
Developmental Shifts in the Microbiome of a Cosmopolitan Pest: Unraveling the Role of Wolbachia and Dominant Bacteria
by

Xiangyu Zhu
1,2,3, 
Jinyang Li
1,2,3,
Ao He
1,2,3,
Geoff M. Gurr
1,2,4, 
Minsheng You
1,2,3 and
Shijun You
1,2,3,* 1
State Key Laboratory for Ecological Pest Control of Fujian and Taiwan Crops, Institute of Applied Ecology, Fujian Agriculture and Forestry University, Fuzhou 350002, China
2
Joint International Research Laboratory of Ecological Pest Control, Ministry of Education, Fuzhou 350002, China
3
Ministerial and Provincial Joint Innovation Centre for Safety Production of Cross-Strait Crops, Fujian Agriculture and Forestry University, Fuzhou 350002, China
4
Gulbali Institute, Charles Sturt University, Orange, NSW 2800, Australia
*
Author to whom correspondence should be addressed.
Insects 2024, 15(2), 132; https://doi.org/10.3390/insects15020132
Submission received: 14 January 2024
/
Revised: 7 February 2024
/
Accepted: 8 February 2024
/
Published: 16 February 2024
(This article belongs to the Section Insect Behavior and Pathology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
The diamondback moth (Plutella xylostella) is a major pest of cruciferous vegetables worldwide. The dominant Wolbachia strain plutWB1 is associated with substantial sex ratio distortion of P. xylostella. In this study, high-throughput 16S rDNA amplicon sequencing was performed to investigate the bacterial community diversity and dynamics across different developmental life stages and Wolbachia infection states in P. xylostella. Our results will provide a framework for studies of Wolbachia and the associated microbiome in P. xylostella interactions.
Abstract
Wolbachia bacteria (phylum Proteobacteria) are ubiquitous intracellular parasites of diverse invertebrates. In insects, coevolution has forged mutualistic associations with Wolbachia species, influencing reproduction, immunity, development, pathogen resistance, and overall fitness. However, the impact of Wolbachia on other microbial associates within the insect microbiome, which are crucial for host fitness, remains less explored. The diamondback moth (Plutella xylostella), a major pest of cruciferous vegetables worldwide, harbors the dominant Wolbachia strain plutWB1, known to distort its sex ratio. This study investigated the bacterial community diversity and dynamics across different developmental life stages and Wolbachia infection states in P. xylostella using high-throughput 16S rDNA amplicon sequencing. Proteobacteria and Firmicutes dominated the P. xylostella microbiome regardless of life stage or Wolbachia infection. However, the relative abundance of dominant genera, including an unclassified genus of Enterobacteriaceae, Wolbachia, Carnobacterium, and Delftia tsuruhatensis, displayed significant stage-specific variations. While significant differences in bacterial diversity and composition were observed across life stages, Wolbachia infection had no substantial impact on overall diversity. Nonetheless, relative abundances of specific genera differed between infection states. Notably, Wolbachia exhibited a stable, high relative abundance across all stages and negatively correlated with an unclassified genus of Enterobacteriaceae, Delftia tsuruhatensis, and Carnobacterium. Our findings provide a foundational understanding of the complex interplay between the host, Wolbachia, and the associated microbiome in P. xylostella, paving the way for a deeper understanding of their complex interactions and potential implications for pest control strategies.
1. Introduction
Insects, the most species-rich and abundant animal class, have captivated researchers for centuries due to their ecological and economic importance [1]. Recently, the complex relationship between insects and their resident microbial communities, the microbiome, has become a hot topic [2,3]. This microbiome plays a crucial role in insect diversity and successful adaptation [4]. A wealth of data reveals the ubiquity and remarkable diversity of microbial communities within insects [5,6,7]. These communities are essential for various biological processes, including growth and development [8], reproduction [9,10], immunity [11,12,13], and metabolism [14,15,16]. Insects rely heavily on their microbiome for survival and normal functions. Depending on the degree of dependence, bacteria can be classified as obligate or facultative [2,17]. Numerous examples highlight the importance of specific microorganisms. For instance, the obligate symbiont Buchnera aphidicola in aphids fulfills part of their nutritional needs, impacting their survival and reproduction [18]. Similarly, Candidatus Ishikawaella capsulatus is crucial for the survival of the shield bug Megacopta cribraria [19]. Vertically transmitted facultative symbionts like Wolbachia and Spiroplasma also have significant effects on reproduction and other aspects in many insect hosts [9,20].
While the structure and diversity of insect microbial communities are influenced by various factors, the underlying mechanisms responsible for shaping microbiomes remain largely unclear. Host life stage (i.e., egg, larva, pupa, and adult) plays a key role, though insects can maintain beneficial microbes across different life stages [21,22,23]. Diet temperature, humidity, elevation, and precipitation can have profound impacts [5,24]. Additionally, microbial communities are shaped by interactions among themselves. For example, the Cry1Ac protoxin from Bacillus thuringiensis (Bt) dynamically altered the P. xylostella microbiome, significantly reducing its diversity [25].
Wolbachia, a genus of facultative endosymbiotic bacteria (phylum Proteobacteria), manipulates host reproduction through diverse strategies like cytoplasmic incompatibility (CI), male-killing, feminization, and parthenogenesis [26]. Initially discovered in mosquitoes, Wolbachia is now known from a vast array of insects [27,28,29]. Beyond manipulating its own propagation, Wolbachia interacts with other microbes, influencing insect hosts on multiple levels. For example, in Drosophila melanogaster and D. simulans, Wolbachia infection mitigates RNA virus-induced death [30,31,32]. Conversely, other microbes can negatively affect Wolbachia, as seen in D. melanogaster where Spiroplasma NSRO reduces the density of Wolbachia wMel [33]. Similarly, the native microbiome of Anopheles mosquitoes hinders Wolbachia’s vertical transmission [34]. These examples highlight the complex interplay between Wolbachia, other microbes, and the host, suggesting that studying Wolbachia phenotypes should also consider its impact on the host’s microbial community.
The diamondback moth, Plutella xylostella L. (Lepidoptera: Plutellidae), is a cosmopolitan agricultural pest of cruciferous vegetables [35]. Its life cycle includes egg, larva (four instars), pupa, and adult stages. Recent research on P. xylostella microbes has focused primarily on the gut community [25,36,37]. Wolbachia infection exists in its wild populations, with the dominant strain plutWB1 potentially causing sex ratio distortion [38,39]. In this study, we employ 16S rRNA gene amplicon Illumina sequencing to explore the microbial communities of Wolbachia-infected (WI) and uninfected (WU) P. xylostella. WI P. xylostella, initially collected in Nepal with 100% plutWB1 infection, serves as the infected population. The WU population, derived from WI individuals treated with antibiotics to eliminate Wolbachia, provides a control with the same genetic background. Notably, after 10 generations, the bacterial community of WU P. xylostella converges with another lab-reared population (different genetic background) lacking Wolbachia infection [40]. To minimize antibiotic effects on P. xylostella itself, we maintained the WI and WU populations for 50 generations after antibiotic treatment before sequencing. Our study represents the first exploration of P. xylostella’s microbial communities across development and Wolbachia infection states. It lays the foundation for further investigation into the intricate interactions between Wolbachia and other bacteria within this important pest.
2. Materials and Methods
2.1. Sample Collection
The WI and WU P. xylostella were used in this study as previously described [40]. Briefly, the WI P. xylostella was initially collected in Nepal; because of the 100% plutWB1 infection rate in WI P. xylostella, we first used rifampicin treatment (1 mg/mL) to obtain the WU P. xylostella. The WI and WU populations were raised in our laboratory for over 50 generations under conditions of 75% ± 5% relative humidity (RH), 25 ± 1 °C temperature, and a photoperiod of 14 to 10 h (light to dark). A total of 10 different developmental stages were established for sample collection. In addition to the conventional six developmental stages (egg, 1–4 instar larvae, pupae), we also subdivided adult P. xylostella males and females into before and after mating. Each group was set with five biological replicates, and a total of 100 P. xylostella samples were collected from 10 different developmental stages of two populations. Samples were collected using a 1.5 mL sterile centrifuge tube, and each sample was placed in a separate centrifuge tube. Eggs were 1-day old, and 50–70 eggs were pooled in each centrifuge tube (considered as an egg stage sample) to obtain sufficient DNA for DNA extraction. The P. xylostella males and females (before and after mating) were all collected at 1 day old. Approximately 20 first instar larvae were placed in each 1st instar larvae sample; approximately 15 second instar larvae were placed in each 2nd instar larvae sample; approximately 10 third instar larvae were placed in each 3rd instar larvae sample; approximately 8 corresponding instars were placed in each of the 4th instar larvae, pupae, and adult samples. Each sample was surface-disinfected with 75% alcohol immediately after sampling. After surface disinfection, all samples were quick-frozen using liquid nitrogen and stored at −80 °C until DNA extraction.
2.2. DNA Extraction and 16S rRNA Gene Amplification
To avoid contamination, the entire sample preparation and DNA extraction process was performed on an ultra-clean workbench. After each tube of sample was frozen in liquid nitrogen, 95% sterilized zirconia beads were placed separately and crushed using a tissue crusher (QIAGEN, Germantown, MD, USA). Subsequent DNA extraction was performed using the DNeasy® PowerSoil® Pro Kit (QIAGEN, Germantown, MD, USA) and according to the instructions provided. DNA purity and concentration were detected using the NanoDrop2000, and DNA integrity was checked using 1% agarose gel electrophoresis. The bacterial universal primers (8F (AGAGTTTGATCCTGGCTCAG) and 1492R (GGTTACCTTGTTACGACTT)) were used to verify the quality of the extracted DNA [41]. The PCR amplification program was set up as follows: initial denaturation at 94 °C for 4 min; then 30 cycles at 94 °C for 30 s, 65 °C for 40 s, and 72 °C for 90 s; and finally, 72 °C extension for 10 min. The PCR products were detected by 1% agarose gel electrophoresis with a single bright band of about 1500 bp.
