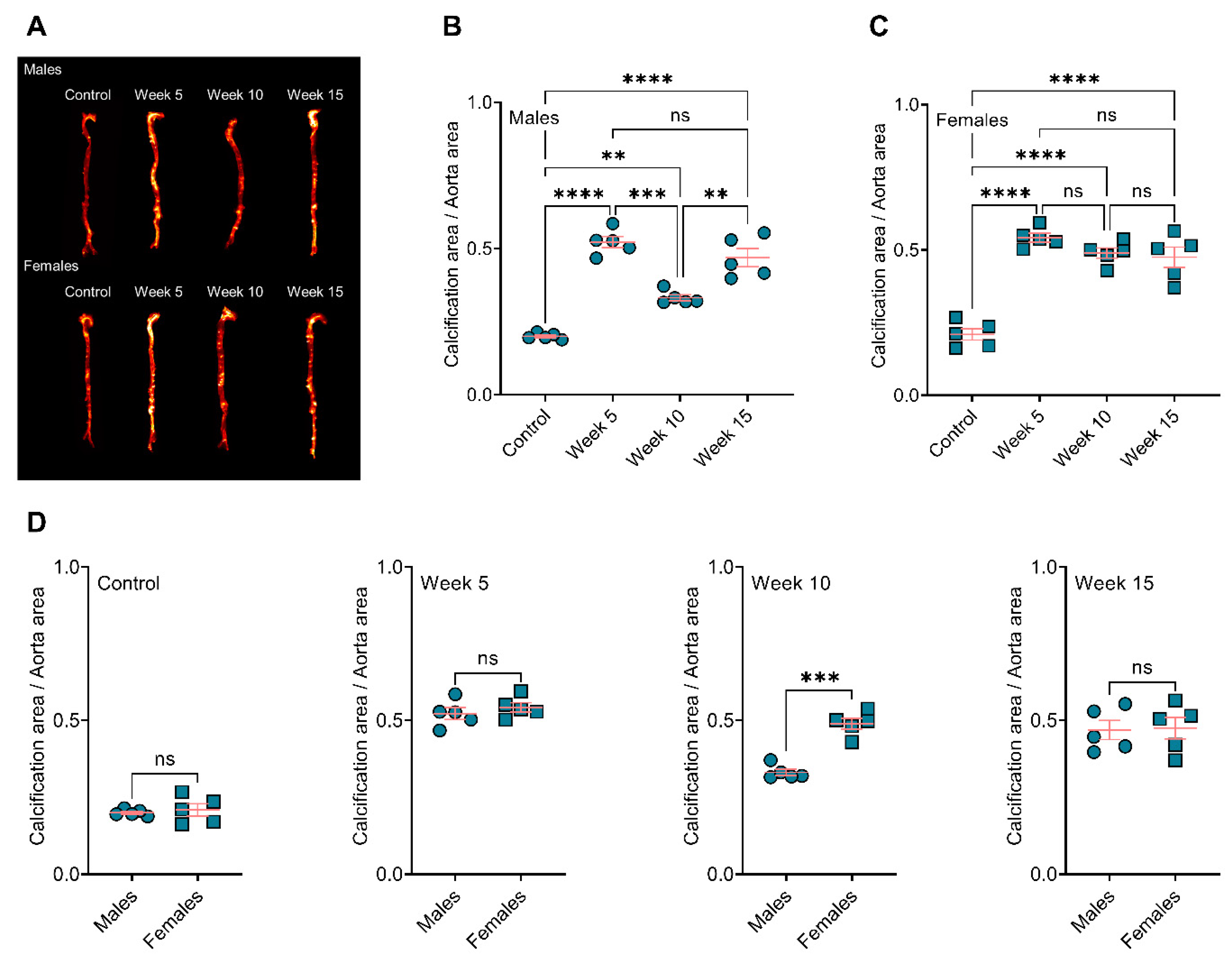
3.1. BiP Treatment Increases Atherosclerotic Plaque Calcification in the ApoE−/− Mice Fed an Atherogenic Diet
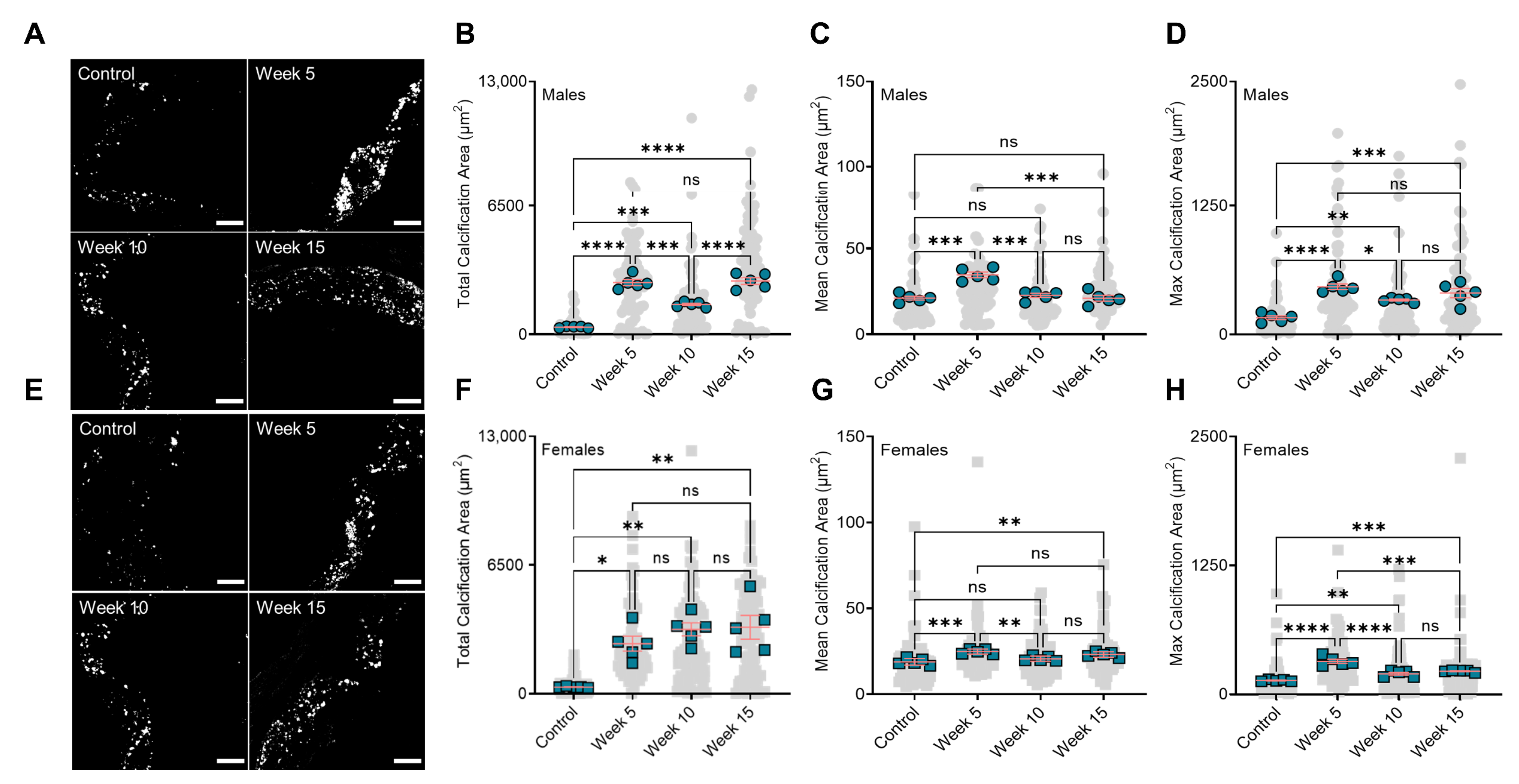
3.2. BiP Treatment Alters the Size of Minerals in the Atherosclerotic Plaque
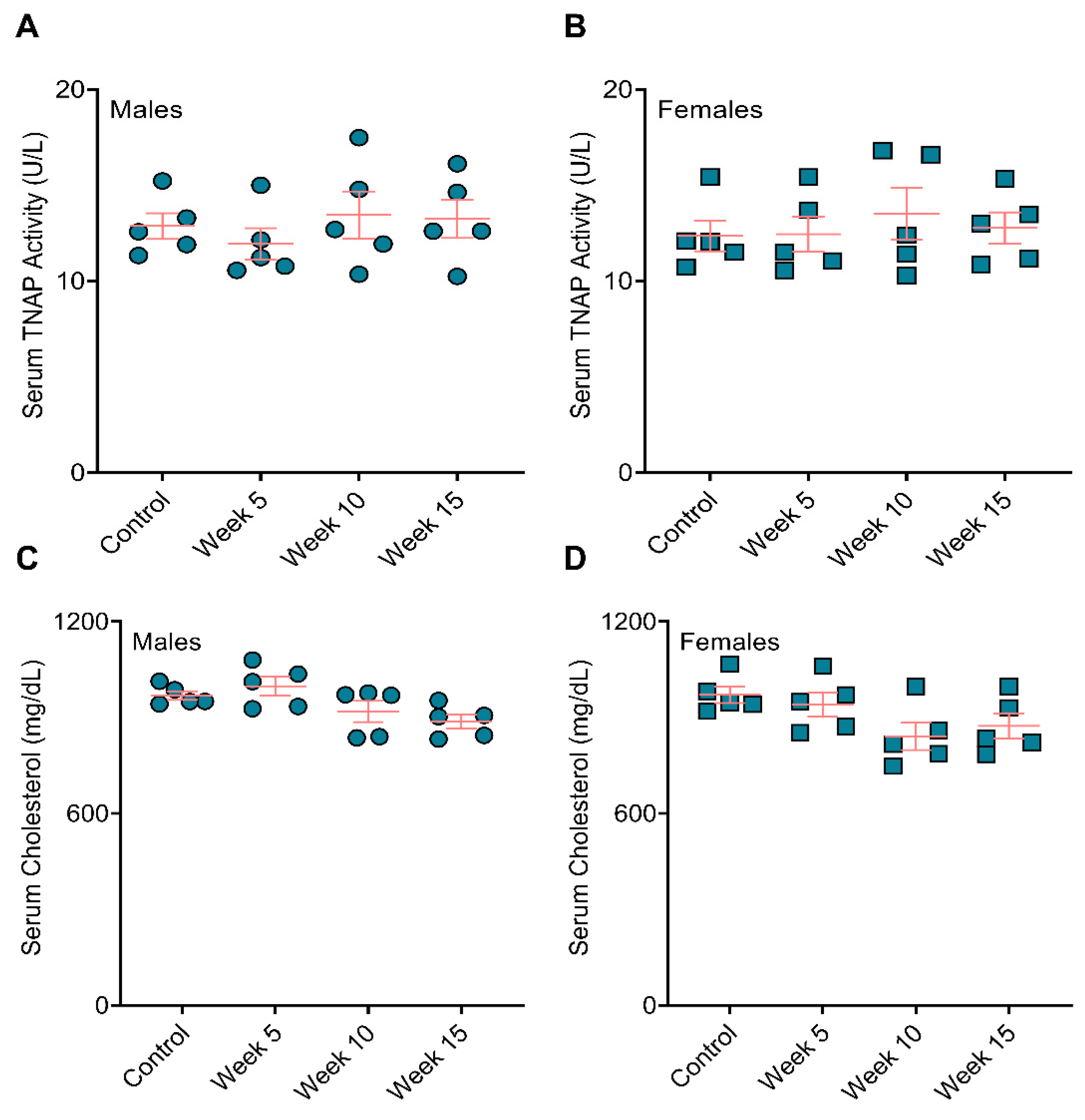
3.3. BiP Treatment Did Not Affect Serum Alkaline Phosphatase Activity and Total Cholesterol
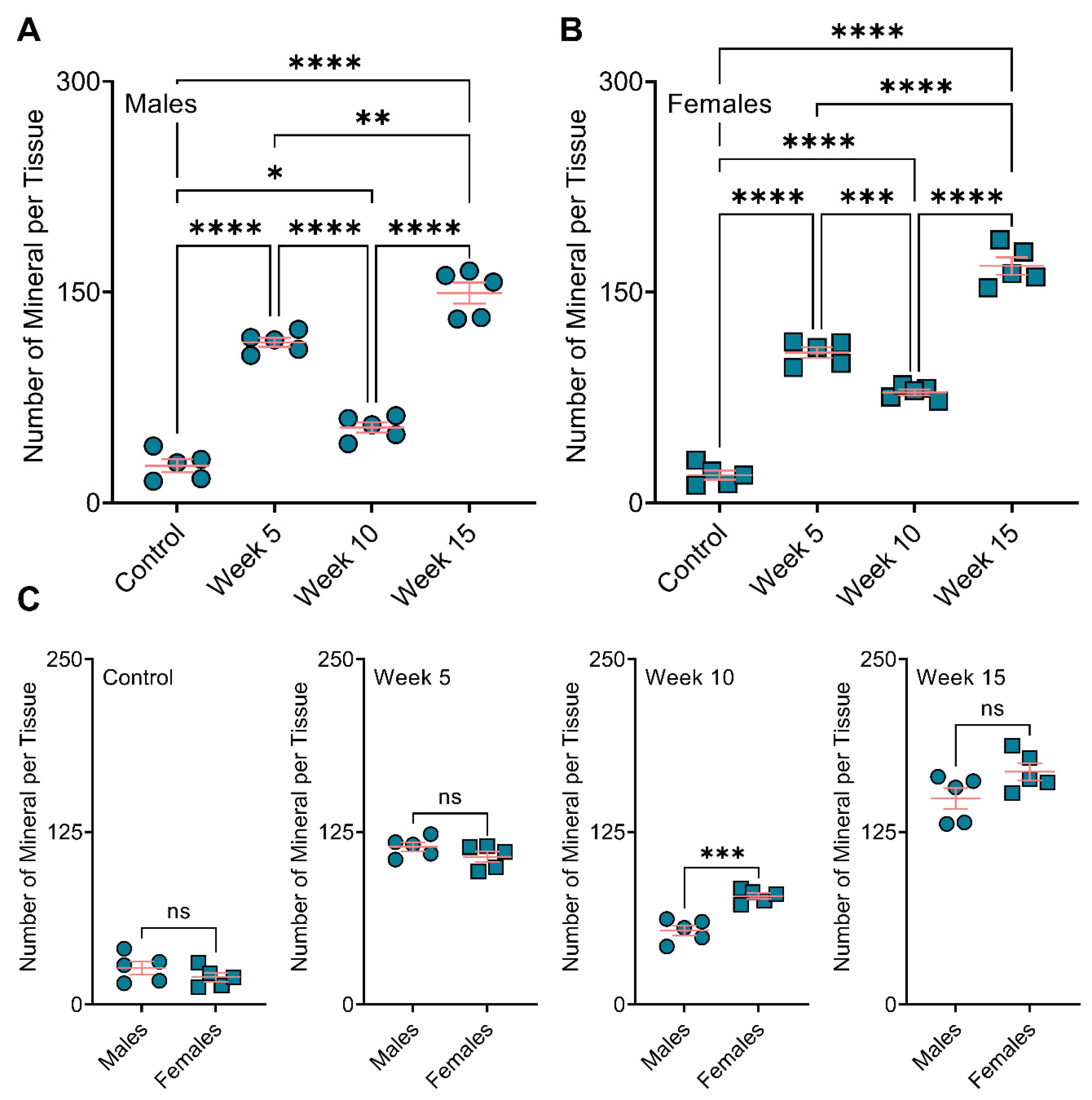
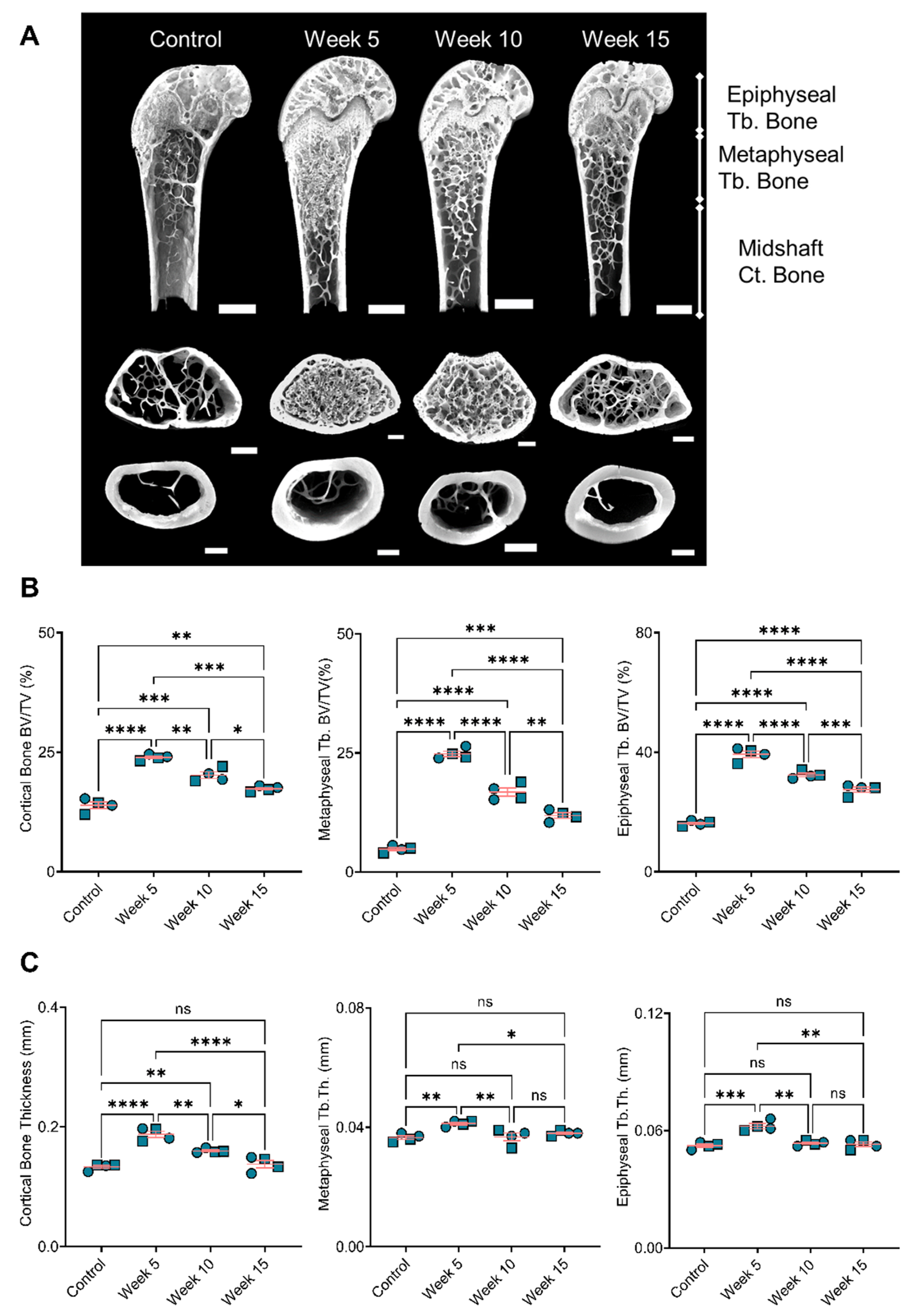
3.4. Bone Remodeling Positively Correlated with the BiP Treatment Duration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruiz, J.L.; Hutcheson, J.D.; Cardoso, L.; Bakhshian Nik, A.; Condado de Abreu, A.; Pham, T.; Buffolo, F.; Busatto, S.; Federici, S.; Ridolfi, A.; et al. Nanoanalytical analysis of bisphosphonate-driven alterations of microcalcifications using a 3D hydrogel system and in vivo mouse model. Proc. Natl. Acad. Sci. USA 2021, 118, e1811725118. [Google Scholar] [CrossRef]
- Hutcheson, J.D.; Goettsch, C.; Bertazzo, S.; Maldonado, N.; Ruiz, J.L.; Goh, W.; Yabusaki, K.; Faits, T.; Bouten, C.; Franck, G.; et al. Genesis and growth of extracellular-vesicle-derived microcalcification in atherosclerotic plaques. Nat. Mater. 2016, 15, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Bentzon, J.F.; Otsuka, F.; Virmani, R.; Falk, E. Mechanisms of Plaque Formation and Rupture. Circ. Res. 2014, 114, 1852–1866. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 Update. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef]
- Peng, A.W.; Mirbolouk, M.; Orimoloye, O.A.; Osei, A.D.; Dardari, Z.; Dzaye, O.; Budoff, M.J.; Shaw, L.; Miedema, M.D.; Rumberger, J.; et al. Long-Term All-Cause and Cause-Specific Mortality in Asymptomatic Patients with CAC ≥ 1000. JACC Cardiovasc. Imaging 2020, 13, 83–93. [Google Scholar] [CrossRef]
