

Functional Divergence of the Tribolium castaneum engrailed and invected Paralogs
 , ,
, , 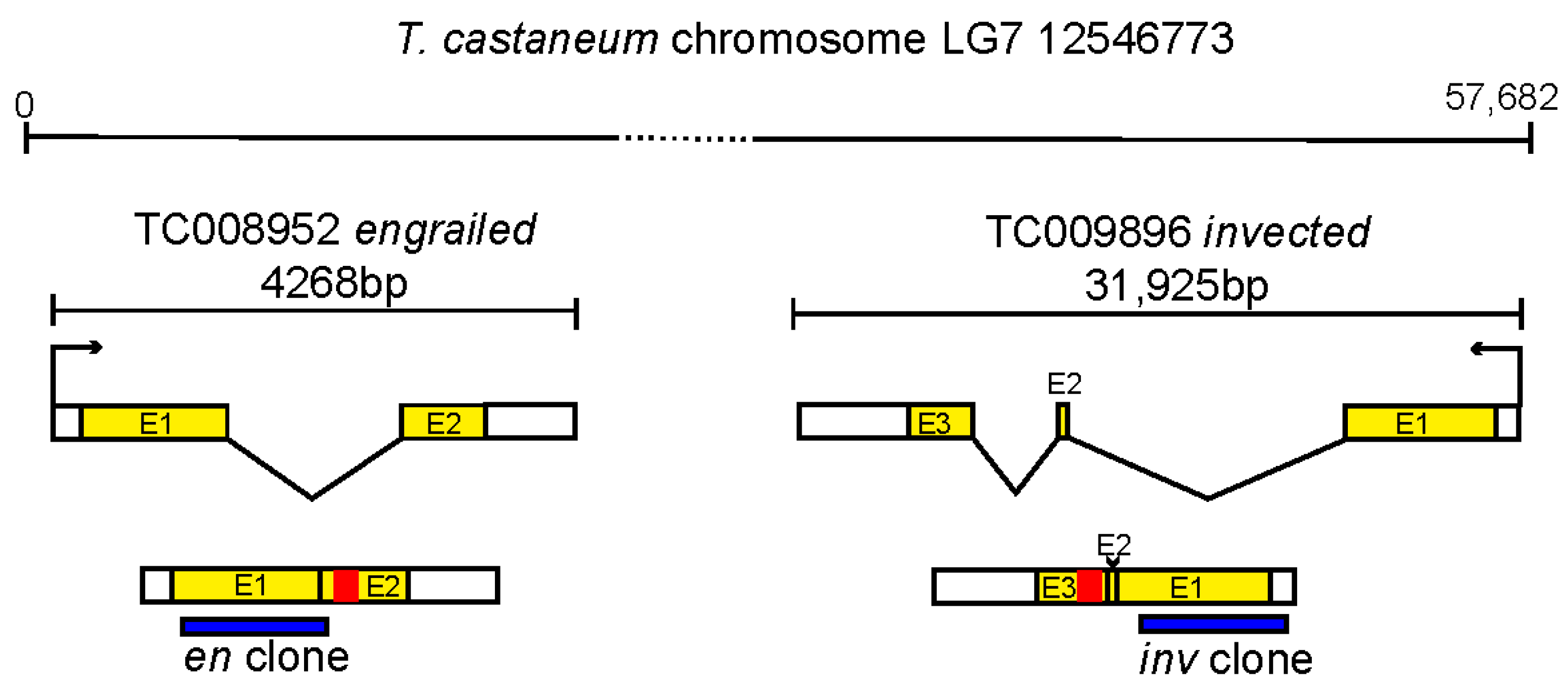{kind=link}
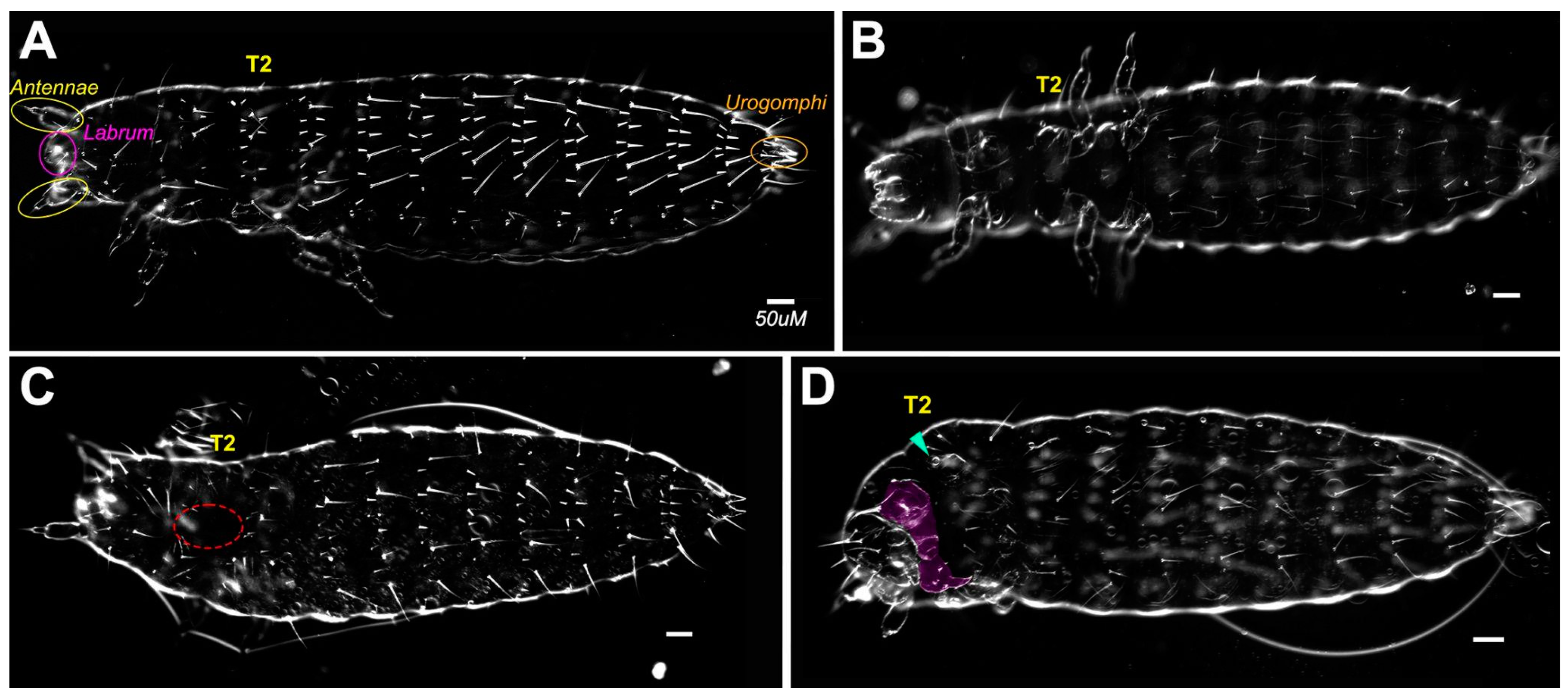{kind=link}
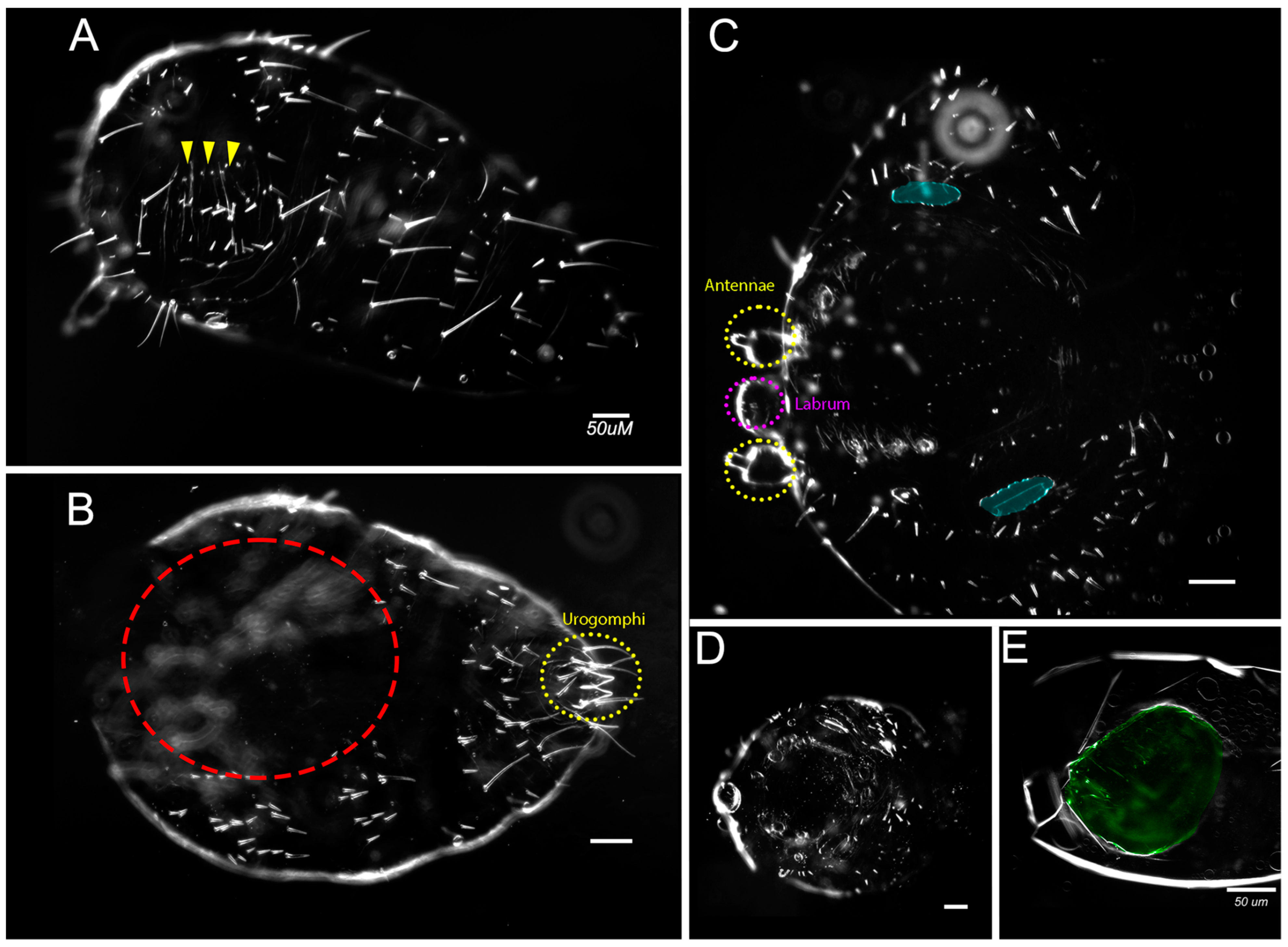{kind=link}
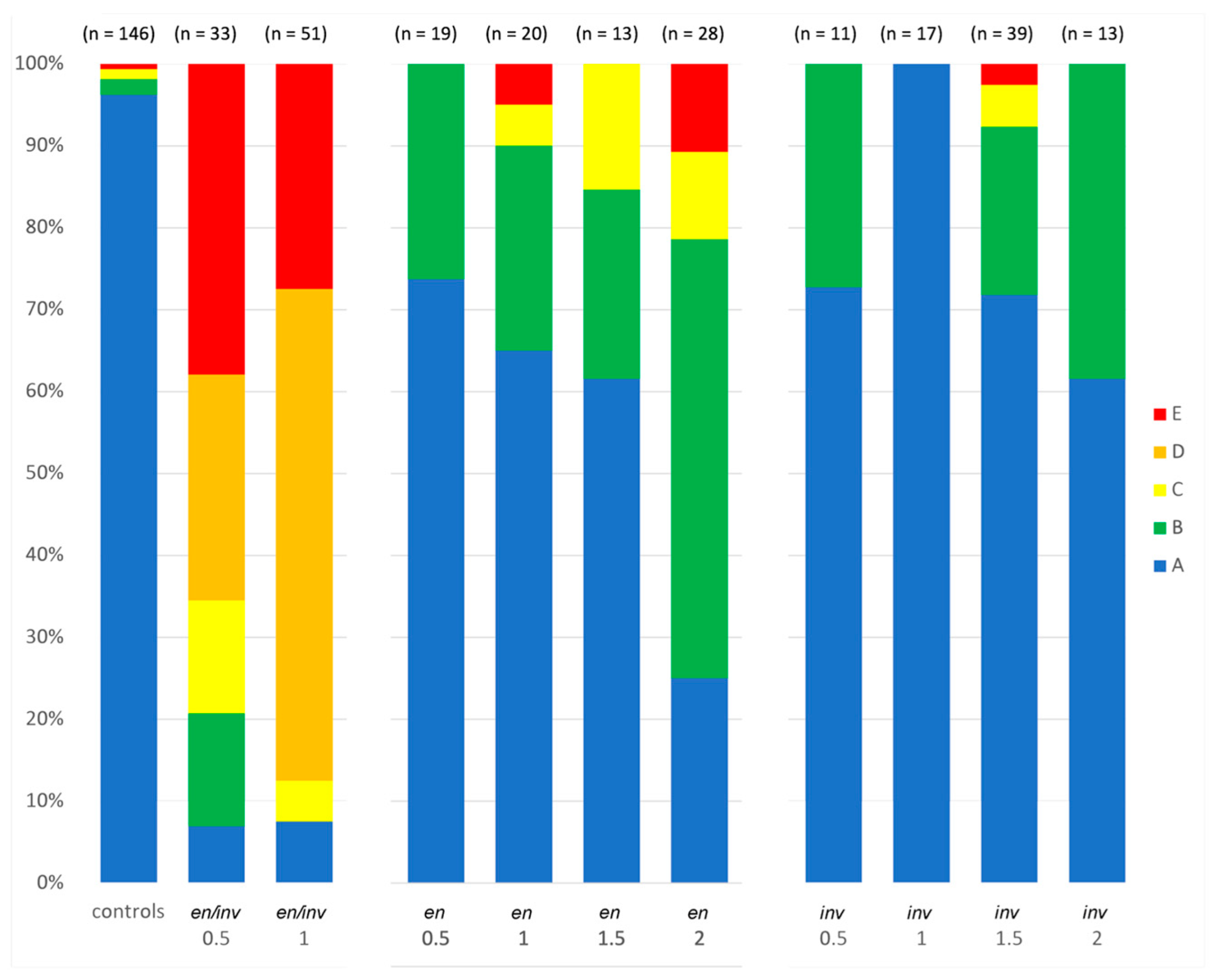{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Husbandry, Egg Collections, and Injection Preparations
2.2. Cloning and dsRNA Synthesis
2.3. Microinjections and Needle Preparation
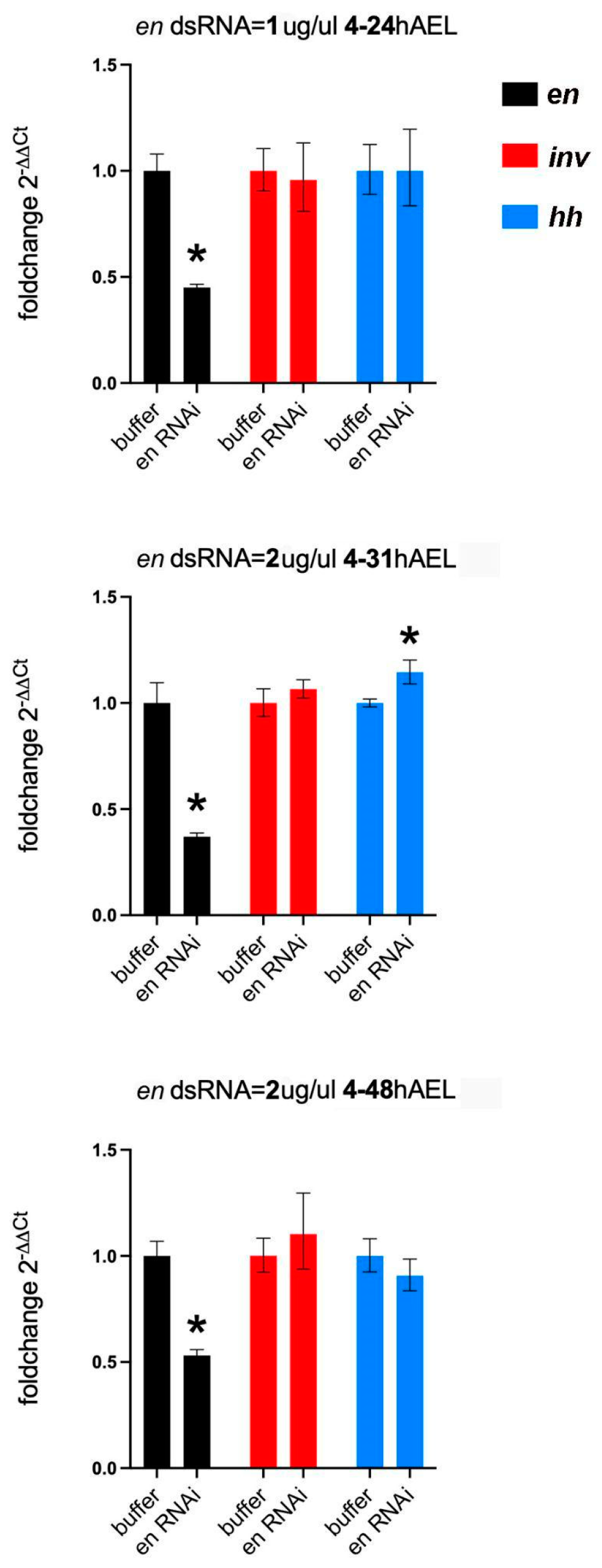
2.4. qPCR Analysis
2.5. Cuticle Preparation
2.6. Statistical Analysis
2.7. Embryo Fixation and Immunohistochemistry
3. Results
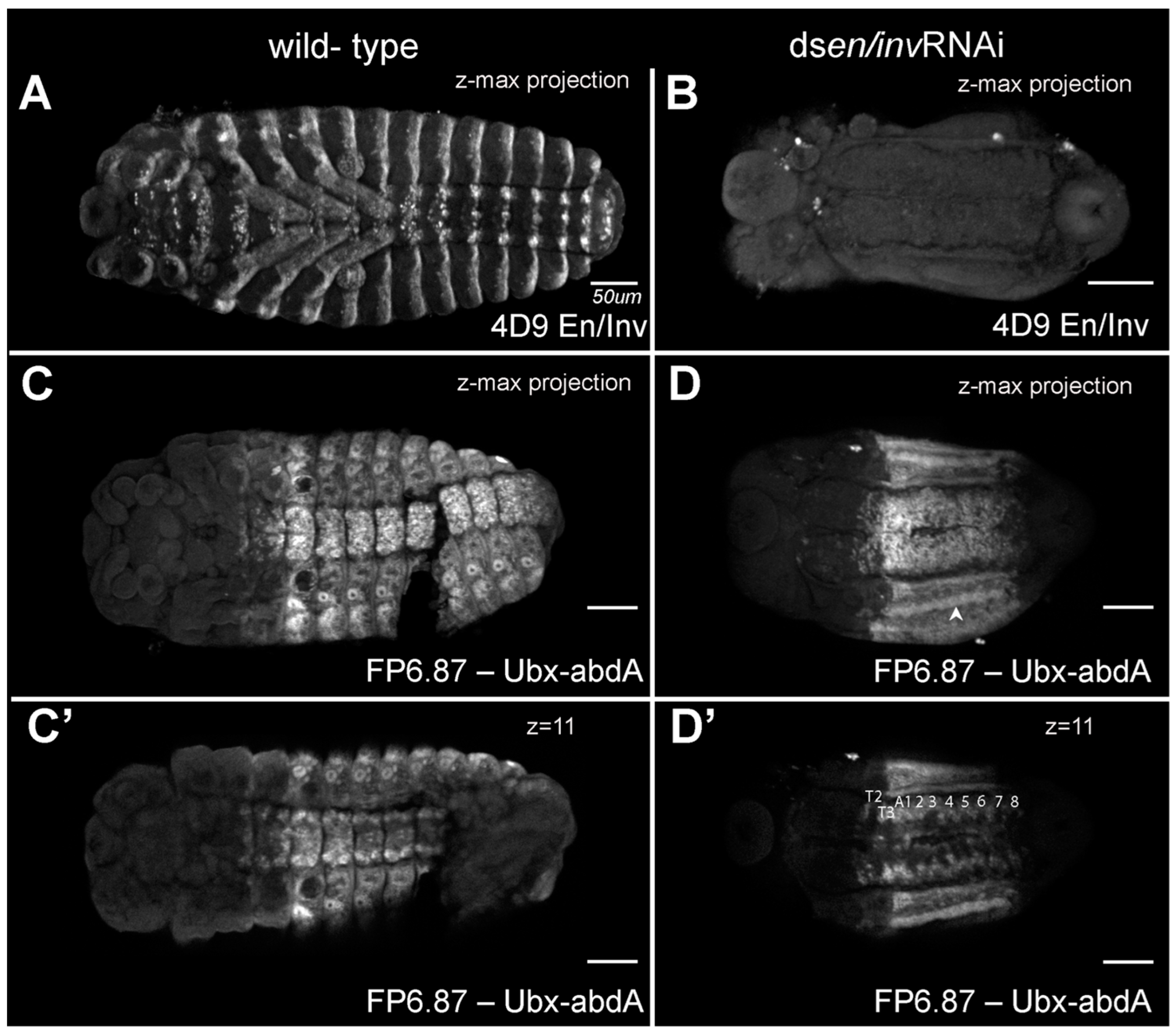
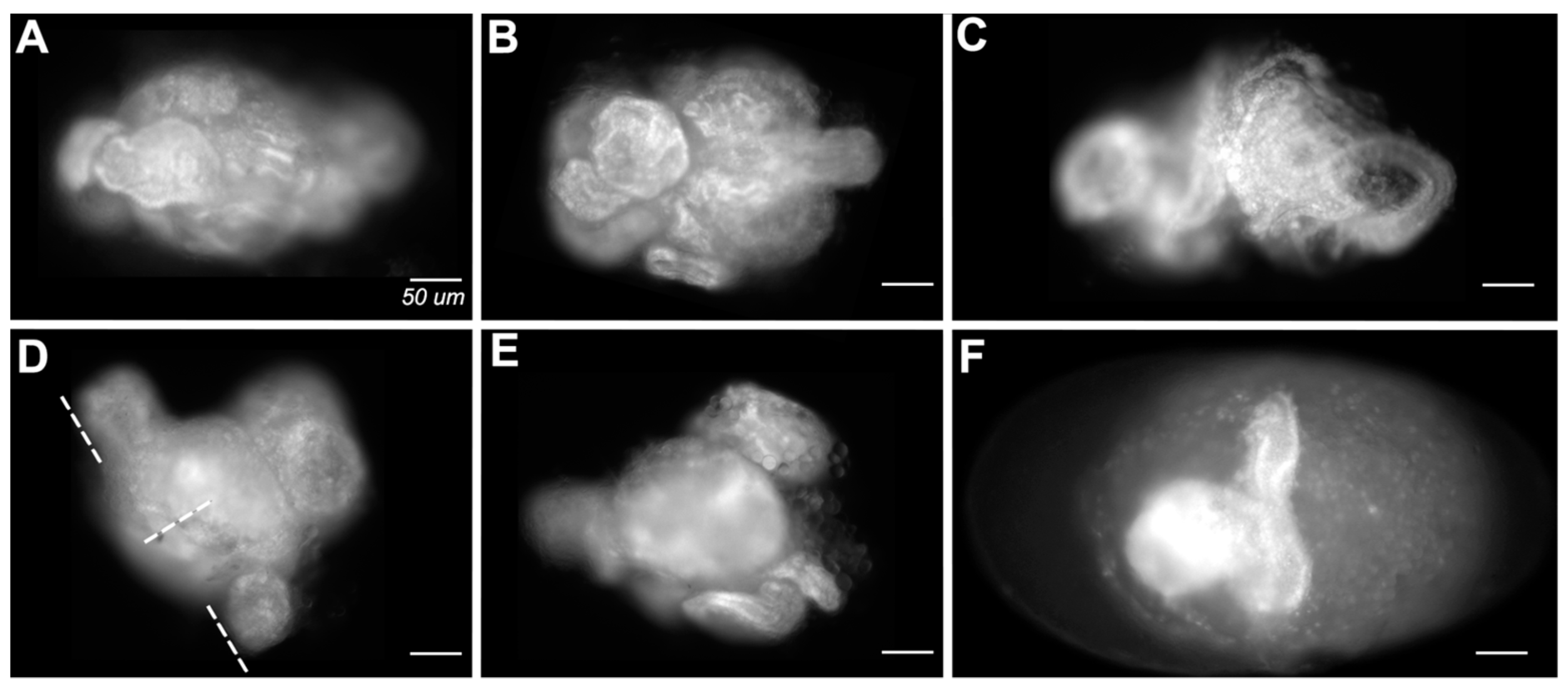
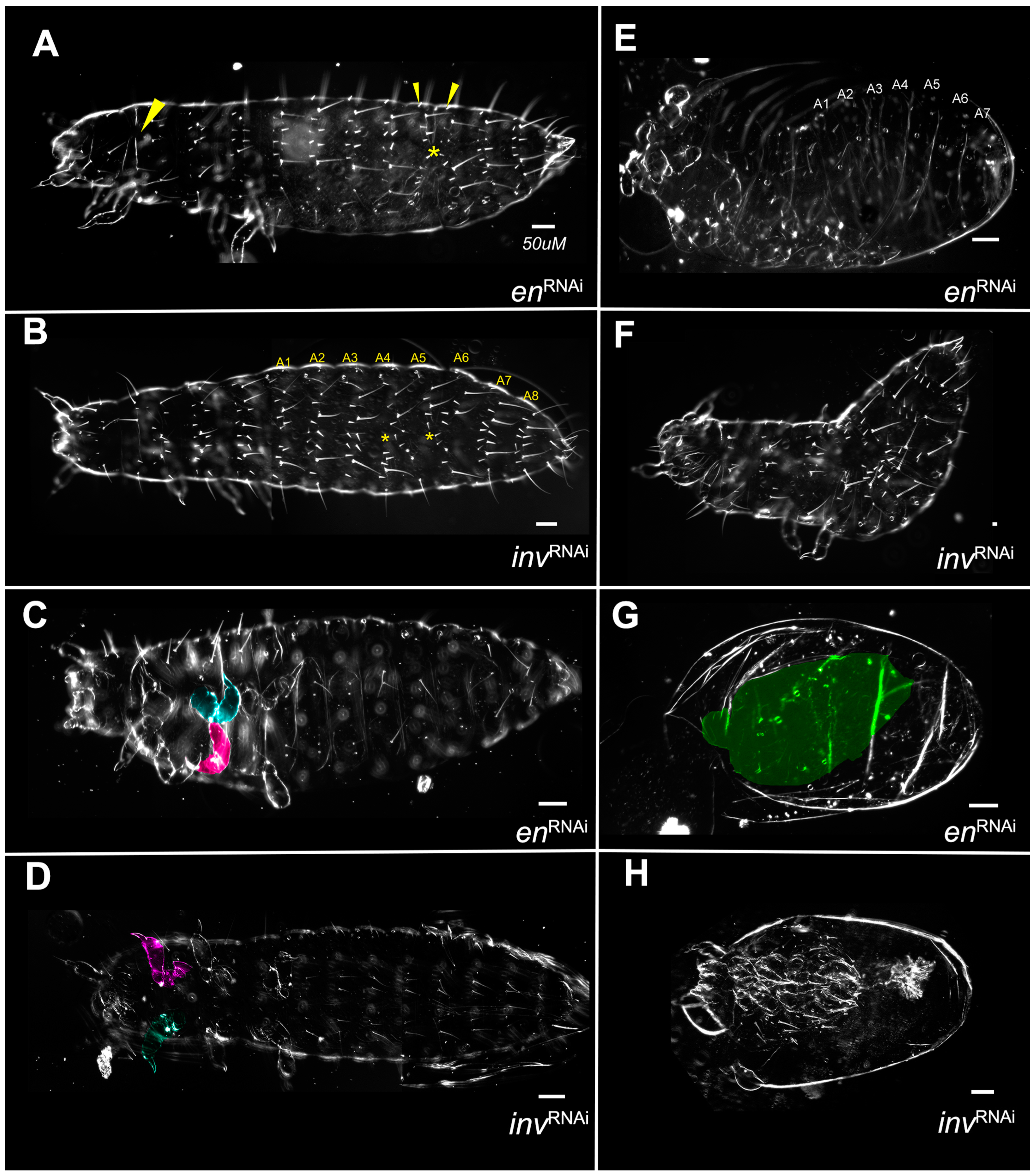
3.1. Knockdown of Both en and inv Produces a Range of Morphological Defects
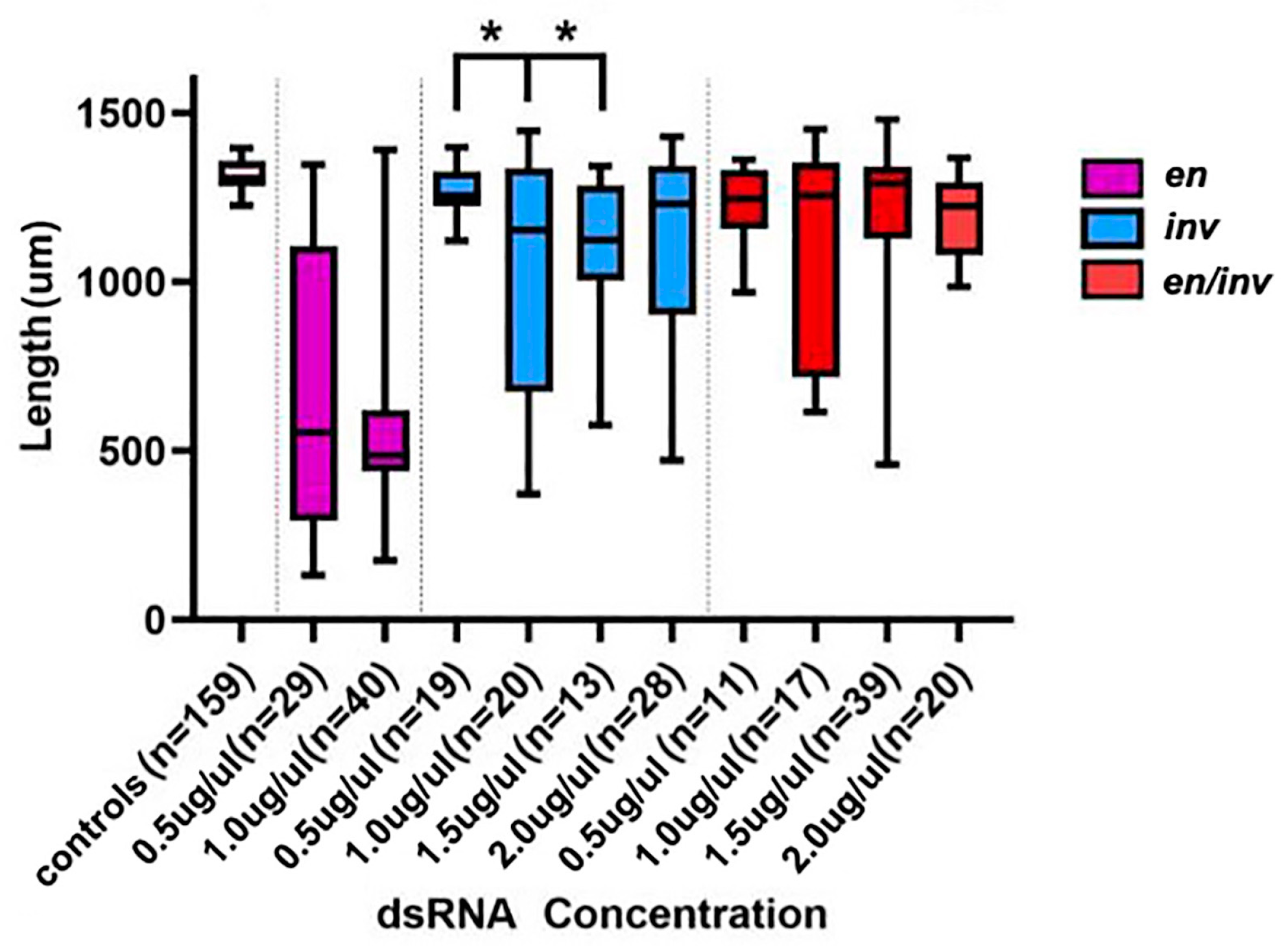
3.1.1. Segment Number
3.1.2. Larval Length
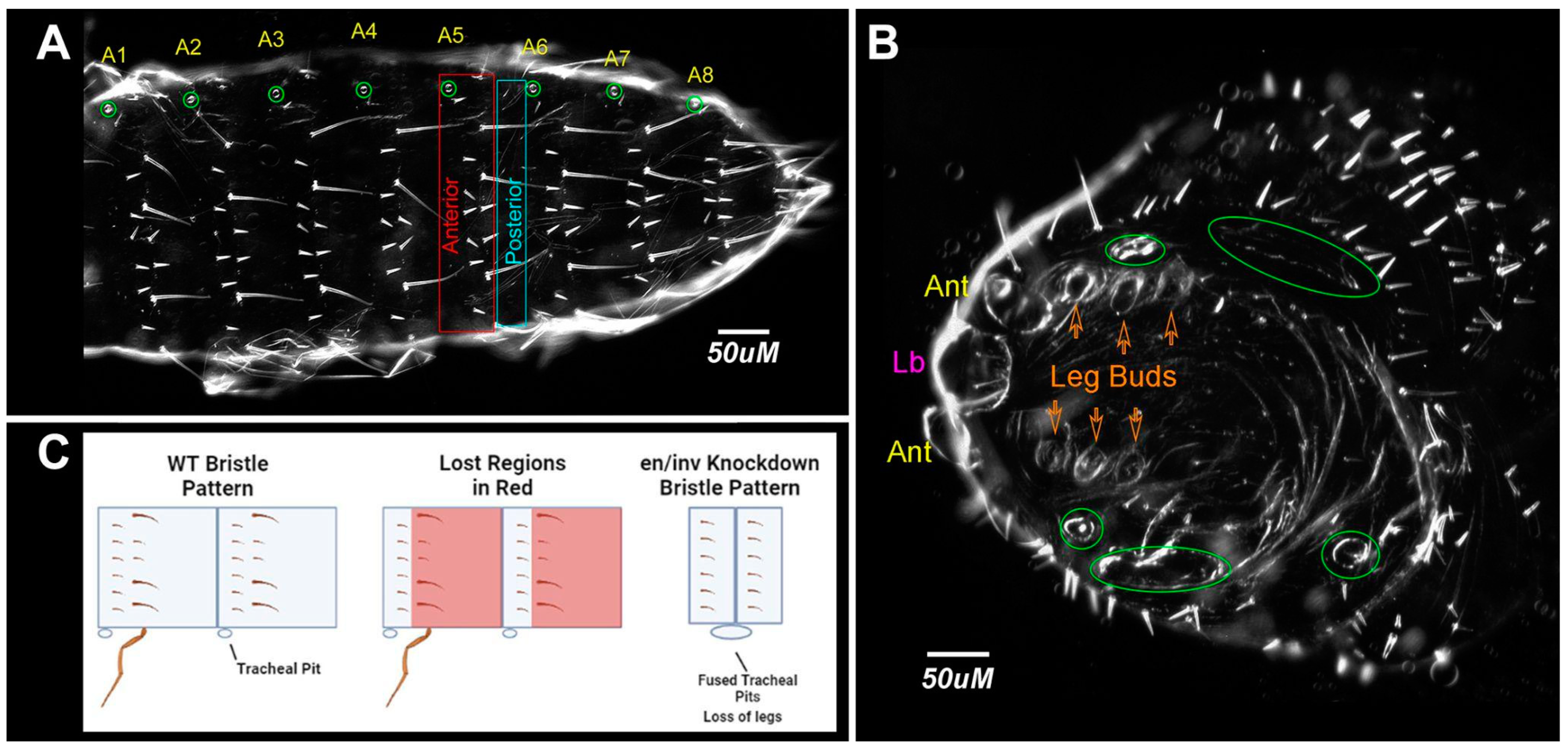
3.1.3. Appendage Formation
3.1.4. Bristle Patterns
3.1.5. Dorsal Closure
3.1.6. Failure to Form Cuticle
3.2. Single Knockdowns
3.2.1. Asegmental embryos at High dsRNA Tc-en or Tc-inv Concentrations
3.2.2. En Single Knockdowns Do Not Significantly Affect inv mRNA Levels
4. Discussion
4.1. Paralog Redundancy—A Model for the Tribolium en/inv Paralogs Acting Synergistically
4.2. En/Inv Are Redundant for Gnathal and Thoracic Appendage Formation
4.3. En/Hh-Wg—Regulatory Loop
4.4. Loss of Intersegmental Cellular Identity vs. Segment Polarity
4.5. Is the Segment Addition Process Disrupted?
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peel, A.D.; Chipman, A.D.; Akam, M. Arthropod segmentation: Beyond the Drosophila paradigm. Nat. Rev. Genet. 2005, 6, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Auman, T.; Chipman, A.D. The Evolution of Gene Regulatory Networks that Define Arthropod Body Plans. Integr. Comp. Biol. 2017, 57, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morata, G.; Lawrence, P.A. Control of compartment development by the engrailed gene in Drosophila. Nature 1975, 255, 614–617. [Google Scholar] [CrossRef]