PCR amplification was performed using primers 338F (ACTCCTACGGGGAGGCAGCAG)—806R (GGACTACHVGGGGTWTCTAAT) specific for the V3-V4 region of the 16S rRNA [42]. The PCR products were detected by 2% agarose gel electrophoresis. The PCR products of all 100 P. xylostella samples were purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) according to the provided instructions and quantified using Quantus™ Fluorometer (Promega, Madison, WI, USA). The library was constructed using the NEXTFLEX Rapid DNA-Seq Kit and sequenced using Illumina’s Miseq PE300 platform (Shanghai Meiji Biomedical Technology Co., Ltd., Shanghai, China).
2.3. Sequencing Data Analysis
Raw sequences were quality-controlled using fastp (v 0.20.0) [43] and spliced in FLASH (v1.2.11) [44]. Bases with quality values below 20 in the tails of the reads were filtered. Then, pairs of reads were merged into a single sequence based on the overlap relationship between the reads, with a minimum overlap length of 10 bp. Based on 97% similarity, OTU clustering was performed using UPARSE software (v 7.1) [45]. Based on the Silva 16S rRNA gene database (v138) [46], representative sequences of OTUs with 97% similarity level were taxonomically annotated using the RDP classifier Bayesian algorithm with the confidence threshold set at 70% [47], and the community composition of each sample was counted at different species classification levels.
2.4. Diversity Analysis
Alpha diversity indices were calculated using mothur software (v1.30) [48] and tested for between-group differences using SPSS v24.0 software (Chicago, IL, USA). Different alpha diversity indices can be used to characterize changes in bacterial species richness (number of bacterial species) and evenness (relative bacterial abundance) among different populations and developmental stages. We calculated a total of four alpha diversity indices: Chao1 and ACE indices reflecting species richness, and Shannon and Simpson indices reflecting bacterial evenness. Larger Chao1, ACE, and Shannon indices indicate higher community diversity; larger Simpson indices indicate lower community diversity. The alpha diversity indices of different developmental stages of WI and WU P. xylostella populations were compared using one-way ANOVA, and the alpha diversity indices of certain developmental stages of WI and WU P. xylostella populations that were more different were compared using independent t-tests.
Beta diversity analysis was performed using non-metric multidimensional scaling (NMDS) based on the Bray-Curtis distance algorithm to test the similarity of microbial community structure among different subgroups of samples, and analysis of similarities (ANOSIM) for statistical difference analysis [49]. We compared the beta diversity of the different subgroups by grouping them according to developmental stages, Wolbachia infection status, sex, and mating status, respectively. The closer the two sample points are, the more similar the species composition of the two samples. Horizontal and vertical coordinates represent relative distances and have no practical significance. The Stress value is used to test the NMDS analysis result. The two-dimensional dot plot of NMDS is generally considered reliable when stress values are less than 0.2.
2.5. Correlation Analysis and Putative Functional Profiling
Based on the sequencing data and the analysis mentioned above, the dominant bacterial genera in each developmental life stage were identified. Spearman correlation coefficients (ρ) between Wolbachia and other dominant genera in the P. xylostella samples at different developmental stages were calculated using the cor function in R package stats v4.2.0 and visualized using heatmaps in R package corrplot v0.92. To further investigate the functional differences in the bacterial community that may result from Wolbachia infection and different developmental stages, and to further explore the relationship between differences in bacterial communities and functional differences, the functional gene composition of the bacterial communities was predicted based on different groupings of bacterial 16s rRNA gene OTUs using PICRUSt2 software (v 2.2.0) [50].
3. Results
3.1. An Overview of P. xylostella Bacterial Microbiota
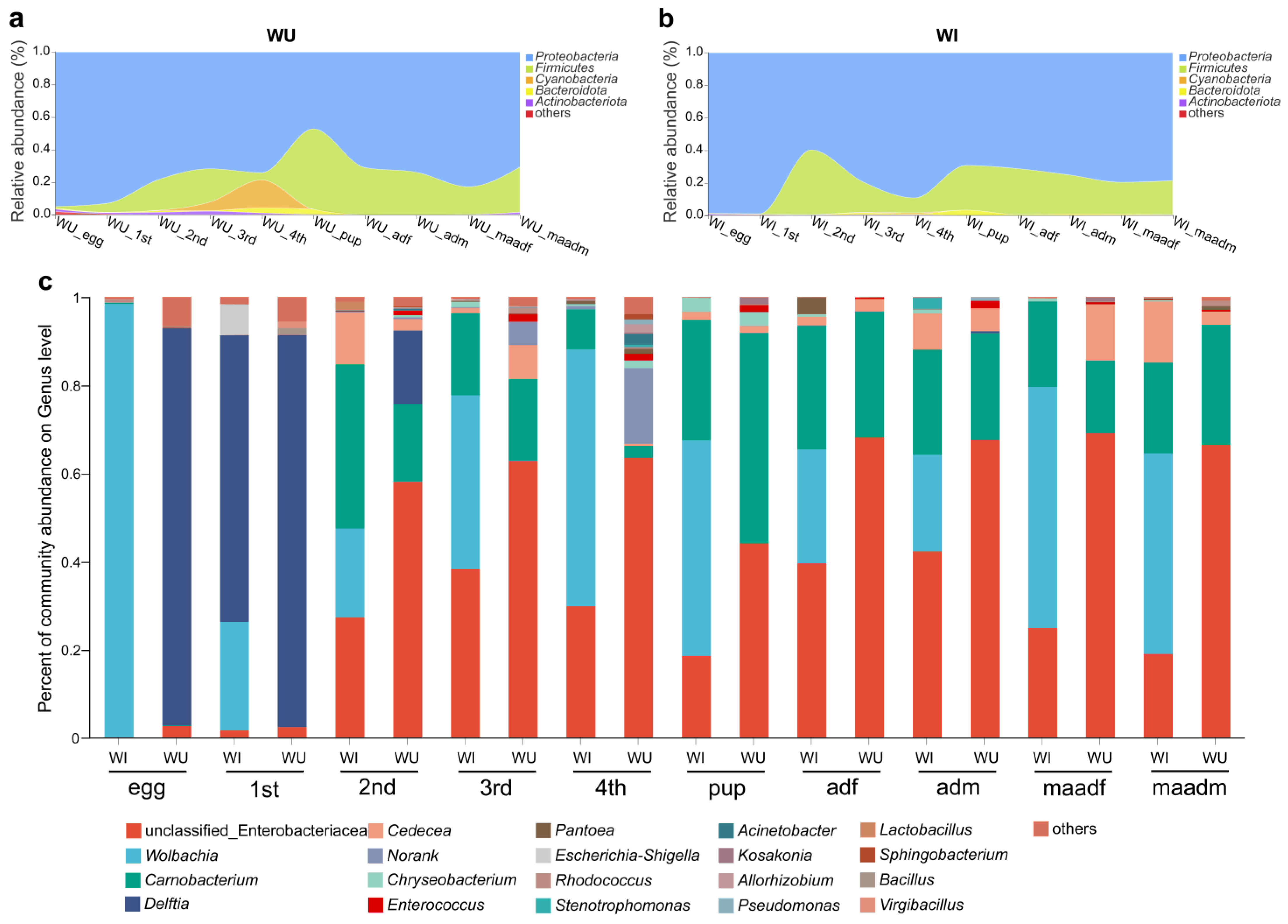
From WI and WU populations of different developmental stages (eggs, larvae (1–4 instar), pupae, males (unmated and mated) and females (unmated and mated)), we collected a total of 100 P. xylostella samples. A total of 3,728,553 optimized sequences were obtained from the 100 P. xylostella samples (after double-ended sequence quality control splicing), with an average sequence length of 422 bp. The dilution curves of all samples flattened with increasing sequencing depth, indicating that the sequencing efficiently covered the actual bacterial diversity of each sample. The coverage of each sample was greater than 99% (Tables S1 and S2). Based on 97% similarity, a total of 2009 OTUs were identified belonging to 41 phyla. Overall, regardless of the developmental stages and Wolbachia infection states, the bacterial microbiota of P. xylostella were dominated by the phyla Proteobacteria and Firmicutes (Figure 1a,b). Within the Proteobacteria, the microbiota was dominated by the following genera: an unclassified genus of Enterobacteriaceae, Wolbachia, Delftia, and Cedecea. Within the Firmicutes, the microbiota was dominated by the genus Carnobacterium (Figure 1c).
3.2. Microbiome Composition Shifts during Developmental Stages
By analyzing and comparing the composition of microbial communities in different developmental stages of WI and WU P. xylostella, we found that the 16S rRNA amplicon profiles at both the phylum and genus levels changed dramatically with developmental stage (egg, larvae, pupae, and adults).