- Jin, H.Y.; Weir-McCall, J.R.; Leipsic, J.A.; Son, J.W.; Sellers, S.L.; Shao, M.; Blanke, P.; Ahmadi, A.; Hadamitzky, M.; Kim, Y.J.; et al. The Relationship Between Coronary Calcification and the Natural History of Coronary Artery Disease. JACC Cardiovasc. Imaging 2021, 14, 233–242. [Google Scholar] [CrossRef]
- Libby, P. Collagenases and cracks in the plaque. J. Clin. Investig. 2013, 123, 3201–3203. [Google Scholar] [CrossRef] [Green Version]
- Kelly-Arnold, A.; Maldonado, N.; Laudier, D.; Aikawa, E.; Cardoso, L.; Weinbaum, S. Revised microcalcification hypothesis for fibrous cap rupture in human coronary arteries. Proc. Natl. Acad. Sci. USA 2013, 110, 10741–10746. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, N.; Kelly-Arnold, A.; Cardoso, L.; Weinbaum, S. The explosive growth of small voids in vulnerable cap rupture; cavitation and interfacial debonding. J. Biomech. 2013, 46, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.L.; Hutcheson, J.D.; Aikawa, E. Cardiovascular calcification: Current controversies and novel concepts. Cardiovasc. Pathol. 2015, 24, 207–212. [Google Scholar] [CrossRef]
- Wang, Y.; Osborne, M.T.; Tung, B.; Li, M.; Li, Y. Imaging Cardiovascular Calcification. J. Am. Heart Assoc. 2018, 7, e008564. [Google Scholar] [CrossRef] [Green Version]
- Criqui, M.H.; Denenberg, J.O.; Ix, J.H.; McClelland, R.L.; Wassel, C.L.; Rifkin, D.E.; Carr, J.J.; Budoff, M.J.; Allison, M.A. Calcium Density of Coronary Artery Plaque and Risk of Incident Cardiovascular Events. JAMA 2014, 311, 271. [Google Scholar] [CrossRef]
- Recker, R.R.; Lewiecki, E.M.; Miller, P.D.; Reiffel, J. Safety of Bisphosphonates in the Treatment of Osteoporosis. Am. J. Med. 2009, 122, S22–S32. [Google Scholar] [CrossRef]
- Lampropoulos, C.E.; Papaioannou, I.; D’Cruz, D.P. Osteoporosis—A risk factor for cardiovascular disease? Nat. Rev. Rheumatol. 2012, 8, 587–598. [Google Scholar] [CrossRef]