- Kornberg, T. Engrailed: A gene controlling compartment and segment formation in Drosophila. Proc. Natl. Acad. Sci. USA 1981, 78, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, P.A.; Struhl, G. Further studies of the engrailed phenotype in Drosophila. EMBO J. 1982, 1, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.H.; Martin-Blanco, E.; Coleman, K.G.; Poole, S.J.; Ellis, M.C.; Kornberg, T.B.; Goodman, C.S. Expression of engrailed proteins in arthropods, annelids, and chordates. Cell 1989, 58, 955–968. [Google Scholar] [CrossRef]
- Condron, B.G.; Patel, N.H.; Zinn, K. engrailed controls glial/neuronal cell fate decisions at the midline of the central nervous system. Neuron 1994, 13, 541–554. [Google Scholar] [CrossRef]
- Whitington, P.M.; Meier, T.; King, P. Segmentation, neurogenesis and formation of early axonal pathways in the centipede, Ethmostigmus rubripes (Brandt). Roux’s Arch. Dev. Biol. 1991, 199, 349–363. [Google Scholar] [CrossRef]
- Keys, D.N.; Lewis, D.L.; Selegue, J.E.; Pearson, B.J.; Goodrich, L.V.; Johnson, R.L.; Gates, J.; Scott, M.P.; Carroll, S.B. Recruitment of a hedgehog Regulatory Circuit in Butterfly Eyespot Evolution. Science 1999, 283, 532–534. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, C.R.; E Selegue, J.; Monteiro, A.; French, V.; Brakefield, P.M.; Carroll, S.B. The generation and diversification of butterfly eyespot color patterns. Curr. Biol. 2001, 11, 1578–1585. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.; Podlaha, O. Wings, horns, and butterfly eyespots: How do complex traits evolve? PLoS Biol. 2009, 7, e37. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, T.D.; Ramos, D.; Monteiro, A. Expression of Multiple engrailed Family Genes in Eyespots of Bicyclus anynana Butterflies Does Not Implicate the Duplication Events in the Evolution of This Morphological Novelty. Front. Ecol. Evol. 2020, 8, 227. [Google Scholar] [CrossRef]
- Patel, N.H.; Kornberg, T.B.; Goodman, C.S. Expression of engrailed during segmentation in grasshopper and crayfish. Development 1989, 107, 201–212. [Google Scholar] [CrossRef]