In WU P. xylostella, the two major phyla Proteobacteria (ranging from 47.2% to 95%) and Firmicutes (ranging from 0.9% to 49.4%) represented the majority of taxa at all developmental stages (ranging from 78.6% to 99.9%) (Figure 1a). The relative abundance of the phylum Cyanobacteria varied significantly, with relative abundance below 1% in all developmental stages except third and fourth instar larvae, with relative abundance of about 5.3% in third instar larvae and as high as about 17.2% in fourth instar larvae. At the genus level, the top three genera include an unclassified genus of Enterobacteriaceae, Carnobacterium, and Delftia (Figure 1c). In most developmental stages, an unclassified genus of Enterobacteriaceae was the most abundant genus (second instar larvae 58%, third instar larvae 62.7%, fourth instar larvae 63.4%, unmated female adults 68.1%, unmated male adults 67.5%, mated female adults 69%, and mated male adults 66.4%). In the egg and first instar larval stage, Delftia was the most abundant genus (eggs 90.1%, first instar larvae 89%). In the pupal stage, Carnobacterium was the most abundant genus (47.8%).
In WI P. xylostella, the two major phyla Proteobacteria (ranging from 59.7% to 99.9%) and Firmicutes (ranging from 0.3% to 39.7%) represented the majority of taxa at all developmental stages (ranging from 96.7% to 99.4%) (Figure 1b). At the genus level, Wolbachia was the most abundant in most developmental stages (eggs 98.4%, third instar larvae 39.5%, fourth instar larvae 58.3%, pupae 48.9%, mated female adults 54.7%, mated male adults 45.5%) (Figure 1c). In the first instar larval stage, Delftia was the most abundant genus (65%). In the second instar larval stage, Carnobacterium was the most abundant genus (37.3%). In the unmated adults (male and female), an unclassified genus of Enterobacteriaceae was the most abundant genus (unmated female adults 39.6%, unmated male adults 42.4%).
3.3. Possible Effects of Wolbachia Infection on the Microbiota of P. xylostella
Next, we compared the dominant phyla and genera of WI and WU P. xylostella in each developmental stage. In WI P. xylostella, Wolbachia was one of the most abundant genera, especially in eggs; the composition of microbial communities in different developmental stages of WI and WU P. xylostella differed significantly based on Wolbachia infection status. Aside from Wolbachia, dominant phyla and genera were identical for WI and WU P. xylostella strains. The relative abundance of dominant phyla (Proteobacteria and Firmicutes) and genera (an unclassified genus of Enterobacteriaceae, Carnobacterium, and Delftia) were different between WI and WU P. xylostella at the same developmental stage. The bacterial microbiota composition of WI and WU P. xylostella differed significantly at the phylum level in the fourth instar larval stage. The dominant phyla in the WU fourth instar larval stage were Proteobacteria (74.1%) and Cyanobacteria (17.2%), while the dominant phyla in the WU fourth instar larval stage were Proteobacteria (89.2%) and Firmicutes (9.1%). Due to the presence of Wolbachia in WI P. xylostella, differences in bacterial microbiota composition between WI and WU P. xylostella were more pronounced at the genus level. The developmental stage with the greatest difference in genus level is the egg stage, the relative abundance of Wolbachia was absolutely dominant in the WI egg stage (98.4%), while the dominant genus in the WU egg stage was Delftia tsuruhatensis (90.1%).
3.4. Sex and Mating Status Effects
In WU P. xylostella adults, the relative abundance of the two major genera an unclassified genus of Enterobacteriaceae (ranging from 66.4% to 69%) and Carnobacterium (ranging from 16.5% to 28.5%) were similar. In WI P. xylostella adults, the three major genera an unclassified genus of Enterobacteriaceae, Wolbachia, and Carnobacterium represented the majority of taxa. The relative abundance of the genus Carnobacterium was at relatively similar levels in different sex and mating situations of WI adults (ranging from 19.4% to 28.1%). The genera an unclassified genus of Enterobacteriaceae and Wolbachia had little difference in relative abundance between females and males, but had a significant difference before and after mating (an unclassified genus of Enterobacteriaceae (unmated females = 39.6%, unmated males = 42.2%, mated females = 24.9%, mated males = 18.9%); Wolbachia (unmated females = 25.8%, unmated males = 21.9%, mated females = 54.7%, mated males = 45.5%)).
3.5. Alpha and Beta Diversity
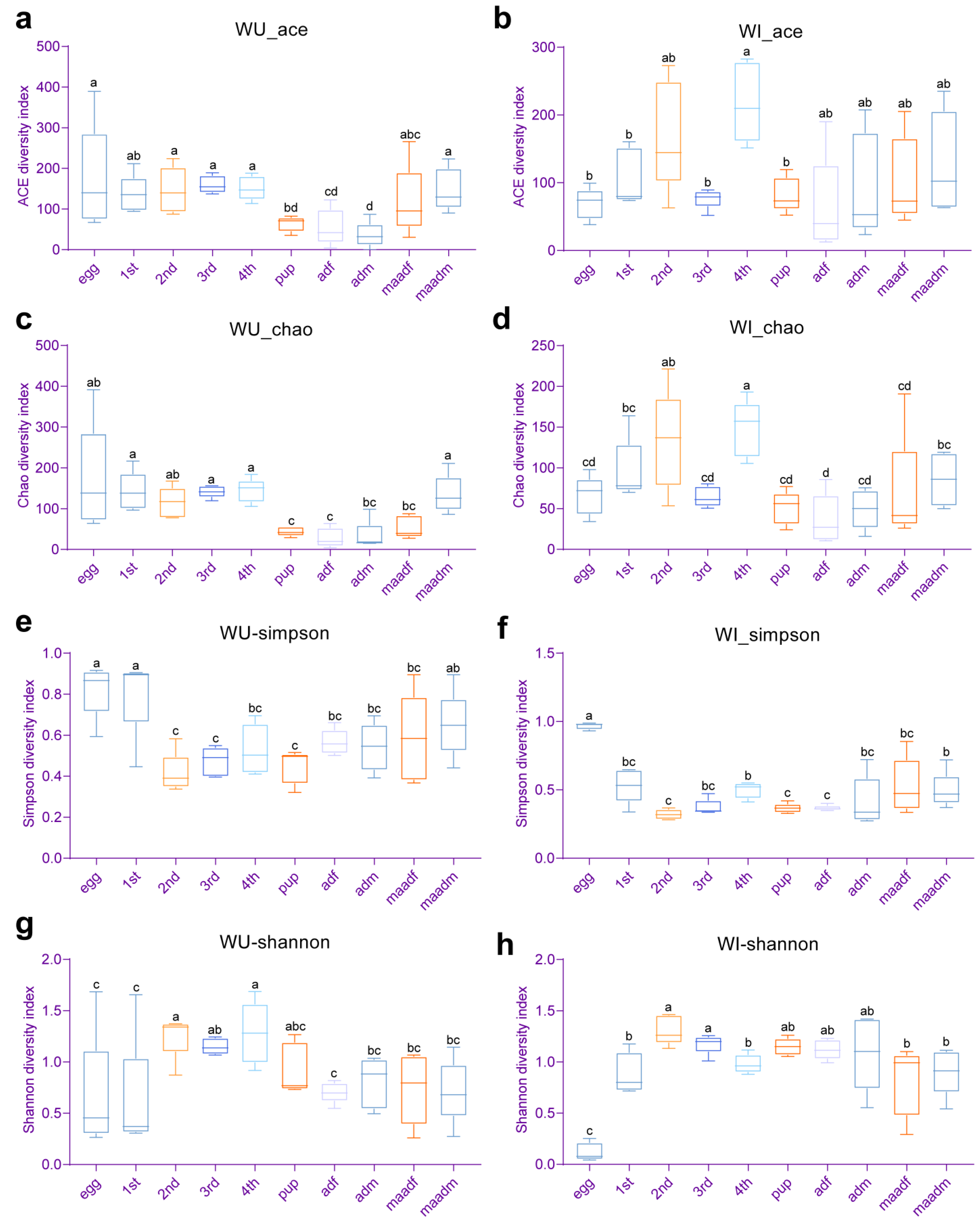
In WU P. xylostella, the species richness indices (Chao1 and ACE) of pupae, female adults (unmated and mated), and unmated male adults was significantly lower than those of other developmental stages (eggs, first–fourth instar larvae and mated male adults) (Figure 2a,c). The bacterial diversity (Simpson and Shannon indices) of eggs and first instar larvae were at a more similar level and significantly lower than those of second and fourth instar larvae in WU P. xylostella (Figure 2e,g). In WI P. xylostella, the species richness indices (Chao1 and ACE) of fourth instar larvae were significantly higher than those of all developmental stages except second instar larvae and mated male adults (egg, first and third instar larvae, female adults (unmated and mated), and unmated male adults) (Figure 2b,d). The Shannon index of WI P. xylostella eggs was significantly higher than that of all other developmental stages, and the Simpson index of WI P. xylostella eggs was significantly lower than that of all other developmental stages, indicating that the diversity of WI P. xylostella eggs was significantly lower than that of all other developmental stages (Figure 2f,h). We also made paired comparisons of the alpha diversity of WI and WU P. xylostella at each developmental stage using Student’s t-test, but none were statistically different.
