
Identification of a Male-Produced Aggregation Sex Pheromone in Rosalia batesi, an Endemic Japanese Longhorn Beetle


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Insect-Produced Volatiles
2.2. Chemicals
2.3. Analysis of Insect-Produced Compounds
2.4. Field Bioassays of the Synthetic Pheromone
2.5. Statistical Analysis
3. Results
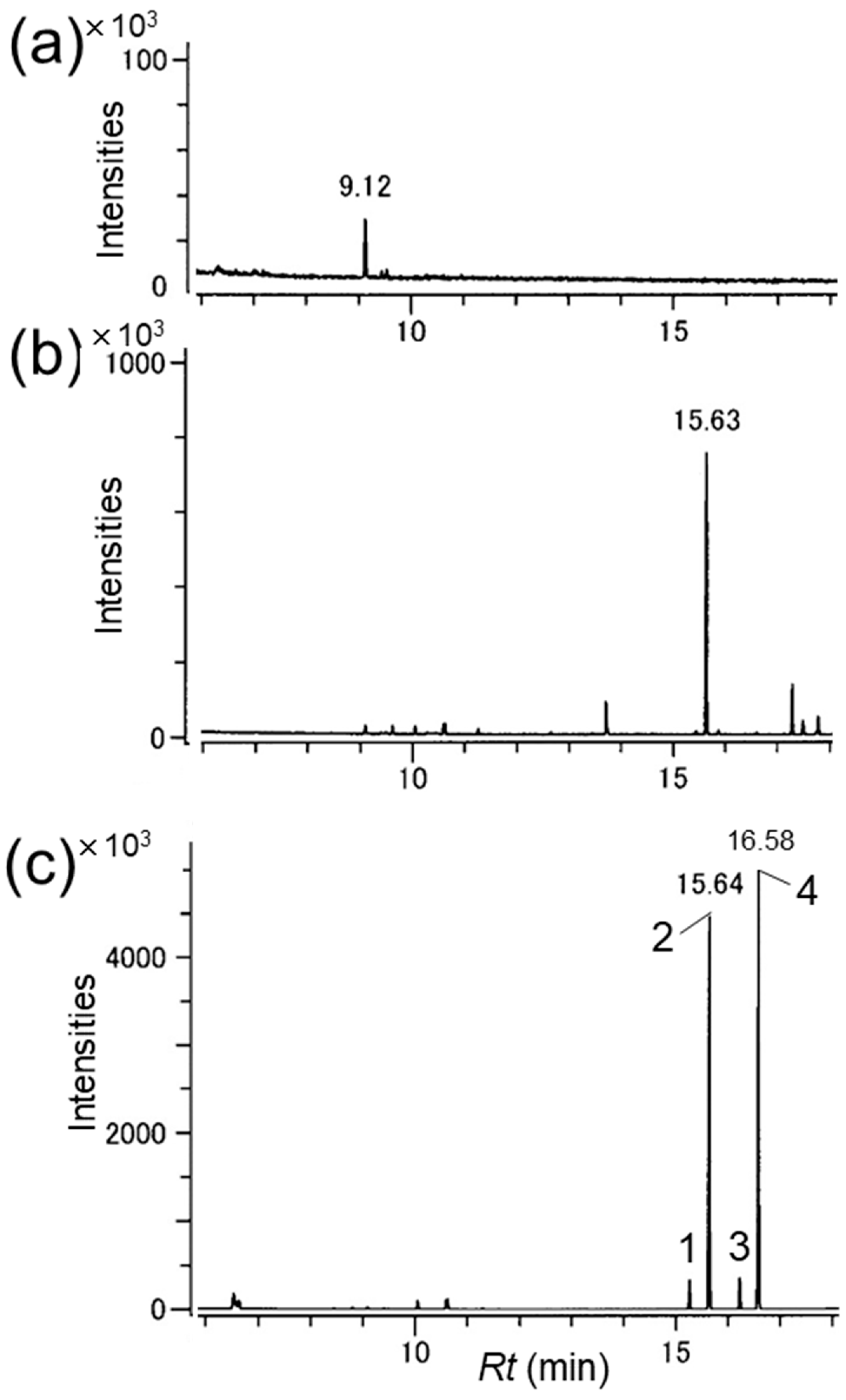
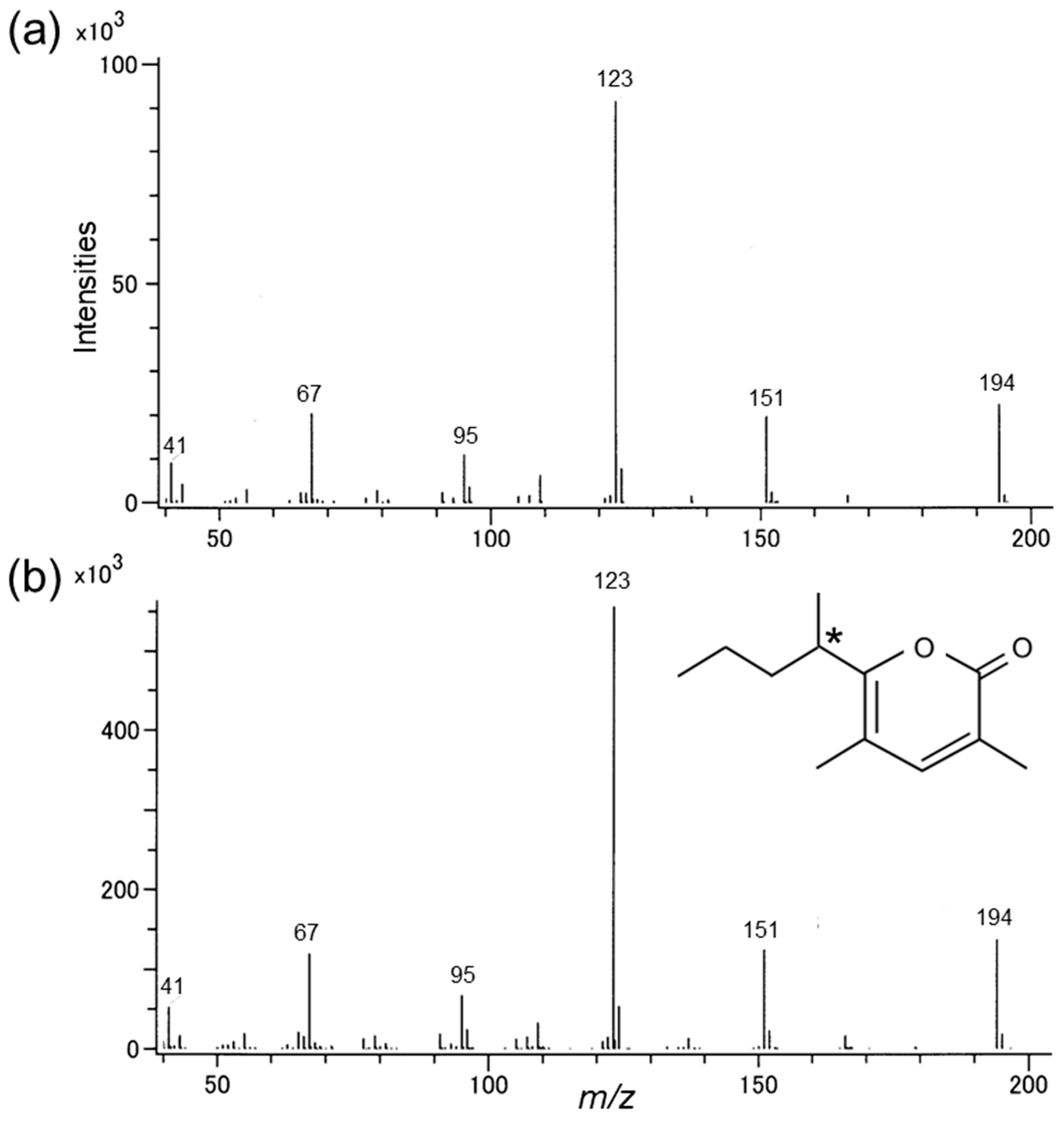
3.1. Identification of the Male-Produced Volatiles
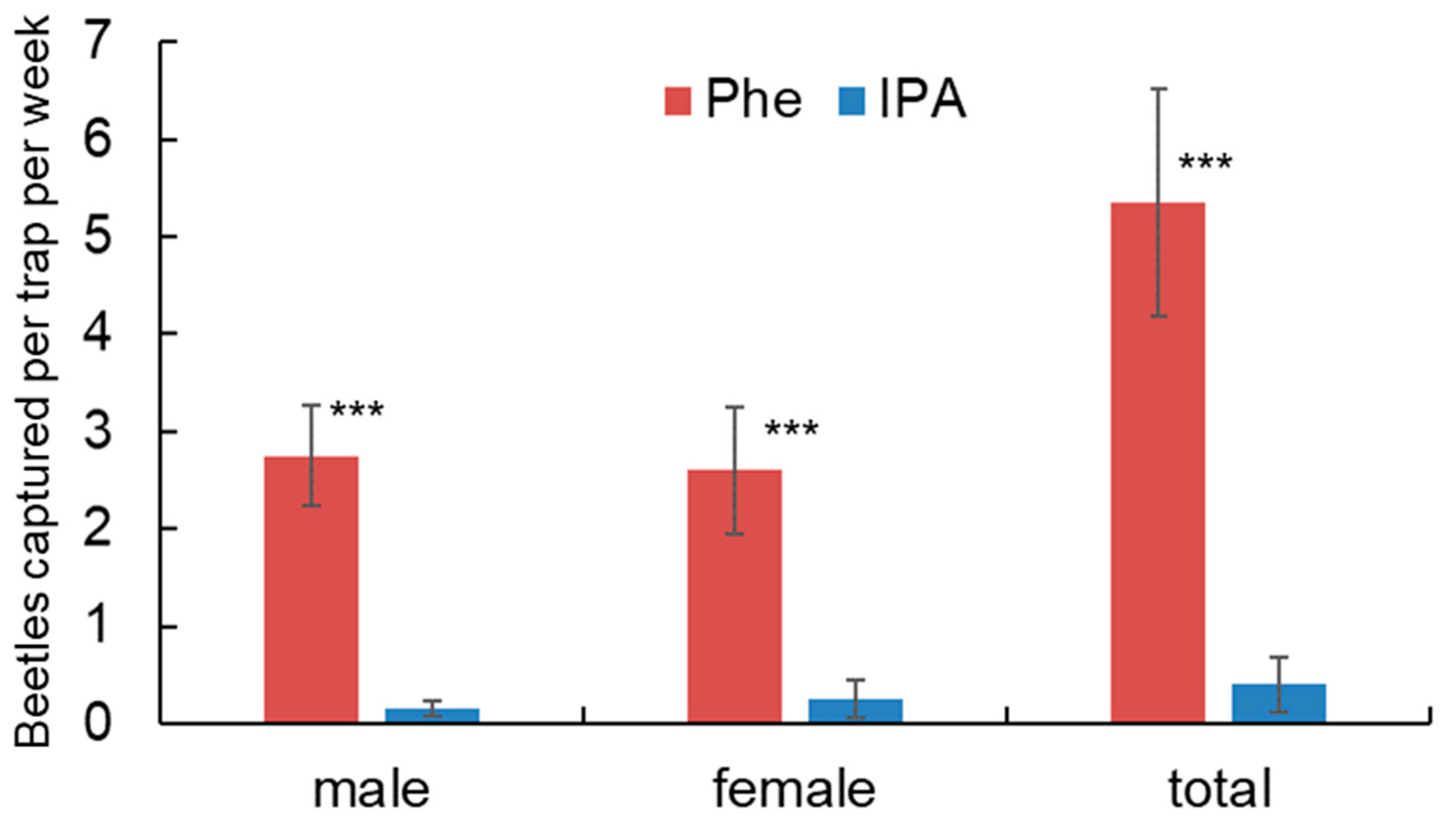
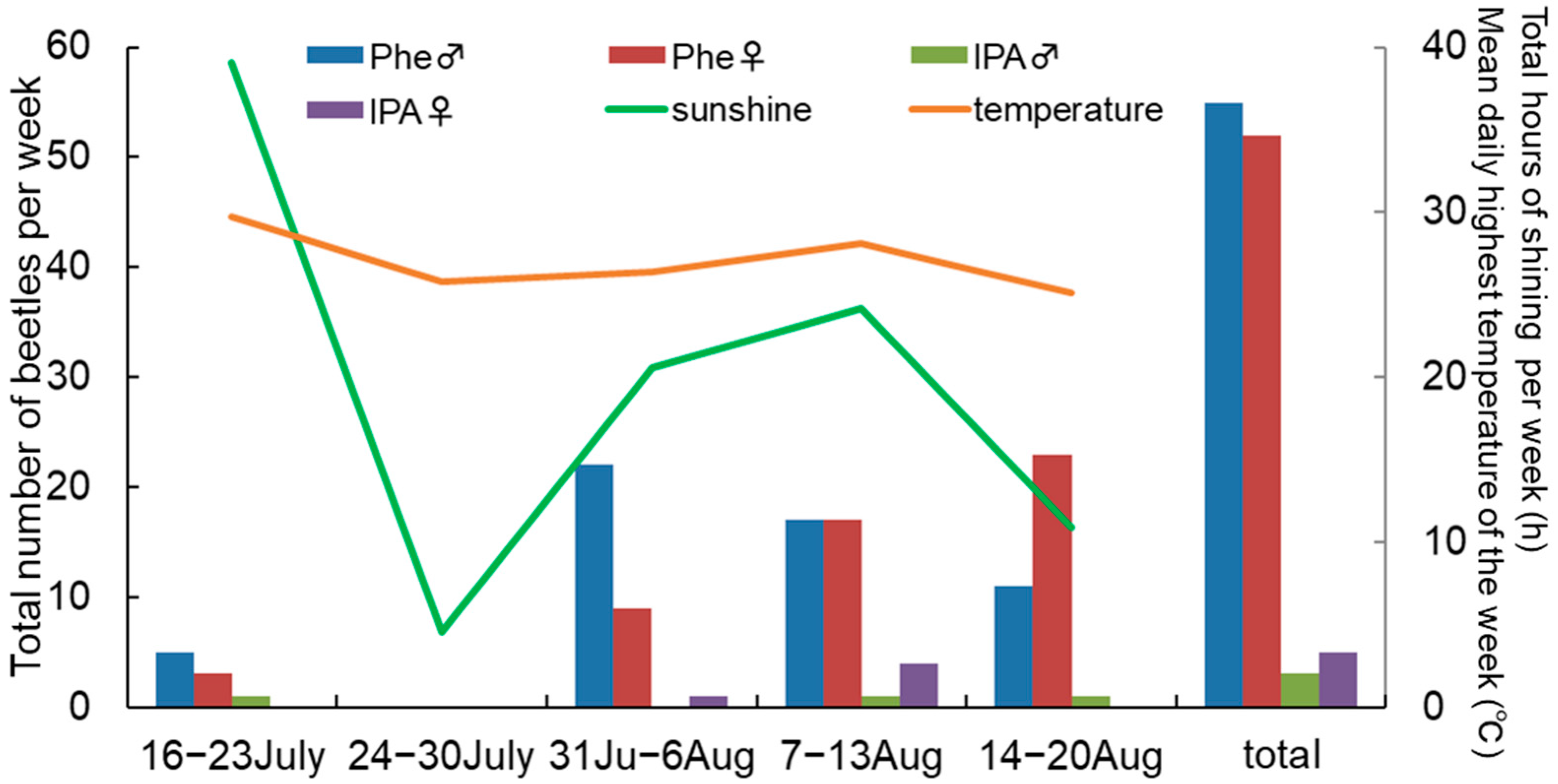
3.2. Field Bioassays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takakuwa, M. World’s Rosalia. In The Rosalia World; Takakuwa, M., Ed.; Mushi-Sha: Tokyo, Japan, 1998; pp. 35–100. (In Japanese) [Google Scholar]
- Takakuwa, M. Distributional expansion of a cerambycine longicorn beetle, R. batesi, in southern Fossa Magna area, central Japan, with special references to its surroundings. Gekkan-Mushi 2000, 353, 2–17. (In Japanese) [Google Scholar]
- Kanno, Y.; Ito, Y. Record of Rosalia batesi (Coleoptera: Cerambycidae) in Yokohama-city. Gekkan-Mushi 2000, 356, 46. (In Japanese) [Google Scholar]
- Hinoki, T. Records of Rosalia batesi (Coleoptera: Cerambycidae) from Tama-city, Tokyo. Gekkan-Mushi 2001, 370, 40. (In Japanese) [Google Scholar]