- Neven, E.G.; De Broe, M.E.; D’Haese, P.C. Prevention of vascular calcification with bisphosphonates without affecting bone mineralization: A new challenge? Kidney Int. 2009, 75, 580–582. [Google Scholar] [CrossRef] [Green Version]
- Rodan, G.A.; Fleisch, H.A. Bisphosphonates: Mechanisms of action. J. Clin. Investig. 1996, 97, 2692–2696. [Google Scholar] [CrossRef]
- Bakhshian Nik, A.; Hutcheson, J.D.; Aikawa, E. Extracellular Vesicles as Mediators of Cardiovascular Calcification. Front. Cardiovasc. Med. 2017, 4, 78. [Google Scholar] [CrossRef] [Green Version]
- Kirchmayer, U.; Sorge, C.; Sultana, J.; Lapi, F.; Onder, G.; Agabiti, N.; Cascini, S.; Roberto, G.; Corrao, G.; Vitale, C.; et al. Bisphosphonates and cardiovascular risk in elderly patients with previous cardiovascular disease: A population-based nested case-control study in Italy. Ther. Adv. Drug Saf. 2019, 10, 2042098619838138. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-T.; Chen, J.-F.; Tsai, C.-J. The impact of bisphosphonates on mortality and cardiovascular risk among osteoporosis patients after cardiovascular disease. J. Formos. Med. Assoc. 2021, 120, 1957–1966. [Google Scholar] [CrossRef]
- Perkins, R.M.; Kirchner, H.L.; Matsushita, K.; Bucaloiu, I.D.; Norfolk, E.; Hartle, J.E. Bisphosphonates and Mortality in Women with CKD and the Presence or Absence of Cardiovascular Disease. Clin. J. Am. Soc. Nephrol. 2014, 9, 874–880. [Google Scholar] [CrossRef] [Green Version]
- Ylitalo, R.; Monkkonen, J.; Urtti, A.; Ylitalo, P. Accumulation of bisphosphonates in the aorta and some other tissues of healthy and atherosclerotic rabbits. J. Lab. Clin. Med. 1996, 127, 200–206. [Google Scholar] [CrossRef]