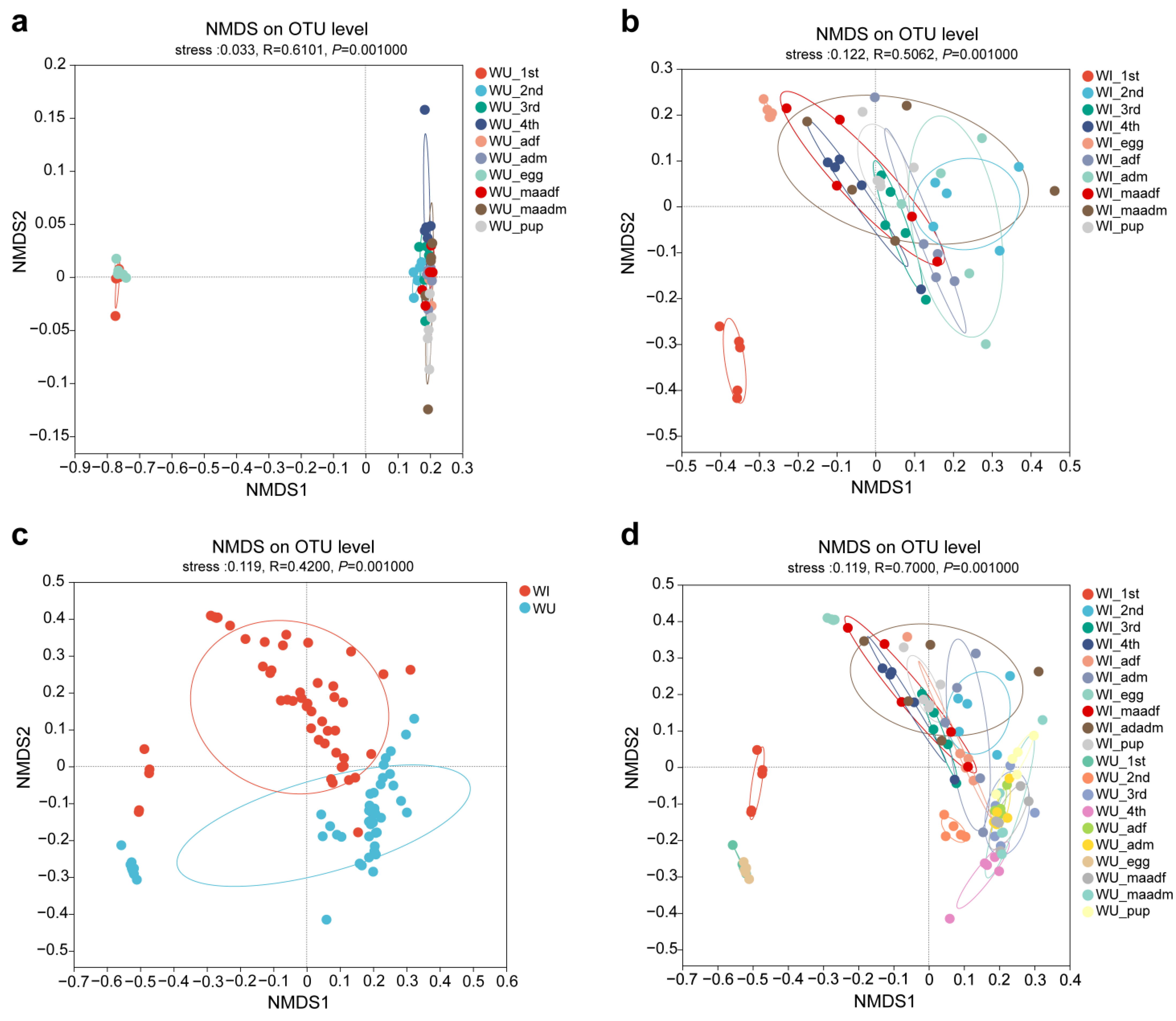
We analyzed the beta diversity to explore the similarity or difference of microbiota community composition between different groups of samples. NMDS plots showed significant differences when plotted by life stage regardless of the Wolbachia infection states (Figure 3a,b). The microbiota of eggs and first instar larvae were relatively closer in WU P. xylostella, while segregated from the other developmental stages. In WI P. xylostella, the microbiota of eggs were segregated from the other developmental stages. NMDS plots also showed significant differences when plotted by Wolbachia infection states at different developmental stages (Figure 3c,d).
3.6. Correlations between Dominant Genera and Predictive Functional Profiling
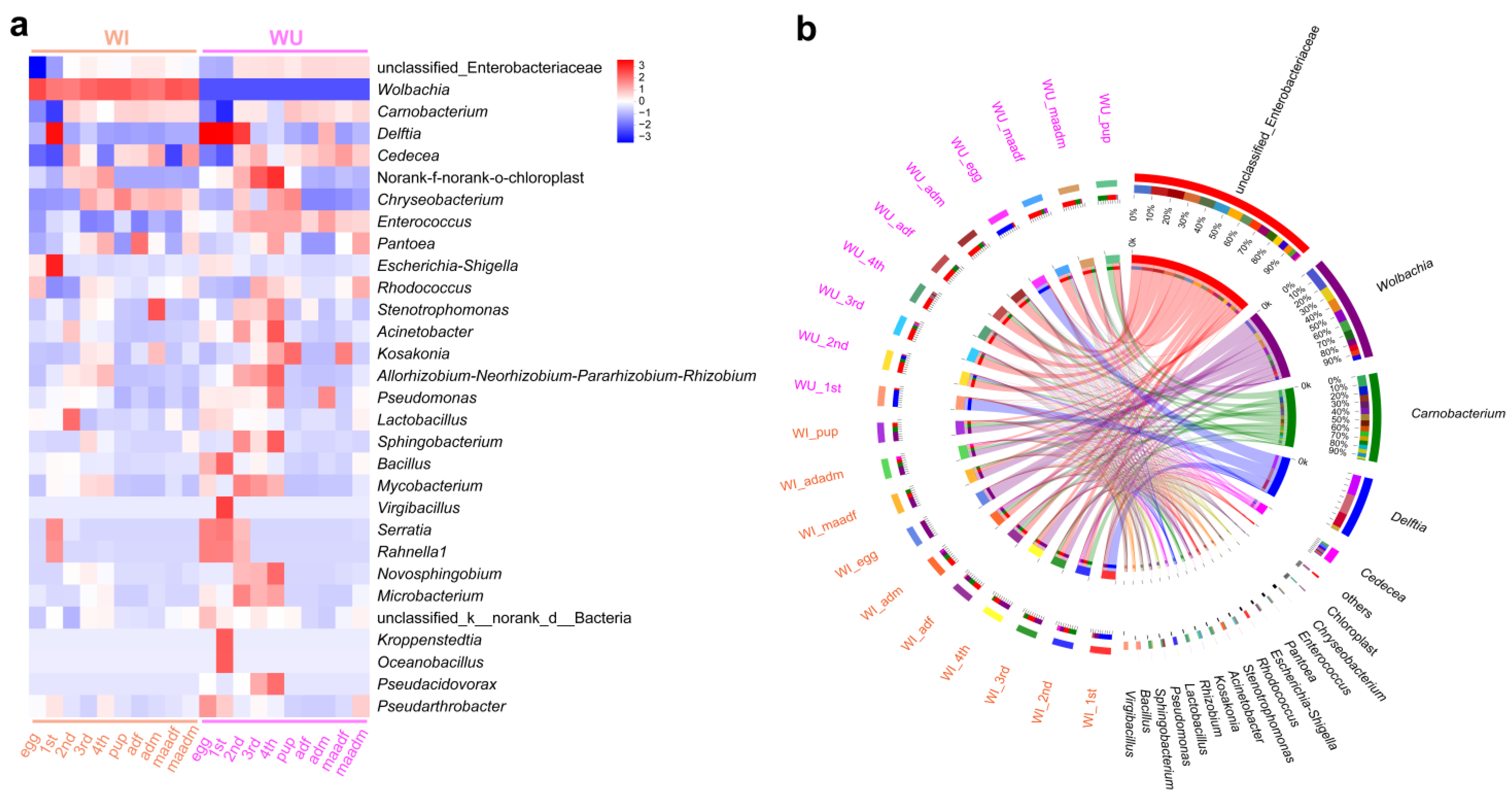
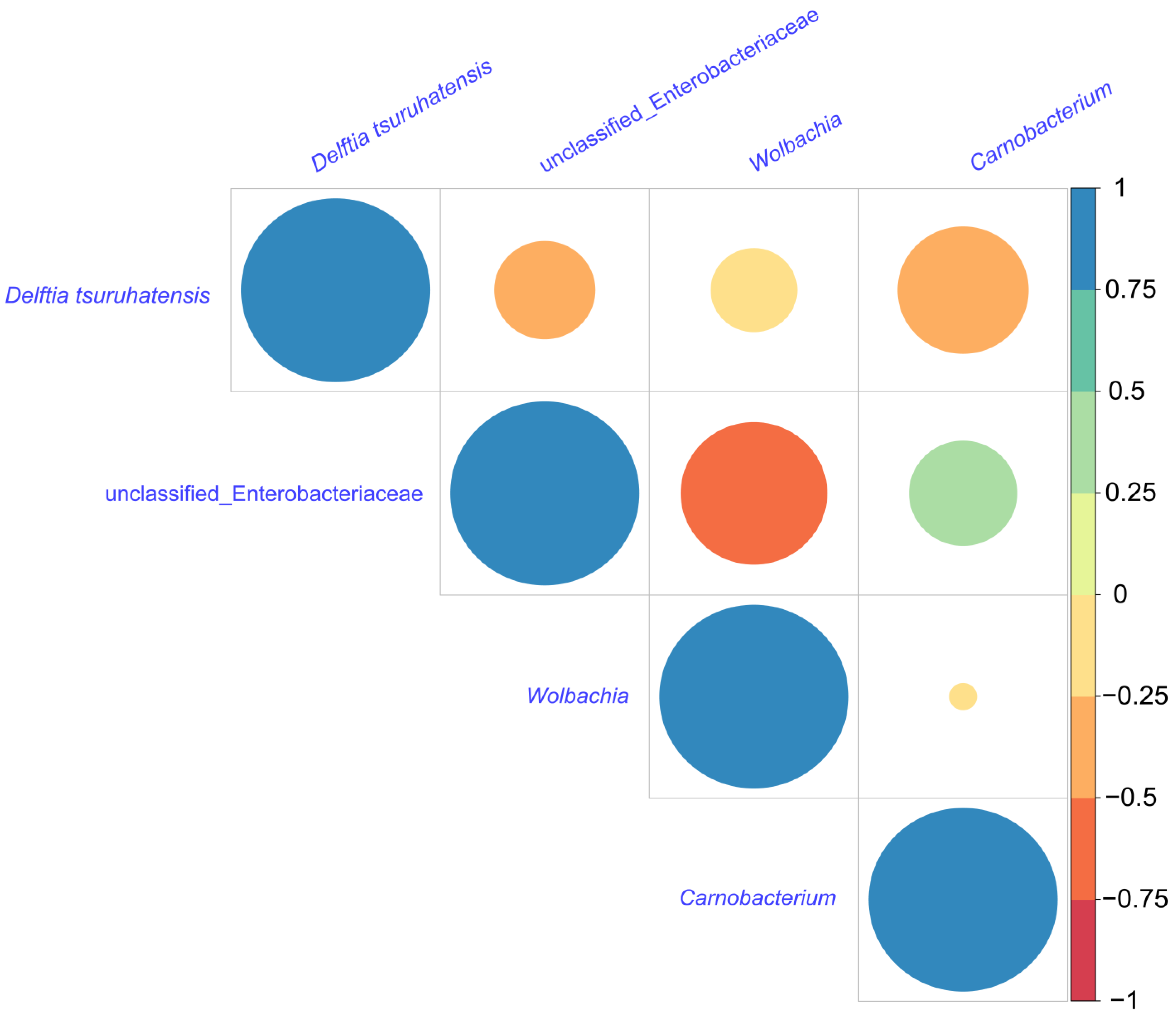
The community heat map demonstrated the relative abundance of the top 30 species in each sample at the species level (Figure 4a), with a total of 20 groups of 10 different developmental stages for WI and WU P. xylostella, and the relative abundance in each group was calculated using the mean value. By visualizing the circle plots, we also reflected the proportion of each dominant genus in different P. xylostella samples (Figure 4b). We also compared the relative abundance of dominant genera (an unclassified genus of Enterobacteriaceae, Wolbachia, Carnobacterium, and D. tsuruhatensis) at different developmental stages. The unclassified genus of Enterobacteriaceae was abundant in every developmental stage except the egg and first instar larval stages. Wolbachia was present only in WI P. xylostella and abundant in every developmental stage of WI P. xylostella, especially at the egg. Carnobacterium was also abundant at every developmental stage except the egg and first instar larval stages. The relative abundance of D. tsuruhatensis was high in the egg, first instar, and second instar larval stages of WU P. xylostella and largely undetectable in other developmental stages, while its abundance was high in the first instar larval stage of WI and largely undetectable in other developmental stages. We selected these four genera for correlation analysis, and the results of Spearman’s correlation coefficient showed that all three bacteria were negatively correlated with Wolbachia (Figure 5). The unclassified genus of Enterobacteriaceae, Wolbachia, and D. tsuruhatensis were all negatively correlated; Carnobacterium also positively correlated with Wolbachia and D. tsuruhatensis, but Carnobacterium positively correlated with the unclassified genus of Enterobacteriaceae.
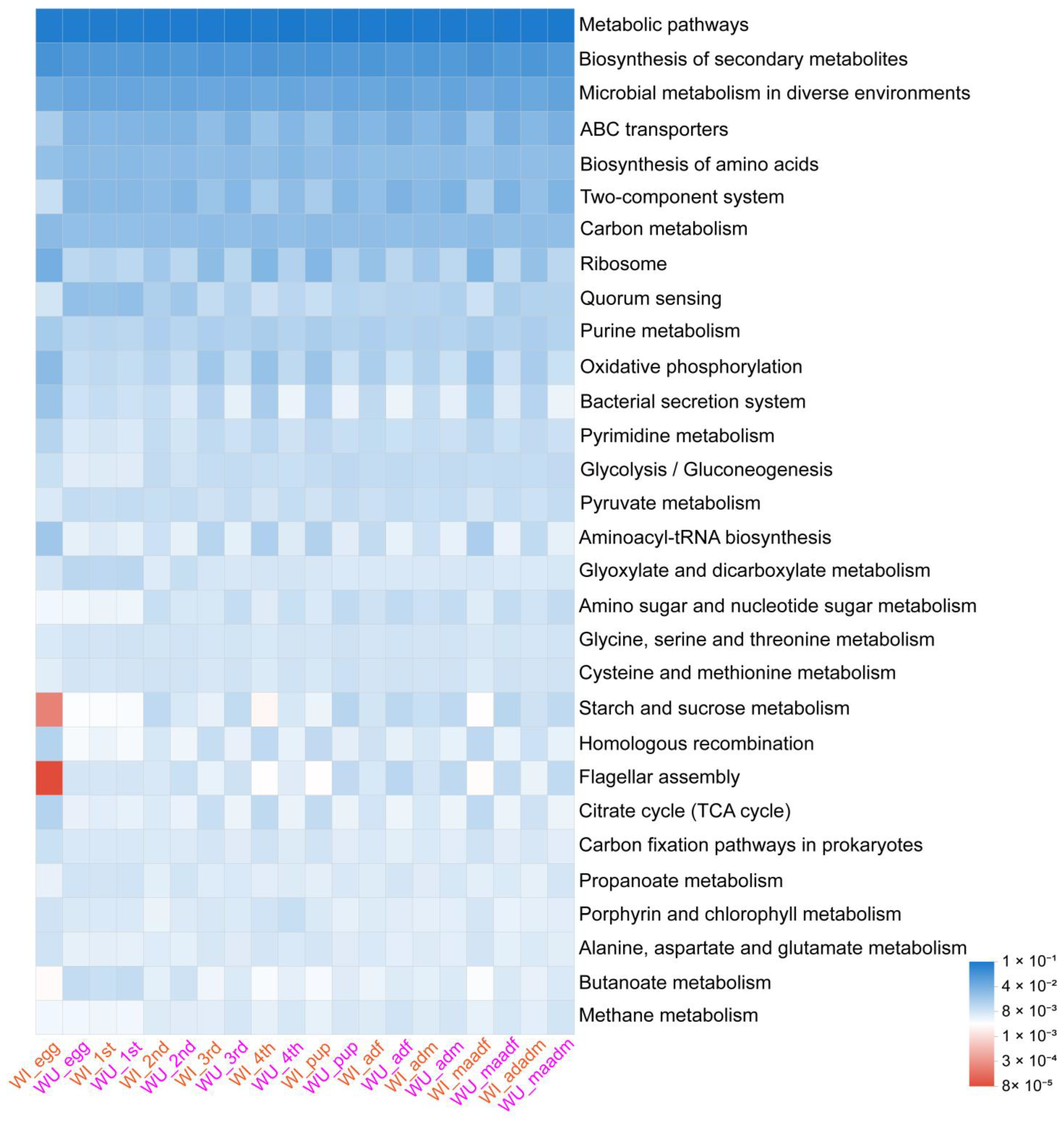
The results of the KEGGlevel3 analysis are shown in Figure 6. Wilcoxon test results showed that the relative abundance in various biologically important pathways in WI and WU P. xylostella differed significantly because of developmental stage (Tables S3 and S4).