- Kanno, Y. Additional records of Rosalia batesi (Coleoptera: Cerambycidae) in Yokohama-city. Gekkan-Mushi 2001, 361, 46–47. (In Japanese) [Google Scholar]
- Kawada, K. Records of Rosalia batesi (Coleoptera: Cerambycidae) in Kawasaki-city in Kanagawa Prefecture. Gekkan-Mushi 2001, 361, 46. (In Japanese) [Google Scholar]
- Saito, K. Additional records of Rosalia batesi (Coleoptera: Cerambycidae) and Parnassius citrinarius (Lepidoptera: Papilionidae) in expanding distribution area. Gekkan-Mushi 2001, 366, 48. (In Japanese) [Google Scholar]
- Ichihashi, H. Distribution of Rosalia batesi (Coleoptera: Cerambycidae) in Sagamihara-city in Kanagawa Prefecture. Gekkan-Mushi 2005, 408, 38–40. (In Japanese) [Google Scholar]
- Miyauchi, H. Record of Rosalia batesi (Coleoptera: Cerambycidae) in Sodegaura-city in Chiba Prefecture. Gekkan-Mushi 2007, 431, 42. (In Japanese) [Google Scholar]
- Makihara, H.; Nakamura, N. Records of Rosalia batesi (Coleoptera: Cerambycidae) in Shimodate-city in Ibaraki Prefecture. Gekkan-Mushi 2009, 466, 35. (In Japanese) [Google Scholar]
- Dejima, T. Record of Rosalia batesi (Coleoptera: Cerambycidae) from Kagawa Prefecture. Gekkan-Mushi 2009, 455, 46. (In Japanese) [Google Scholar]