- Angelini, D.R.; Kaufman, T.C. Functional analyses in the milkweed bug Oncopeltus fasciatus (Hemiptera) support a role for Wnt signaling in body segmentation but not appendage development. Dev. Biol. 2005, 283, 409–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegler, M.V.; Jia, X.X. engrailed negatively regulates the expression of cell adhesion molecules connectin and neuroglian in embryonic Drosophila nervous system. Neuron 1999, 22, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marie, B.; Bacon, J.P. Two engrailed -related genes in the cockroach: Cloning, phylogenetic analysis, expression and isolation of splice variants. Dev. Genes Evol. 2000, 210, 436–448. [Google Scholar] [CrossRef]
- Williams, T.; Blachuta, B.; Hegna, T.A.; Nagy, L.M. Decoupling elongation and segmentation: Notch involvement in anostracan crustacean segmentation. Evol. Dev. 2012, 14, 372–382. [Google Scholar] [CrossRef]
- Janssen, R. Segment polarity gene expression in a myriapod reveals conserved and diverged aspects of early head patterning in arthropods. Dev. Genes Evol. 2012, 222, 299–309. [Google Scholar] [CrossRef]
- Green, J.; Akam, M. Evolution of the pair rule gene network: Insights from a centipede. Dev. Biol. 2013, 382, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Gibert, J.M.; Mouchel-Vielh, E.; Quéinnec, E.; Deutsch, J.S. Barnacle duplicate engrailed genes: Divergent expression patterns and evidence for a vestigial abdomen. Evol. Dev. 2000, 2, 194–202. [Google Scholar] [CrossRef]
- Nakagaki, Y.; Sakuma, M.; Machida, R. Expression of engrailed -family genes in the jumping bristletail and discussion on the primitive pattern of insect segmentation. Dev. Genes Evol. 2015, 225, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, T.; Sidén, I.; O’Farrell, P.; Simon, M. The engrailed locus of Drosophila: In situ localization of transcripts reveals compartment-specific expression. Cell 1985, 40, 45–53. [Google Scholar] [CrossRef]