4. Discussion
The diamondback moth is a notorious agricultural pest of cruciferous vegetables [35], and Wolbachia infection is known in some field populations, with the dominant strain identified as plutWB1 [39]. Similar to other Lepidoptera, P. xylostella undergoes complete metamorphosis, progressing through distinct life stages. This study sheds new light on the composition and diversity of P. xylostella’s microbiota across these life stages, examining both Wolbachia-infected and uninfected individuals.
Our investigation explored the microbial communities within P. xylostella across its life cycle. Recognizing that different microbes can possess specialized functions, the insect microbiome potentially adjusts to maintain optimal bacteria at each developmental stage [3]. Consistent with previous studies focusing on P. xylostella larvae [36,37], Proteobacteria and Firmicutes emerged as the dominant phyla across all stages and Wolbachia infection states in this work. These two phyla consistently dominate the microbiome of numerous lepidopteran insects, including Spodoptera frugiperda [51], Helicoverpa armigera [52], and Lymantria dispar [53]. Members of Firmicutes and Proteobacteria play crucial roles in key biological processes within insect hosts, including nutrient acquisition, energy absorption, gut homeostasis maintenance, and immunity [10,54,55].
Intriguingly, significant differences in microbiota composition were observed between egg and first-instar larvae of Wolbachia-infected and uninfected P. xylostella, compared to other stages. In WU individuals, D. tsuruhatensis dominated these early stages, reaching relative abundances of 90% in eggs and 89% in first-instar larvae. Notably, this dominance diminished dramatically in subsequent stages to barely detectable levels. Interestingly, Mereghetti et al. also detected Delftia in Indian meal moth, Plodia interpunctella, eggs, and early larvae, although at lower relative abundances than the present study [56]. A recent study suggests that D. tsuruhatensis in the mosquito gut produces a compound that inhibits parasite development [57]. Further research is needed to elucidate the specific function of bacterium in early life stages of P. xylostella. For later stages (second to fourth instar larvae, pupae, and adults), both infected and uninfected P. xylostella exhibited relatively high abundances of Enterobacteriaceae and Carnobacteriaceae. These families are relatively common gut microbes in insects and have been linked to growth and development, immunity, and agrochemical resistance [58]. Interestingly, Carnobacterium represents a major component of the microbiota in other moth species such as Ocnogyna loewii [59] and Thitarodes/Hepialus ghost moths [60], and may participate in insect metabolic activity [36].