- Suda, H. Record of Rosalia batesi (Coleoptera: Cerambycidae) in Kawagoe-city in Saitama Prefecture. Gekkan-Mushi 2009, 466, 34–35. (In Japanese) [Google Scholar]
- Takahashi, K. Records of Rosalia batesi (Coleoptera: Cerambycidae) in Hiratsuka-city in Kanagawa Prefecture. Kanagawa Pref. Plant Prot. 2010, 171, 70–71. (In Japanese) [Google Scholar]
- Hirano, M.; Higashijima, K.; Koga, Y.; Ochi, T. Distributional expansion of Rosalia batesi in central Kinki district, Japan. Gekkan-Mushi 2017, 556, 44–47. (In Japanese) [Google Scholar]
- Kojima, K.; Nakamura, S. Food plants of cerambycid beetles (Cerambycidae, Coleoptera) in Japan. Hiba Soc. Nat. Hist. Shobara 1986, 11, 336. (In Japanese) [Google Scholar]
- Yamaguchi, K. A wooden house damaged by Rosalia batesi Harold (Coleoptera: Cerambycidae). House Househ. Insect Pests 1998, 20, 79–81. (In Japanese) [Google Scholar]
- Iwata, R.; Aoki, M.; Nozaki, T.; Yamaguchi, M. Some notes on the biology of a hardwood-log-boring beetle, Rosalia batesi Harold (Coleoptera: Cerambycidae), with special reference to its occurrences in a building and a suburban lumberyard. Jpn. J. Environ. Entomol. Zool. 1998, 9, 83–97. [Google Scholar]