- Ylitalo, R.; Kalliovalkama, J.; Wu, X.; Kankaanranta, H.; Salenius, J.P.; Sisto, T.; Lahteenmaki, T.; Ylitalo, P.; Porsti, I. Accumulation of Bisphosphonates in Human Artery and their Effects on Human and Rat Arterial Functionin vitro. Pharmacol. Toxicol. 1998, 83, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Goettsch, C.; Iwata, H.; Aikawa, E. Parathyroid hormone: Critical bridge between bone metabolism and cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1333–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacey, A.; Qaradakhi, T.; Brennan-Speranza, T.; Hayes, A.; Zulli, A.; Levinger, I. Potential Role for Osteocalcin in the Development of Atherosclerosis and Blood Vessel Disease. Nutrients 2018, 10, 1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, Z.; Monier-Faugere, M.C.; Bauss, F.; Malluche, H.H. Short-term administration of the bisphosphonate ibandronate increases bone volume and prevents hyperparathyroid bone changes in mild experimental renal failure. Clin. Nephrol. 2000, 54, 45–53. [Google Scholar] [PubMed]
- Lomashvili, K.A.; Monier-Faugere, M.C.; Wang, X.; Malluche, H.H.; O’Neill, W.C. Effect of bisphosphonates on vascular calcification and bone metabolism in experimental renal failure. Kidney Int. 2009, 75, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Lo Sasso, G.; Schlage, W.K.; Boué, S.; Veljkovic, E.; Peitsch, M.C.; Hoeng, J. The Apoe−/− mouse model: A suitable model to study cardiovascular and respiratory diseases in the context of cigarette smoke exposure and harm reduction. J. Transl. Med. 2016, 14, 146. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Liu, C.C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Pirmohamed, M. Commentary—Bisphosphonates and calcium homoeostasis. Postgrad. Med. J. 2000, 76, 418–419. [Google Scholar] [CrossRef] [Green Version]
- Aikawa, E.; Nahrendorf, M.; Figueiredo, J.-L.; Swirski, F.K.; Shtatland, T.; Kohler, R.H.; Jaffer, F.A.; Aikawa, M.; Weissleder, R. Osteogenesis Associates With Inflammation in Early-Stage Atherosclerosis Evaluated by Molecular Imaging In Vivo. Circulation 2007, 116, 2841–2850. [Google Scholar] [CrossRef] [Green Version]