- Peterson, M.D.; Popadic, A.; Kaufman, T.C. The expression of two engrailed-related genes in an apterygote insect and a phylogenetic analysis of insect engrailed -related genes. Dev. Genes Evol. 1998, 208, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Peel, A.D.; Telford, M.J.; Akam, M. The evolution of hexapod engrailed-family genes: Evidence for conservation and concerted evolution. Proc. Biol. Sci. 2006, 273, 1733–1742. [Google Scholar] [CrossRef]
- Perrimon, N.; Mahowald, A.P. Multiple functions of segment polarity genes in Drosophila. Dev. Biol. 1987, 119, 587–600. [Google Scholar] [CrossRef]
- Baker, N.E. Embryonic and imaginal requirements for wingless, a segment-polarity gene in Drosophila. Dev. Biol. 1988, 125, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Bejsovec, A.; Martinez Arias, A. Roles of wingless in patterning the larval epidermis of Drosophila. Development 1991, 113, 471–485. [Google Scholar] [CrossRef]
- Bejsovec, A.; Wieschaus, E. Segment polarity gene interactions modulate epidermal patterning in Drosophila embryos. Development 1993, 119, 501–517. [Google Scholar] [CrossRef]
- Heemskerk, J.; DiNardo, S.; Kostriken, R.; O’Farrell, P.H. Multiple modes of engrailed regulation in the progression towards cell fate determination. Nature 1991, 352, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Campbell, G.L.; Caveney, S. engrailed gene expression in the abdominal segment of Oncopeltus: Gradients and cell states in the insect segment. Development 1989, 106, 727–737. [Google Scholar] [CrossRef]
- Oppenheimer, D.I.; MacNicol, A.M.; Patel, N.H. Functional conservation of the wingless-engrailed interaction as shown by a widely applicable baculovirus misexpression system. Curr. Biol. 1999, 9, 1288–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Choe, C.P. Functional analysis of engrailed in Tribolium segmentation. Mech. Dev. 2020, 161, 103594. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Gustavson, E.; Goldsborough, A.S.; Ali, Z.; Kornberg, T.B. The Drosophila engrailed and invected genes: Partners in regulation, expression and function. Genetics 1996, 142, 893–906. [Google Scholar] [CrossRef]