- Kiriyama, S.; Iwata, R. Two cases of indoor occurrences of Rosalia batesi Harold (Coleoptera: Carambycidae) emerged from lumbers at Shonan Campus, Nihon University, Fujisawa, Japan. Urban Pest Manag. 2013, 3, 19–22. (In Japanese) [Google Scholar]
- Kiriyama, S.; Iwata, R.; Fukaya, M.; Hoshino, Y.; Yamanaka, Y. Mating behavior of Rosalia batesi (Coleoptera: Cerambycidae) is mediated by male-produced sex pheromones. Insects 2018, 9, 48. [Google Scholar] [CrossRef]
- Sama, G. Subfamily Cerambicinae Latreille, 1802. In Atlas of the Cerambycidae of Europe and the Mediterranean Area; Sama, G., Ed.; Kabourek: Zlin, Czech Republic, 2002; Volume 1, p. 54. [Google Scholar]
- Lachat, T.; Ecker, K.; Duelli, P.; Wermelinger, B. Population trends of Rosalia alpina (L.) in Switzerland: A lasting turnaround? J. Insect Conserv. 2013, 17, 653–662. [Google Scholar] [CrossRef]
- Michalcewicz, J.; Ciach, M. Current distribution of the Rosalia longicorn Rosalia alpina (Linnaeus, 1758) (Coleoptera: Cerambycidae) in Poland. Pol. J. Entomol. 2015, 84, 9–20. [Google Scholar] [CrossRef]
- Linsley, E.G. The Cerambycidae of North America: Part V. Taxonomy and classification of the subfamily Cerambycinae, tribes Callichromini through Ancylocerini. Univ. Calif. Publ. Entomol. 1964, 22, 4–6. [Google Scholar]