- MacAskill, M.G.; McDougald, W.; Alcaide-Corral, C.; Newby, D.E.; Tavares, A.A.; Hadoke, P.W.; Wu, J. Characterisation of an atherosclerotic micro-calcification model using ApoE−/− mice and PET/CT. Int. J. Cardiol. Heart Vasc. 2020, 31, 100672. [Google Scholar] [CrossRef] [PubMed]
- Domander, R.; Felder, A.A.; Doube, M. BoneJ2—Refactoring established research software. Wellcome Open Res. 2021, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Steiner, L.; Synek, A.; Pahr, D.H. Comparison of different microCT-based morphology assessment tools using human trabecular bone. Bone Rep. 2020, 12, 100261. [Google Scholar] [CrossRef] [PubMed]
- Rattazzi, M.; Bennett, B.J.; Bea, F.; Kirk, E.A.; Ricks, J.L.; Speer, M.; Schwartz, S.M.; Giachelli, C.M.; Rosenfeld, M.E. Calcification of Advanced Atherosclerotic Lesions in the Innominate Arteries of ApoE-Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1420–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emini Veseli, B.; Perrotta, P.; De Meyer, G.R.A.; Roth, L.; Van der Donckt, C.; Martinet, W.; De Meyer, G.R.Y. Animal models of atherosclerosis. Eur. J. Pharmacol. 2017, 816, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Whitman, S.C. A practical approach to using mice in atherosclerosis research. Clin. Biochem. Rev. 2004, 25, 81. [Google Scholar] [PubMed]
- Casula, M.; Olmastroni, E.; Galimberti, F.; Tragni, E.; Corrao, G.; Scotti, L.; Catapano, A.L. Association between the cumulative exposure to bisphosphonates and hospitalization for atherosclerotic cardiovascular events: A population-based study. Atherosclerosis 2020, 301, 1–7. [Google Scholar] [CrossRef]