- Cheng, Y.; Brunner, A.L.; Kremer, S.; DeVido, S.K.; Stefaniuk, C.M.; Kassis, J.A. Co-regulation of invected and engrailed by a complex array of regulatory sequences in Drosophila. Dev. Biol. 2014, 395, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.-L.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Ahzhanov, A.; Kaufman, T.C. Evolution of distinct expression patterns for engrailed paralogues in higher crustaceans (Malacostraca). Dev. Genes Evol. 2000, 210, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Marie, B.; Blagburn, J.M. Differential roles of engrailed paralogs in determining sensory axon guidance and synaptic target recognition. J. Neurosci. 2003, 23, 7854–7862. [Google Scholar] [CrossRef] [Green Version]
- Henschel, A.; Buchholz, F.; Habermann, B. DEQOR: A web-based tool for the design and quality control of siRNAs. Nucleic Acids Res. 2004, 32 (Suppl. 2), W113–W120. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.C.; Miyata, K.; Brown, S.J.; Tomoyasu, Y. Dissecting systemic RNA interference in the red flour beetle Tribolium castaneum: Parameters affecting the efficiency of RNAi. PLoS ONE 2012, 7, e47431. [Google Scholar] [CrossRef] [Green Version]
- Kelsh, R.; Weinzierl, R.O.J.; White, R.A.H.; Akam, M. Homeotic gene expression in the locust Schistocerca: An antibody that detects conserved epitopes in ultrabithorax and abdominal-A proteins. Dev. Genet. 1994, 15, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Tabata, T.; Eaton, S.; Kornberg, T.B. The Drosophila hedgehog gene is expressed specifically in posterior compartment cells and is a target of engrailed regulation. Genes Dev. 1992, 6, 2635–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nodine, M.D.; Yadegari, R.; Tax, F.E. RPK1 and TOAD2 are two receptor-like kinases redundantly required for arabidopsis embryonic pattern formation. Dev. Cell 2007, 12, 943–956. [Google Scholar] [CrossRef] [Green Version]