Our analysis revealed significant differences in alpha diversity (species richness within samples) across the life stages of P. xylostella with minimal effect of sex, mating status, and Wolbachia infection. This finding contrasts with the earlier studies showing the role of these factors in shaping insect microbiomes [10,55,61,62]. Developmental stages, particularly early stages like eggs, are known to significantly affect bacterial diversity in many insects [21,23]. For example, eggs of fall armyworm, Spodoptera frugiperda, have markedly higher diversity than the larval, pupal, and adult stages [10]. Similar patterns have been observed in the beetle Octodonta nipae [61] and the moths Grapholita molesta [55], and Spodoptera littoralis [63]. However, our study contrasts with that trend. Notably, WI P. xylostella eggs exhibited significantly lower diversity than other stages, and the same pattern was observed in eggs and first instar larvae of WU P. xylostella. Insect hosts have different dominant microbiota at different life stages, which may reflect the different roles of different microbiota across particular life stages [63]. The reason for the low diversity in early P. xylostella stages deserves further investigation. While sex, mating status, and Wolbachia infection did not significantly influence alpha diversity, they influenced the relative abundance of some bacteria. This aligns with our previous study showing little effect of Wolbachia infection (or antibiotic treatment) on P. xylostella’s bacterial diversity [40]. Further analysis using beta diversity (species differences between samples) confirmed the results of alpha diversity and bacterial community composition analyses. Developmental stages significantly affected both diversity and composition, while sex, mating status, and Wolbachia infection had minimal influence on diversity but subtly affected certain bacterial proportions.
Wolbachia, the most ubiquitous insect endosymbiont, can dramatically manipulate host reproduction [15,26]. In P. xylostella, the dominant Wolbachia strain, plutWB1, biases the sex ratio towards females [38]. Our study unveils, for the first time, the intricate relationship between Wolbachia and the gut bacterial community across P. xylostella’s life cycle. We found Wolbachia persistently colonizing all developmental stages, consistently maintaining a high relative abundance. Notably, its abundance in WI P. xylostella eggs reached a remarkable 98.4%, mirroring the insect’s vertical transmission pattern and aligning with Wolbachia’s crucial role in manipulating host reproduction [26]. Wolbachia primarily spreads through egg cytoplasm, ensuring its passage to the next generation [64].
Moreover, Wolbachia manipulates sex ratios in Lepidoptera by targeting key sex-determination pathways, often employing either male-killing or feminization strategies [65,66]. In the oriental moth, O. furnacalis, Wolbachia specifically targets the host’s masculinizing gene to eliminate males [67]. Given the high Wolbachia abundance in P. xylostella eggs and the established egg-stage sex differentiation in this species [68,69], we propose a potentially intriguing scenario: Wolbachia’s manipulation at the egg stage might be driving the observed female-biased sex ratio [38].
Interestingly, Wolbachia abundance displayed a female-biased pattern, further aligning with its natural population transmission dynamics [64]. We also observed higher Wolbachia levels in mated adults compared to unmated ones. This aligns with Wolbachia’s goal: ultimate reproductive control of the host for its own population expansion [26]. Consequently, female bias in Wolbachia abundance is a common phenomenon in infected insect species [64,70].
While Wolbachia is suspected of manipulating the sex ratio in P. xylostella, other microbes like Spiroplasma, Cardinium, and RNA viruses can also have this effect in other insects [71,72]. Although no other sex-associated bacteria were detected in our study, the presence of undetected, potentially sex-influencing microbes warrants further exploration.
Furthermore, it has been suggested that interactions between microbes within P. xylostella, including plutWB1, could contribute to the sex ratio imbalance [38]. Wolbachia can engage in complex interactions with other microorganisms, both competitive and synergistic. For example, bacterial community composition differed significantly between Wolbachia-free and infected plant hopper, Laodelphax striatellus [5]. Similarly, Wolbachia significantly altered the gut microbiome of D. melanogaster and reduced Acetobacter abundance [73]. Our study also demonstrates a significant impact of Wolbachia on the P. xylostella microbiome.
We further analyzed correlations between Wolbachia and other dominant bacterial genera through abundance data. Interestingly, negative correlations were observed with an unclassified genus of Enterobacteriaceae, D. tsuruhatensis, and Carnobacterium. However, it is crucial to emphasize that these effects and correlations remain tentative. Our study utilized WU P. xylostella obtained by antibiotic treatment of WI individuals with nearly 100% infection rates [40]. To mitigate potential antibiotic effects, WU P. xylostella were reared for over 50 generations before sequencing. Additionally, at the 10th generation post-treatment, we confirmed that antibiotics no longer significantly affected the bacterial community compared to an untreated, genetically distinct population [40]. Despite these measures, further validation is necessary to fully elucidate Wolbachia’s specific effects on the bacterial community.
5. Conclusions
Our study unveils, for the first time, the dynamics of P. xylostella’s bacterial communities across life stage, Wolbachia infection, sex, and mating status. Bacterial diversity and composition shifted strikingly with developmental stage. While Wolbachia infection, sex, and mating status did not significantly affect diversity, they subtly influenced the relative abundance of specific bacterial genera. Notably, the dominant taxa (an unclassified genus of Enterobacteriaceae, Wolbachia, Carnobacterium, and D. tsuruhatensis) varied significantly across developmental stages, highlighting life stage as a key driver of microbiome structure. Intriguingly, Wolbachia displayed remarkable stability, maintaining high relative abundance throughout development, albeit with a peak in eggs. This finding suggests a potentially crucial role for Wolbachia at this early stage. Finally, we explored preliminary correlations between Wolbachia and other P. xylostella bacteria, providing a springboard for future research to delve deeper into the intricate host–bacteria relationships within this insect pest.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects15020132/s1, Table S1: Detailed information of microbiome diversity indices of all WI P. xylostella samples; Table S2: Detailed information of microbiome diversity indices of all WU P. xylostella samples; Table S3: The abundance test of the top 50 pathways enriched in KEGGL3 levels in WI P. xylostella at different developmental stages; Table S4: The abundance test of the top 50 pathways enriched in KEGGL3 levels in WU P. xylostella at different developmental stages.
Author Contributions
Conceptualization, X.Z. and S.Y.; methodology, X.Z., J.L. and A.H.; software, X.Z. and A.H.; formal analysis, X.Z. and J.L.; resources, X.Z., J.L. and A.H.; data curation, X.Z.; writing—original draft preparation, X.Z.; writing—review and editing, all authors; visualization, X.Z. and J.L.; supervision, S.Y., M.Y. and G.M.G.; project administration, S.Y., M.Y. and G.M.G.; funding acquisition, S.Y., M.Y. and G.M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Fujian Natural Science Fund for Distinguished Young Scholars (grant number 2022J06013) and Central Guided Local Science and Technology Development Project (grant number 2022L3087).
Data Availability Statement
The raw sequencing data are deposited in the NCBI Sequence Read. Archive (SRA) database with accession number PRJNA1068811.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Nair, S. Dynamics of Insect-Microbiome Interaction Influence Host and Microbial Symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Boyer, S.; Bezemer, T.M.; Lefort, M.C.; Dhami, M.K.; Biggs, E.; Groenteman, R.; Fowler, S.V.; Paynter, Q.; Verdecia Mogena, A.M.; et al. Impact of intraspecific variation in insect microbiomes on host phenotype and evolution. ISME J. 2023, 17, 1798–1807. [Google Scholar] [CrossRef] [PubMed]
- Janson, E.M.; Stireman, J.O., 3rd; Singer, M.S.; Abbot, P. Phytophagous insect-microbe mutualisms and adaptive evolutionary diversification. Evolution 2008, 62, 997–1012. [Google Scholar] [CrossRef]
- Duan, X.Z.; Sun, J.T.; Wang, L.T.; Shu, X.H.; Guo, Y.; Keiichiro, M.; Zhu, Y.X.; Bing, X.L.; Hoffmann, A.A.; Hong, X.Y. Recent infection by Wolbachia alters microbial communities in wild Laodelphax striatellus populations. Microbiome 2020, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Wang, Q.; Mu, X.; Chen, H.; Meng, Y.; Zhang, X.; Zheng, L.; Hu, X.; Zhai, Y.; Zheng, H. Strain-level analysis reveals the vertical microbial transmission during the life cycle of bumblebee. Microbiome 2021, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Li, T.P.; Zhou, C.Y.; Wang, M.K.; Zha, S.S.; Chen, J.; Bing, X.L.; Hoffmann, A.A.; Hong, X.Y. Endosymbionts Reduce Microbiome Diversity and Modify Host Metabolism and Fecundity in the Planthopper Sogatella furcifera. mSystems 2022, 7, e0151621. [Google Scholar] [CrossRef]