- Maldonado, N.; Kelly-Arnold, A.; Vengrenyuk, Y.; Laudier, D.; Fallon, J.T.; Virmani, R.; Cardoso, L.; Weinbaum, S. A mechanistic analysis of the role of microcalcifications in atherosclerotic plaque stability: Potential implications for plaque rupture. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H619–H628. [Google Scholar] [CrossRef] [Green Version]
- Halling Linder, C.; Englund, U.H.; Narisawa, S.; Millan, J.L.; Magnusson, P. Isozyme profile and tissue-origin of alkaline phosphatases in mouse serum. Bone 2013, 53, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Mukaiyama, K.; Kamimura, M.; Uchiyama, S.; Ikegami, S.; Nakamura, Y.; Kato, H. Elevation of serum alkaline phosphatase (ALP) level in postmenopausal women is caused by high bone turnover. Aging Clin. Exp. Res. 2014, 27, 413–418. [Google Scholar] [CrossRef]
- Kovesdy, C.P.; Ureche, V.; Lu, J.L.; Kalantar-Zadeh, K. Outcome predictability of serum alkaline phosphatase in men with pre-dialysis CKD. Nephrol. Dial. Transplant. 2010, 25, 3003–3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploumis, A.; Donovan, J.M.; Olurinde, M.O.; Clark, D.M.; Wu, J.C.; Sohn, D.J.; O’Connor, K.C. Association between alendronate, serum alkaline phosphatase level, and heterotopic ossification in individuals with spinal cord injury. J. Spinal Cord Med. 2015, 38, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.Y.; Ben-Bassat, H.; Breuer, E.; Solomon, V.; Golomb, G. Structurally different bisphosphonates exert opposing effects on alkaline phosphatase and mineralization in marrow osteoprogenitors. J. Cell. Biochem. 1998, 68, 186–194. [Google Scholar] [CrossRef]
- Guney, E.; Kisakol, G.; Ozgen, A.G.; Yilmaz, C.; Kabalak, T. Effects of bisphosphonates on lipid metabolism. Neuroendocrinol. Lett. 2008, 29, 252–255. [Google Scholar]
- Adami, S.; Braga, V.; Guidi, G.; Gatti, D.; Gerardi, D.; Fracassi, E. Chronic Intravenous Aminobisphosphonate Therapy Increases High-Density Lipoprotein Cholesterol and Decreases Low-Density Lipoprotein Cholesterol. J. Bone Miner. Res. 2010, 15, 599–604. [Google Scholar] [CrossRef]