- Linsley, E.G. The banded alder beetle in natural and urban environments (Coleoptera: Cerambycidae). Pan-Pac. Entomol. 1995, 71, 133–134. [Google Scholar]
- Žunič Kosi, A.; Zou, Y.; Hoskovec, M.; Vrezec, A.; Stritih, N.; Millar, J.G. Novel, male-produced aggregation pheromone of the cerambycid beetle Rosalia alpina, a priority species of European conservation concern. PLoS ONE 2017, 12, e0183279. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.M.; Millar, J.G.; McElfresh, J.S.; Swift, I.P.; Barbour, J.D.; Hanks, L.M. Male-produced aggregation pheromone of the cerambycid beetle Rosalia funebris. J. Chem. Ecol. 2009, 35, 96–103. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species. Version 2022.2. 2023. Available online: www.iucnredlist.org (accessed on 6 October 2023).
- Komonen, A.; Jonsell, M.; Ranius, T. Red-listing saproxylic beetles in Fennoscandia: Current status and future perspectives. Endanger. Species Res. 2008, 6, 149–154. [Google Scholar] [CrossRef]
- Drag, L.; Hauck, D.; Pokluda, P.; Zimmermann, K.; Cizek, L. Demography and dispersal ability of a threatened saproxylic beetle: A mark-recapture study of the Rosalia longicorn (Rosalia alpina). PLoS ONE 2011, 6, e21345. [Google Scholar] [CrossRef] [PubMed]