- Lambie, E.J.; Kimble, J. Two homologous regulatory genes, lin-12 and glp-1, have overlapping functions. Development 1991, 112, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Vanden Broek, K.; Han, X.; Hansen, D. Redundant mechanisms regulating the proliferation vs. differentiation balance in the C. elegans germline. Front. Cell Dev. Biol. 2022, 10, 960999. [Google Scholar] [CrossRef]
- Simcox, A.A.; Roberts, I.J.; Hersperger, E.; Gribbin, M.C.; Shearn, A.; Whittle, J.R. Imaginal discs can be recovered from cultured embryos mutant for the segment-polarity genes engrailed, naked and patched but not from wingless. Development 1989, 107, 715–722. [Google Scholar] [CrossRef]
- Cohen, B.; Simcox, A.A.; Cohen, S.M. Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development 1993, 117, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M. Specification of limb development in the Drosophila embryo by positional cues from segmentation genes. Nature 1990, 343, 173–177. [Google Scholar] [CrossRef]
- Ober, K.A.; Jockusch, E.L. The roles of wingless and decapentaplegic in axis and appendage development in the red flour beetle, Tribolium castaneum. Dev. Biol. 2006, 294, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Posnien, N.; Bashasab, F.; Bucher, G. The insect upper lip (labrum) is a nonsegmental appendage-like structure. Evol. Dev. 2009, 11, 480–488. [Google Scholar] [CrossRef]
- Farzana, L.; Brown, S.J. Hedgehog signaling pathway function conserved in Tribolium segmentation. Dev. Genes Evol. 2008, 218, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lev, O.; Edgecombe, G.D.; Chipman, A.D. Serial Homology and Segment Identity in the Arthropod Head. Integr. Org. Biol. 2022, 4, obac015. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, R.E. Principles of Insect Morphology, 1st ed.; McGraw-Hill Publications in the Zoological Sciences: New York, NY, USA, 1935; Volume IX, 667p. [Google Scholar]
- Struhl, G. A homoeotic mutation transforming leg to antenna in Drosophila. Nature 1981, 292, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Rogers, B.T.; Kaufman, T.C. Structure of the insect head in ontogeny and phylogeny: A view from Drosophila. Int. Rev. Cytol. 1997, 174, 1–84. [Google Scholar]