- Luo, X.; Fang, G.; Chen, K.; Song, Y.; Lu, T.; Tomberlin, J.K.; Zhan, S.; Huang, Y. A gut commensal bacterium promotes black soldier fly larval growth and development partly via modulation of intestinal protein metabolism. mBio 2023, 14, e0117423. [Google Scholar] [CrossRef]
- Harumoto, T.; Lemaitre, B. Male-killing toxin in a bacterial symbiont of Drosophila. Nature 2018, 557, 252–255. [Google Scholar] [CrossRef]
- Fu, J.; Wang, J.; Huang, X.; Guan, B.; Feng, Q.; Deng, H. Composition and diversity of gut microbiota across developmental stages of Spodoptera frugiperda and its effect on the reproduction. Front. Microbiol. 2023, 14, 1237684. [Google Scholar] [CrossRef]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Immune system stimulation by the native gut microbiota of honey bees. R. Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef] [PubMed]
- Arias-Rojas, A.; Frahm, D.; Hurwitz, R.; Brinkmann, V.; Iatsenko, I. Resistance to host antimicrobial peptides mediates resilience of gut commensals during infection and aging in Drosophila. Proc. Natl. Acad. Sci. USA 2023, 120, e2305649120. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, J.; Tian, X.; Toufeeq, S.; Huang, W. Immunometabolic regulation during the presence of microorganisms and parasitoids in insects. Front. Immunol. 2023, 14, 905467. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Fang, Y.; Hou, Y.; Shi, Z. The Gut Entomotype of Red Palm Weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Dryophthoridae) and Their Effect on Host Nutrition Metabolism. Front. Microbiol. 2017, 8, 2291. [Google Scholar] [CrossRef]
- Ju, J.F.; Bing, X.L.; Zhao, D.S.; Guo, Y.; Xi, Z.; Hoffmann, A.A.; Zhang, K.J.; Huang, H.J.; Gong, J.T.; Zhang, X.; et al. Wolbachia supplement biotin and riboflavin to enhance reproduction in planthoppers. ISME J. 2020, 14, 676–687. [Google Scholar] [CrossRef]
- Yuan, S.; Sun, Y.; Chang, W.; Zhang, J.; Sang, J.; Zhao, J.; Song, M.; Qiao, Y.; Zhang, C.; Zhu, M.; et al. The silkworm (Bombyx mori) gut microbiota is involved in metabolic detoxification by glucosylation of plant toxins. Commun. Biol. 2023, 6, 790. [Google Scholar] [CrossRef]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef]
- Feng, H.; Edwards, N.; Anderson, C.M.H.; Althaus, M.; Duncan, R.P.; Hsu, Y.C.; Luetje, C.W.; Price, D.R.G.; Wilson, A.C.C.; Thwaites, D.T. Trading amino acids at the aphid-Buchnera symbiotic interface. Proc. Natl. Acad. Sci. USA 2019, 116, 16003–16011. [Google Scholar] [CrossRef]
- Couret, J.; Huynh-Griffin, L.; Antolic-Soban, I.; Acevedo-Gonzalez, T.S.; Gerardo, N.M. Even obligate symbioses show signs of ecological contingency: Impacts of symbiosis for an invasive stinkbug are mediated by host plant context. Ecol. Evol. 2019, 9, 9087–9099. [Google Scholar] [CrossRef]
- LePage, D.P.; Metcalf, J.A.; Bordenstein, S.R.; On, J.; Perlmutter, J.I.; Shropshire, J.D.; Layton, E.M.; Funkhouser-Jones, L.J.; Beckmann, J.F.; Bordenstein, S.R. Prophage WO genes recapitulate and enhance Wolbachia-induced cytoplasmic incompatibility. Nature 2017, 543, 243–247. [Google Scholar] [CrossRef]
- Hammer, T.J.; McMillan, W.O.; Fierer, N. Metamorphosis of a butterfly-associated bacterial community. PLoS ONE 2014, 9, e86995. [Google Scholar] [CrossRef] [PubMed]
- Kowallik, V.; Mikheyev, A.S. Honey Bee Larval and Adult Microbiome Life Stages Are Effectively Decoupled with Vertical Transmission Overcoming Early Life Perturbations. mBio 2021, 12, e0296621. [Google Scholar] [CrossRef] [PubMed]
- Janke, R.S.; Kaftan, F.; Niehs, S.P.; Scherlach, K.; Rodrigues, A.; Svatoš, A.; Hertweck, C.; Kaltenpoth, M.; Flórez, L.V. Bacterial ectosymbionts in cuticular organs chemically protect a beetle during molting stages. ISME J. 2022, 16, 2691–2701. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Wagner, D.; Friedrich, M.W.; Wagner, B.; Brune, A. Axial dynamics, stability, and interspecies similarity of bacterial community structure in the highly compartmentalized gut of soil-feeding termites (Cubitermes spp.). Appl. Environ. Microbiol. 2003, 69, 6018–6024. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, X.; De Mandal, S.; Shakeel, M.; Hua, Y.; Shoukat, R.F.; Fu, D.; Jin, F. Gut microbiota mediate Plutella xylostella susceptibility to Bt Cry1Ac protoxin is associated with host immune response. Environ. Pollut. 2021, 271, 116271. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Hertig, M.; Wolbach, S.B. Studies on Rickettsia-Like Micro-Organisms in Insects. J. Med. Res. 1924, 44, 329. [Google Scholar]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef]
- Teixeira, L.; Ferreira, A.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e2. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.E.; Leong, Y.S.; O’Neill, S.L.; Johnson, K.N. Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PLoS Pathog. 2009, 5, e1000656. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Anbutsu, H.; Fukatsu, T. Asymmetrical interactions between Wolbachia and Spiroplasma endosymbionts coexisting in the same insect host. Appl. Environ. Microbiol. 2006, 72, 4805–4810. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.L.; Dodson, B.L.; Johnson, R.M.; Murdock, C.C.; Tsujimoto, H.; Suzuki, Y.; Patt, A.A.; Cui, L.; Nossa, C.W.; Barry, R.M.; et al. Native microbiome impedes vertical transmission of Wolbachia in Anopheles mosquitoes. Proc. Natl. Acad. Sci. USA 2014, 111, 12498–12503. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Ke, F.; You, S.; Wu, Z.; Liu, Q.; He, W.; Baxter, S.W.; Yuchi, Z.; Vasseur, L.; Gurr, G.M.; et al. Variation among 532 genomes unveils the origin and evolutionary history of a global insect herbivore. Nat. Commun. 2020, 11, 2321. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Gurr, G.M.; Vasseur, L.; Zheng, D.; Zhong, H.; Qin, B.; Lin, J.; Wang, Y.; Song, F.; Li, Y.; et al. Metagenomic Sequencing of Diamondback Moth Gut Microbiome Unveils Key Holobiont Adaptations for Herbivory. Front. Microbiol. 2017, 8, 663. [Google Scholar] [CrossRef]
- Xia, X.; Zheng, D.; Zhong, H.; Qin, B.; Gurr, G.M.; Vasseur, L.; Lin, H.; Bai, J.; He, W.; You, M. DNA sequencing reveals the midgut microbiota of diamondback moth, Plutella xylostella (L.) and a possible relationship with insecticide resistance. PLoS ONE 2013, 8, e68852. [Google Scholar] [CrossRef]
- Delgado, A.M.; Cook, J.M. Effects of a sex-ratio distorting endosymbiont on mtDNA variation in a global insect pest. BMC Evol. Biol. 2009, 9, 49. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, T.; He, A.; Zhang, L.; Li, J.; Li, T.; Miao, X.; You, M.; You, S. Diversity of Wolbachia infection and its influence on mitochondrial DNA variation in the diamondback moth, Plutella xylostella. Mol. Phylogenetics Evol. 2023, 182, 107751. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, L.; Li, J.; He, A.; You, M.; You, S. Effects of Antibiotic Treatment on the Development and Bacterial Community of the Wolbachia-Infected Diamondback Moth. Evol. Bioinform. Online 2023, 19, 11769343231175269. [Google Scholar] [CrossRef]
- Ledbetter, R.N.; Connon, S.A.; Neal, A.L.; Dohnalkova, A.; Magnuson, T.S. Biogenic mineral production by a novel arsenic-metabolizing thermophilic bacterium from the Alvord Basin, Oregon. Appl. Environ. Microbiol. 2007, 73, 5928–5936. [Google Scholar] [CrossRef]
- Srinivasan, S.; Hoffman, N.G.; Morgan, M.T.; Matsen, F.A.; Fiedler, T.L.; Hall, R.W.; Ross, F.J.; McCoy, C.O.; Bumgarner, R.; Marrazzo, J.M.; et al. Bacterial communities in women with bacterial vaginosis: High resolution phylogenetic analyses reveal relationships of microbiota to clinical criteria. PLoS ONE 2012, 7, e37818. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Molina, Y.; Marulanda-Moreno, S.M.; Saldamando-Benjumea, C.; Junca, H.; Moreno-Herrera, C.X.; Cadavid-Restrepo, G. Microbiome analysis of Spodoptera frugiperda (Lepidoptera, Noctuidae) larvae exposed to Bacillus thuringiensis (Bt) endotoxins. PeerJ 2023, 11, e15916. [Google Scholar] [CrossRef]
- Xiang, H.; Wei, G.F.; Jia, S.; Huang, J.; Miao, X.X.; Zhou, Z.; Zhao, L.P.; Huang, Y.P. Microbial communities in the larval midgut of laboratory and field populations of cotton bollworm (Helicoverpa armigera). Can. J. Microbiol. 2006, 52, 1085–1092. [Google Scholar] [CrossRef]
- Broderick, N.A.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Colston, T.J.; Jackson, C.R. Microbiome evolution along divergent branches of the vertebrate tree of life: What is known and unknown. Mol. Ecol. 2016, 25, 3776–3800. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, S.; Yang, X.; Cheng, J.; Wei, H.; Li, Z.; Michaud, J.P.; Liu, X. Variability of Gut Microbiota across the Life Cycle of Grapholita molesta (Lepidoptera: Tortricidae). Front. Microbiol. 2020, 11, 1366. [Google Scholar] [CrossRef] [PubMed]