- Cizek, L.; Schlaghamerský, J.; Bořucký, J.; Hauck, D.; Helešic, J. Range expansion of an endangered beetle: Alpine longhorn Rosalia alpina (Coleoptera: Cerambycidae) spreads to the lowlands of Central Europe. Entomol. Fenn. 2009, 20, 200–206. [Google Scholar] [CrossRef]
- Cardé, R.T. Defining attraction and aggregation pheromones: Teleological versus functional perspective. J. Chem. Ecol. 2014, 40, 519–520. [Google Scholar] [CrossRef]
- Yasui, H.; Wakamura, S.; Arakaki, N.; Yasuda, T.; Akino, T.; Fukaya, M. Collection and quantification of airborne pheromone from individual females of the black chafer Holotrichia loochooana loochooana (Coleoptera: Scarabaeidae): Heterogeneity of feral females in respect to pheromone release. Appl. Entomol. Zool. 2007, 42, 143–150. [Google Scholar] [CrossRef]
- Itoh, M.; Shimizu, M.; Hirano, K.; Satoh, T.; Miura, M. Rhodium-catalyzed decarboxylative and dehydrogenative coupling of maleic acids with alkynes and alkenes. J. Org. Chem. 2013, 78, 11427–11432. [Google Scholar] [CrossRef]
- Kovát, E. Gas chromatographic characterization of organic substances in the retention index system. Adv. Chromatogr. 1965, 1, 229–247. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 22 December 2022).