- Scholtz, G.; Edgecombe, G.D. The evolution of arthropod heads: Reconciling morphological, developmental and palaeontological evidence. Dev. Genes Evol. 2006, 216, 395–415. [Google Scholar] [CrossRef]
- Ortega-Hernández, J.; Janssen, R.; Budd, G.E. Origin and evolution of the panarthropod head—A palaeobiological and developmental perspective. Arthropod Struct. Dev. 2017, 46, 354–379. [Google Scholar] [CrossRef]
- Haas, M.S.; Brown, S.J.; Beeman, R.W. Pondering the procephalon: The segmental origin of the labrum. Dev. Genes Evol. 2001, 211, 89–95. [Google Scholar] [CrossRef]
- Popadić, A.; Panganiban, G.; Rusch, D.; Shear, W.A.; Kaufman, T.C. Molecular evidence for the gnathobasic derivation of arthropod mandibles and for the appendicular origin of the labrum and other structures. Dev. Genes Evol. 1998, 208, 142–150. [Google Scholar] [CrossRef]
- Prpic, N.-M.; Wigand, B.; Damen, W.G.; Klingler, M. Expression of dachshund in wild-type and Distal-less mutant Tribolium corroborates serial homologies in insect appendages. Dev. Genes Evol. 2001, 211, 467–477. [Google Scholar] [CrossRef]
- Janssen, R. The embryonic expression pattern of a second, hitherto unrecognized, paralog of the pair-rule gene sloppy-paired in the beetle Tribolium castaneum. Dev. Genes Evol. 2020, 230, 247–256. [Google Scholar] [CrossRef]
- Karr, T.L.; Ali, Z.; Drees, B.; Kornberg, T. The engrailed locus of D. melanogaster provides an essential zygotic function in precellular embryos. Cell 1985, 43 Pt 2, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A. Three distinct roles for the engrailed gene in Drosophila wing development. Curr. Biol. 1994, 4, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Tabata, T.; Schwartz, C.; Gustavson, E.; Ali, Z.; Kornberg, T.B. Creating a Drosophila wing de novo, the role of engrailed, and the compartment border hypothesis. Development 1995, 121, 3359–3369. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Struhl, G. Complementary and mutually exclusive activities of decapentaplegic and wingless organize axial patterning during Drosophila leg development. Cell 1996, 86, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Johnston, L.A.; Schubiger, G. Ectopic expression of wingless in imaginal discs interferes with decapentaplegic expression and alters cell determination. Development 1996, 122, 3519–3529. [Google Scholar] [CrossRef] [PubMed]
- Panfilio, K.A.; Jentzsch, I.M.V.; Benoit, J.B.; Erezyilmaz, D.; Suzuki, Y.; Colella, S.; Robertson, H.M.; Poelchau, M.F.; Waterhouse, R.M.; Ioannidis, P.; et al. Molecular evolutionary trends and feeding ecology diversification in the Hemiptera, anchored by the milkweed bug genome. Genome Biol. 2019, 20, 64. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; Patel, N.H.; Denell, R.E. Embryonic expression of the single Tribolium engrailed homolog. Dev. Genet. 1994, 15, 7–18. [Google Scholar] [CrossRef]
- Goldman-Huertas, B.; Sagun, J.; Nagy, L.M.; Williams, T.A. Transcriptome of the segmenting embryo of Tribolium castaneum reveals contribution of alternative pathways to developmental regulation, manuscript in preparation. 2023. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blunk, S.; Garcia-Verdugo, H.; O’Sullivan, S.; Camp, J.; Haines, M.; Coalter, T.; Williams, T.A.; Nagy, L.M. Functional Divergence of the Tribolium castaneum engrailed and invected Paralogs. Insects 2023, 14, 691. https://doi.org/10.3390/insects14080691
Blunk S, Garcia-Verdugo H, O’Sullivan S, Camp J, Haines M, Coalter T, Williams TA, Nagy LM. Functional Divergence of the Tribolium castaneum engrailed and invected Paralogs. Insects. 2023; 14(8):691. https://doi.org/10.3390/insects14080691
Chicago/Turabian StyleBlunk, Summer, Hector Garcia-Verdugo, Sierra O’Sullivan, James Camp, Michael Haines, Tara Coalter, Terri A. Williams, and Lisa M. Nagy. 2023. "Functional Divergence of the Tribolium castaneum engrailed and invected Paralogs" Insects 14, no. 8: 691. https://doi.org/10.3390/insects14080691