- Mereghetti, V.; Chouaia, B.; Limonta, L.; Locatelli, D.P.; Montagna, M. Evidence for a conserved microbiota across the different developmental stages of Plodia interpunctella. Insect Sci. 2019, 26, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Rodrigues, J.; Bilgo, E.; Tormo, J.R.; Challenger, J.D.; De Cozar-Gallardo, C.; Pérez-Victoria, I.; Reyes, F.; Castañeda-Casado, P.; Gnambani, E.J.; et al. Delftia tsuruhatensis TC1 symbiont suppresses malaria transmission by anopheline mosquitoes. Science 2023, 381, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.A.; Vennison, S.J.; Sankar, S.G.; Prabhu, D.I.; Vasan, P.T.; Raghuraman, T.; Geoffrey, C.J.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Berman, T.S.; Laviad-Shitrit, S.; Lalzar, M.; Halpern, M.; Inbar, M. Cascading effects on bacterial communities: Cattle grazing causes a shift in the microbiome of a herbivorous caterpillar. ISME J. 2018, 12, 1952–1963. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zheng, X.; Long, H.; Rao, Z.; Cao, L.; Han, R. Gut Bacterial and Fungal Communities of the Wild and Laboratory-Reared Thitarodes Larvae, Host of the Chinese Medicinal Fungus Ophiocordyceps sinensis on Tibetan Plateau. Insects 2021, 12, 327. [Google Scholar] [CrossRef]
- Ali, H.; Muhammad, A.; Sanda, N.B.; Huang, Y.; Hou, Y. Pyrosequencing Uncovers a Shift in Bacterial Communities Across Life Stages of Octodonta nipae (Coleoptera: Chrysomelidae). Front. Microbiol. 2019, 10, 466. [Google Scholar] [CrossRef]
- Tang, X.; Adler, P.H.; Vogel, H.; Ping, L. Gender-specific bacterial composition of black flies (Diptera: Simuliidae). FEMS Microbiol. Ecol. 2012, 80, 659–670. [Google Scholar] [CrossRef]
- Chen, B.; Teh, B.S.; Sun, C.; Hu, S.; Lu, X.; Boland, W.; Shao, Y. Biodiversity and Activity of the Gut Microbiota across the Life History of the Insect Herbivore Spodoptera littoralis. Sci. Rep. 2016, 6, 29505. [Google Scholar] [CrossRef]
- Kaur, R.; Shropshire, J.D.; Cross, K.L.; Leigh, B.; Mansueto, A.J.; Stewart, V.; Bordenstein, S.R.; Bordenstein, S.R. Living in the endosymbiotic world of Wolbachia: A centennial review. Cell Host Microbe 2021, 29, 879–893. [Google Scholar] [CrossRef]
- Sugimoto, T.N.; Fujii, T.; Kayukawa, T.; Sakamoto, H.; Ishikawa, Y. Expression of a doublesex homologue is altered in sexual mosaics of Ostrinia scapulalis moths infected with Wolbachia. Insect Biochem. Mol. Biol. 2010, 40, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, T.N.; Kayukawa, T.; Shinoda, T.; Ishikawa, Y.; Tsuchida, T. Misdirection of dosage compensation underlies bidirectional sex-specific death in Wolbachia-infected Ostrinia scapulalis. Insect Biochem. Mol. Biol. 2015, 66, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Fukui, T.; Kawamoto, M.; Shoji, K.; Kiuchi, T.; Sugano, S.; Shimada, T.; Suzuki, Y.; Katsuma, S. The Endosymbiotic Bacterium Wolbachia Selectively Kills Male Hosts by Targeting the Masculinizing Gene. PLoS Pathog. 2015, 11, e1005048. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Liu, Z.; Xu, J.; Li, X.; Bi, H.; Andongma, A.A.; Niu, C.; Huang, Y. Mutation of doublesex induces sex-specific sterility of the diamondback moth Plutella xylostella. Insect Biochem. Mol. Biol. 2019, 112, 103180. [Google Scholar] [CrossRef] [PubMed]
- Harvey-Samuel, T.; Norman, V.C.; Carter, R.; Lovett, E.; Alphey, L. Identification and characterization of a Masculinizer homologue in the diamondback moth, Plutella xylostella. Insect Mol. Biol. 2020, 29, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.; Sullivan, W. The cellular lives of Wolbachia. Nat. Rev. Microbiol. 2023, 21, 750–766. [Google Scholar] [CrossRef] [PubMed]
- Duplouy, A.; Hornett, E.A. Uncovering the hidden players in Lepidoptera biology: The heritable microbial endosymbionts. PeerJ 2018, 6, e4629. [Google Scholar] [CrossRef] [PubMed]
- Massey, J.H.; Newton, I.L.G. Diversity and function of arthropod endosymbiont toxins. Trends Microbiol. 2022, 30, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Simhadri, R.K.; Fast, E.M.; Guo, R.; Schultz, M.J.; Vaisman, N.; Ortiz, L.; Bybee, J.; Slatko, B.E.; Frydman, H.M. The Gut Commensal Microbiome of Drosophila melanogaster Is Modified by the Endosymbiont Wolbachia. mSphere 2017, 2, e00287-17. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Taxonomic composition and relative abundance of bacteria with P. xylostella across different populations (WI and WU) and developmental stages. (a) Relative abundance of phyla shifts of WU P. xylostella during the developmental stages as shown in a stream graph. (b) Relative abundance of phyla shifts of WI P. xylostella during the developmental stages as shown in a stream graph. (c) Relative phylotype-level abundance profiles for the WI and WU P. xylostella samples.
Figure 1.
Taxonomic composition and relative abundance of bacteria with P. xylostella across different populations (WI and WU) and developmental stages. (a) Relative abundance of phyla shifts of WU P. xylostella during the developmental stages as shown in a stream graph. (b) Relative abundance of phyla shifts of WI P. xylostella during the developmental stages as shown in a stream graph. (c) Relative phylotype-level abundance profiles for the WI and WU P. xylostella samples.

Figure 2.
Comparison of alpha diversity of bacterial communities between different developmental stages of WI and WU P. xylostella. ACE diversity index of (a) WU and (b) WI P. xylostella. Chao1 diversity index of (c) WU and (d) WI P. xylostella. Simpson diversity index of (e) WU and (f) WI P. xylostella. Shannon diversity index of (g) WU and (h) WI P. xylostella. Error bars indicate SEM. Means with the same letters do not differ significantly (one-way ANOVA, p > 0.05) and means with the different letters differ significantly (one-way ANOVA, p < 0.05) (a, b, c and d).
Figure 2.
Comparison of alpha diversity of bacterial communities between different developmental stages of WI and WU P. xylostella. ACE diversity index of (a) WU and (b) WI P. xylostella. Chao1 diversity index of (c) WU and (d) WI P. xylostella. Simpson diversity index of (e) WU and (f) WI P. xylostella. Shannon diversity index of (g) WU and (h) WI P. xylostella. Error bars indicate SEM. Means with the same letters do not differ significantly (one-way ANOVA, p > 0.05) and means with the different letters differ significantly (one-way ANOVA, p < 0.05) (a, b, c and d).

Figure 3.
Microbiota compositions of P. xylostella shift along the developmental stages and Wolbachia infections. Bray–Curtis distance: (a) the developmental stages of WU P. xylostella; (b) the developmental stages of WI P. xylostella; (c) all samples of WI and WU P. xylostella; (d) the developmental stages of WI and WU P. xylostella. Colors correspond to different groups, as shown in the legend.
Figure 3.
Microbiota compositions of P. xylostella shift along the developmental stages and Wolbachia infections. Bray–Curtis distance: (a) the developmental stages of WU P. xylostella; (b) the developmental stages of WI P. xylostella; (c) all samples of WI and WU P. xylostella; (d) the developmental stages of WI and WU P. xylostella. Colors correspond to different groups, as shown in the legend.

Figure 4.
Major taxa of WI and WU P. xylostella during development. (a) Heatmap shows the top 30 taxa from samples. (b) Circos plot shows the proportion of the dominant bacteria in different samples.
Figure 4.
Major taxa of WI and WU P. xylostella during development. (a) Heatmap shows the top 30 taxa from samples. (b) Circos plot shows the proportion of the dominant bacteria in different samples.

Figure 5.
Analysis of differential bacteria. Heatmap of the pairwise Spearman correlation coefcients of four dominant bacteria. Positive correlations are displayed as cyan to blue (0 to 1) gradients, and negative correlations are displayed as yellow to red (0 to −1) gradients.
Figure 5.
Analysis of differential bacteria. Heatmap of the pairwise Spearman correlation coefcients of four dominant bacteria. Positive correlations are displayed as cyan to blue (0 to 1) gradients, and negative correlations are displayed as yellow to red (0 to −1) gradients.

Figure 6.
Heat-map showing the putative functional profiling. The different color pattern indicates the relative abundance of bacterial operational taxonomic units (OTUs) involved in various biological functions.
Figure 6.
Heat-map showing the putative functional profiling. The different color pattern indicates the relative abundance of bacterial operational taxonomic units (OTUs) involved in various biological functions.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhu, X.; Li, J.; He, A.; Gurr, G.M.; You, M.; You, S. Developmental Shifts in the Microbiome of a Cosmopolitan Pest: Unraveling the Role of Wolbachia and Dominant Bacteria. Insects 2024, 15, 132. https://doi.org/10.3390/insects15020132
AMA Style
Zhu X, Li J, He A, Gurr GM, You M, You S. Developmental Shifts in the Microbiome of a Cosmopolitan Pest: Unraveling the Role of Wolbachia and Dominant Bacteria. Insects. 2024; 15(2):132. https://doi.org/10.3390/insects15020132
Chicago/Turabian StyleZhu, Xiangyu, Jinyang Li, Ao He, Geoff M. Gurr, Minsheng You, and Shijun You. 2024. "Developmental Shifts in the Microbiome of a Cosmopolitan Pest: Unraveling the Role of Wolbachia and Dominant Bacteria" Insects 15, no. 2: 132. https://doi.org/10.3390/insects15020132
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.