- Allison, J.D.; Borden, J.H.; Seybold, S.J. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Fettköther, R.; Dettner, K.; Schröder, F.; Meyer, H.; Francke, W.; Noldt, U. The male pheromone of the old house borer Hylotrupes bajulus (L.) (Coleoptera: Cerambycidae): Identification and female response. Experientia 1995, 51, 270–277. [Google Scholar] [CrossRef]
- Hanks, L.M.; Millar, J.G. Sex and aggregation-sex pheromones of cerambycid beetles: Basic science and practical applications. J. Chem. Ecol. 2016, 42, 631–654. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.; Xu, T.; Wickham, J.; Chen, Y.; Hao, D.; Hanks, L.M.; Millar, J.G.; Teale, S.A. Identification of a male-produced pheromone component of the citrus longhorned beetle, Anoplophora chinensis. PLoS ONE 2015, 10, e0134358. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Takahashi, J.; Nakagawa, Y.; Sakai, T. Behavioral responses of female grape borer to synthetic male sex pheromone components. Appl. Entomol. Zool. 1986, 21, 21–27. [Google Scholar] [CrossRef]
- Lemay, M.A.; Silk, P.J.; Sweeney, J. Calling behavior of Tetropium fuscum (Coleoptera: Cerambycidae: Spondylidinae). Can. Entomol. 2010, 142, 256–260. [Google Scholar] [CrossRef]
- Noldt, U.; Fettköther, R.; Dettner, K. Structure of the sex pheromone-producing prothoracic glands of the male old house borer, Hylotrupes bajulus (L.) (Coleoptera: Cerambycidae). Int. J. Insect Morphol. Embryol. 1995, 24, 223–234. [Google Scholar] [CrossRef]
- Sakai, T.; Nakagawa, Y.; Takahashi, J.; Iwabuchi, K.; Ishii, K. Isolation and identification of the male sex pheromone of the grape borer Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae). Chem. Lett. 1984, 13, 263–264. [Google Scholar] [CrossRef]
- Silk, P.J.; Sweeney, J.; Wu, J.; Price, J.; Gutowski, J.M.; Kettela, E.G. Evidence for a male-produced pheromone in Tetropium fuscum (F.) and Tetropium cinnamopterum (Kirby) (Coleoptera: Cerambycidae). Naturwissenschaften 2007, 94, 697–701. [Google Scholar] [CrossRef]
- Teale, S.A.; Wickham, J.D.; Zhang, F.; Su, J.; Chen, Y.; Xiao, W.; Hanks, L.M.; Millar, J.G. Male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. J. Econ. Entomol. 2011, 104, 1592–1598. [Google Scholar] [CrossRef]
- Cohen, C.; Liltved, W.R.; Colville, J.F.; Shuttleworth, A.; Weissflog, J.; Svatoš, A.; Bytebier, B.; Johnson, S.D. Sexual deception of a beetle pollinator through floral mimicry. Curr. Biol. 2021, 31, 1962–1969. [Google Scholar] [CrossRef] [PubMed]
- Ginzel, M.D.; Hanks, L.M. Role of host plant volatiles in mate location for three species of longhorned beetles. J. Chem. Ecol. 2005, 31, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef] [PubMed]
- Millar, J.G.; Hanks, L.M. Chemical ecology of Cerambycids. In Cerambycidae of the World: Biology and Pest Management; Wang, Q., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 161–208. [Google Scholar]
- Miller, D.R. Ethanol and (−)-α-pinene: Attractant kairomones for some large wood-boring beetles in southeastern USA. J. Chem. Ecol. 2006, 32, 779–794. [Google Scholar] [CrossRef]