- Whyte, M.P.; Kempa, L.G.; McAlister, W.H.; Zhang, F.; Mumm, S.; Wenkert, D. Elevated serum lactate dehydrogenase isoenzymes and aspartate transaminase distinguish Albers-Schönberg disease (Chloride Channel 7 Deficiency Osteopetrosis) among the sclerosing bone disorders. J. Bone Miner. Res. 2010, 25, 2515–2526. [Google Scholar] [CrossRef]
- New, S.E.; Goettsch, C.; Aikawa, M.; Marchini, J.F.; Shibasaki, M.; Yabusaki, K.; Libby, P.; Shanahan, C.M.; Croce, K.; Aikawa, E. Macrophage-Derived Matrix Vesicles. Circ. Res. 2013, 113, 72–77. [Google Scholar] [CrossRef]
- Rogers, M.A.; Aikawa, M.; Aikawa, E. Macrophage heterogeneity complicates reversal of calcification in cardiovascular tissues. Circ. Res. 2017, 121, 5–7. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control n = 4 | Week 5 n = 4 | Week 10 n = 4 | Week 15 n = 4 |
|---|---|---|---|---|
| Distal femur | ||||
| Epiphyseal Trabecular Bone | ||||
| BV/TV (%) | 16.22 ± 0.37 | 39.3 ± 0.96 | 32.4 ± 0.5 | 27.48 ± 0.76 |
| Tb.N (mm−1) | 7.63 ± 1.11 | 12.86 ± 0.11 | 12.82 ± 0.32 | 12.05 ± 0.34 |
| Tb.Th (mm) | 0.052 ± 0.001 | 0.062 ± 0.001 | 0.053 ± 0.001 | 0.053 ± 0.001 |
| Tb.Sp (mm) | 0.59 ± 0.06 | 0.41 ± 0.02 | 0.42 ± 0.02 | 0.43 ± 0.02 |
| BS/BV (mm2/mm3) | 0.065 ± 0.002 | 0.059 ± 0.003 | 0.057 ± 0.003 | 0.062 ± 0.003 |
| EF | 0.017 ± 0.003 | 0.033 ± 0.005 | 0.034 ± 0.009 | 0.016 ± 0.003 |
| EFmax | 0.885 ± 0.004 | 0.902 ± 0.005 | 0.887 ± 0.004 | 0.88 ± 0.009 |
| EFmin | −0.82 ± 0.003 | −0.825 ± 0.008 | −0.812 ± 0.007 | −0.81 ± 0.009 |
| DA | 1.43 ± 0.05 | 1.39 ± 0.04 | 1.42 ± 0.03 | 1.4 ± 0.07 |
| Metaphyseal Trabecular Bone | ||||
| BV/TV (%) | 4.82 ± 0.29 | 24.8 ± 0.5 | 16.8 ± 0.75 | 11.86 ± 0.5 |
| Tb.N (mm−1) | 4.1 ± 0.3 | 13.1 ± 0.09 | 11.02 ± 0.07 | 7.62 ± 0.49 |
| Tb.Th (mm) | 0.036 ± 0.001 | 0.041 ± 0.001 | 0.037 ± 0.001 | 0.038 ± 0.001 |
| Tb.Sp (mm) | 0.56 ± 0.094 | 0.29 ± 0.04 | 0.27 ± 0.015 | 0.44 ± 0.022 |
| BS/BV (mm2/mm3) | 0.092 ± 0.003 | 0.085 ± 0.001 | 0.092 ± 0.003 | 0.092 ± 0.01 |
| EF | 0.12 ± 0.02 | 0.07 ± 0.005 | 0.086 ± 0.017 | 0.07 ± 0.03 |
| EFmax | 0.835 ± 0.019 | 0.87 ± 0.002 | 0.86 ± 0.012 | 0.855 ± 0.03 |
| EFmin | −0.67 ± 0.036 | −0.765 ± 0.013 | −0.71 ± 0.022 | −0.73 ± 0.05 |
| DA | 1.77 ± 0.14 | 1.6 ± 0.15 | 1.522 ± 0.09 | 1.78 ± 0.09 |
| Femoral midshaft | ||||
| Cortical Bone | ||||
| Mean Ct.Th (mm) | 0.133 ± 0.002 | 0.187 ± 0.004 | 0.159 ± 0.001 | 0.137 ± 0.005 |
| BV/TV (%) | 13.9 ± 0.48 | 23.9 ± 0.25 | 20.2 ± 0.6 | 17.3 ± 0.24 |
| Ps.Pm (mm) | 5.65 ± 0.11 | 5.37 ± 0.2 | 5.35 ± 0.22 | 5.33 ± 0.18 |
| Ec.Pm (mm) | 4.75 ± 0.11 | 4.2 ± 0.18 | 4.15 ± 0.2 | 4.17 ± 0.21 |
| J (mm4) | 0.49 ± 0.03 | 0.455 ± 0.04 | 0.417 ± 0.007 | 0.5 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakhshian Nik, A.; Ng, H.H.; Garcia Russo, M.; Iacoviello, F.; Shearing, P.R.; Bertazzo, S.; Hutcheson, J.D. The Time-Dependent Role of Bisphosphonates on Atherosclerotic Plaque Calcification. J. Cardiovasc. Dev. Dis. 2022, 9, 168. https://doi.org/10.3390/jcdd9060168
Bakhshian Nik A, Ng HH, Garcia Russo M, Iacoviello F, Shearing PR, Bertazzo S, Hutcheson JD. The Time-Dependent Role of Bisphosphonates on Atherosclerotic Plaque Calcification. Journal of Cardiovascular Development and Disease. 2022; 9(6):168. https://doi.org/10.3390/jcdd9060168
Chicago/Turabian StyleBakhshian Nik, Amirala, Hooi Hooi Ng, Manuel Garcia Russo, Francesco Iacoviello, Paul R. Shearing, Sergio Bertazzo, and Joshua D. Hutcheson. 2022. "The Time-Dependent Role of Bisphosphonates on Atherosclerotic Plaque Calcification" Journal of Cardiovascular Development and Disease 9, no. 6: 168. https://doi.org/10.3390/jcdd9060168