- Iwabuchi, K. Mating behavior of Xylotrechus pyrrhoderus Bates. III. Pheromone secretion by male. App. Entomol. Zool. 1986, 21, 606–612. [Google Scholar] [CrossRef]
- Iwabuchi, K.; Takahashi, J.; Sakai, T. Occurrence of 2,3-octanediol and 2-hydroxy-3-octanone, possible male sex pheromone in Xylotrechus chinensis. Appl. Entomol. Zool. 1987, 22, 110–111. [Google Scholar] [CrossRef]
- Hall, D.R.; Cork, A.; Phythian, S.J.; Chittamuru, S.B.; Jayarama, K.; Venkatesha, M.G.; Sreedharan, K.; Vinod Kumar, P.K.; Seetharama, H.G.; Naidu, R. Identification of components of male-produced pheromone of coffee white stem borer, Xylotrechus quadripes. J. Chem. Ecol. 2006, 32, 195–219. [Google Scholar] [CrossRef]
- Pajares, J.A.; Álvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef]
- Zhang, A.; Oliver, J.E.; Aldrich, J.R.; Wang, B.; Mastro, V.C. Stimulatory beetle volatiles for the Asian longhorned beetle, Anoplophora glabripennis (Motschulsky). Z. Naturforsch. 2002, 57, 553–558. [Google Scholar] [CrossRef]
- Hanks, L.M.; Wang, Q. Reproductive biology of Cerambycids. In Cerambycidae of the World: Biology and Pest Management; Wang, Q., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 133–160. [Google Scholar]
- Larsson, M.C. Pheromones and other semiochemicals for monitoring rare and endangered species. J. Chem. Ecol. 2016, 42, 853–868. [Google Scholar] [CrossRef]
- Musa, N.; Andersson, K.; Burman, J.; Andersson, F.; Hedenström, E.; Jansson, N.; Paltto, H.; Westerberg, L.; Winde, I.; Larsson, M.C.; et al. Using sex pheromone and a multi-scale approach to predict the distribution of a rare saproxylic beetle. PLoS ONE 2013, 8, e66149. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.; Bergman, K.O.; Andersson, F.; Hedenström, E.; Jansson, N.; Burman, J.; Winde, I.; Larsson, M.C.; Milberg, P. High-accuracy sampling of saproxylic diversity indicators at regional scales with pheromones: The case of Elater ferrugineus (Coleoptera, Elateridae). Biol. Conserv. 2014, 171, 156–166. [Google Scholar] [CrossRef]
- Bosso, L.; Rebelob, H.; Garonnac, A.P.; Russo, D. Modelling geographic distribution and detecting conservation gaps in Italy for the threatened beetle Rosalia alpina. J. Nat. Conserv. 2013, 21, 72–80. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukaya, M.; Kiriyama, S.; Yagami, S.; Iwata, R.; Yasui, H.; Tokoro, M.; Zou, Y.; Millar, J.G. Identification of a Male-Produced Aggregation Sex Pheromone in Rosalia batesi, an Endemic Japanese Longhorn Beetle. Insects 2023, 14, 867. https://doi.org/10.3390/insects14110867
Fukaya M, Kiriyama S, Yagami S, Iwata R, Yasui H, Tokoro M, Zou Y, Millar JG. Identification of a Male-Produced Aggregation Sex Pheromone in Rosalia batesi, an Endemic Japanese Longhorn Beetle. Insects. 2023; 14(11):867. https://doi.org/10.3390/insects14110867
Chicago/Turabian StyleFukaya, Midori, Satoshi Kiriyama, Saki Yagami, Ryûtarô Iwata, Hiroe Yasui, Masahiko Tokoro, Yunfan Zou, and Jocelyn G. Millar. 2023. "Identification of a Male-Produced Aggregation Sex Pheromone in Rosalia batesi, an Endemic Japanese Longhorn Beetle" Insects 14, no. 11: 867. https://doi.org/10.3390/insects14110867