
Elongases of Long-Chain Fatty Acids ELO2 and ELO9 Are Involved in Cuticle Formation and Function in Fecundity in the Yellow Fever Mosquito, Aedes aegypti
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
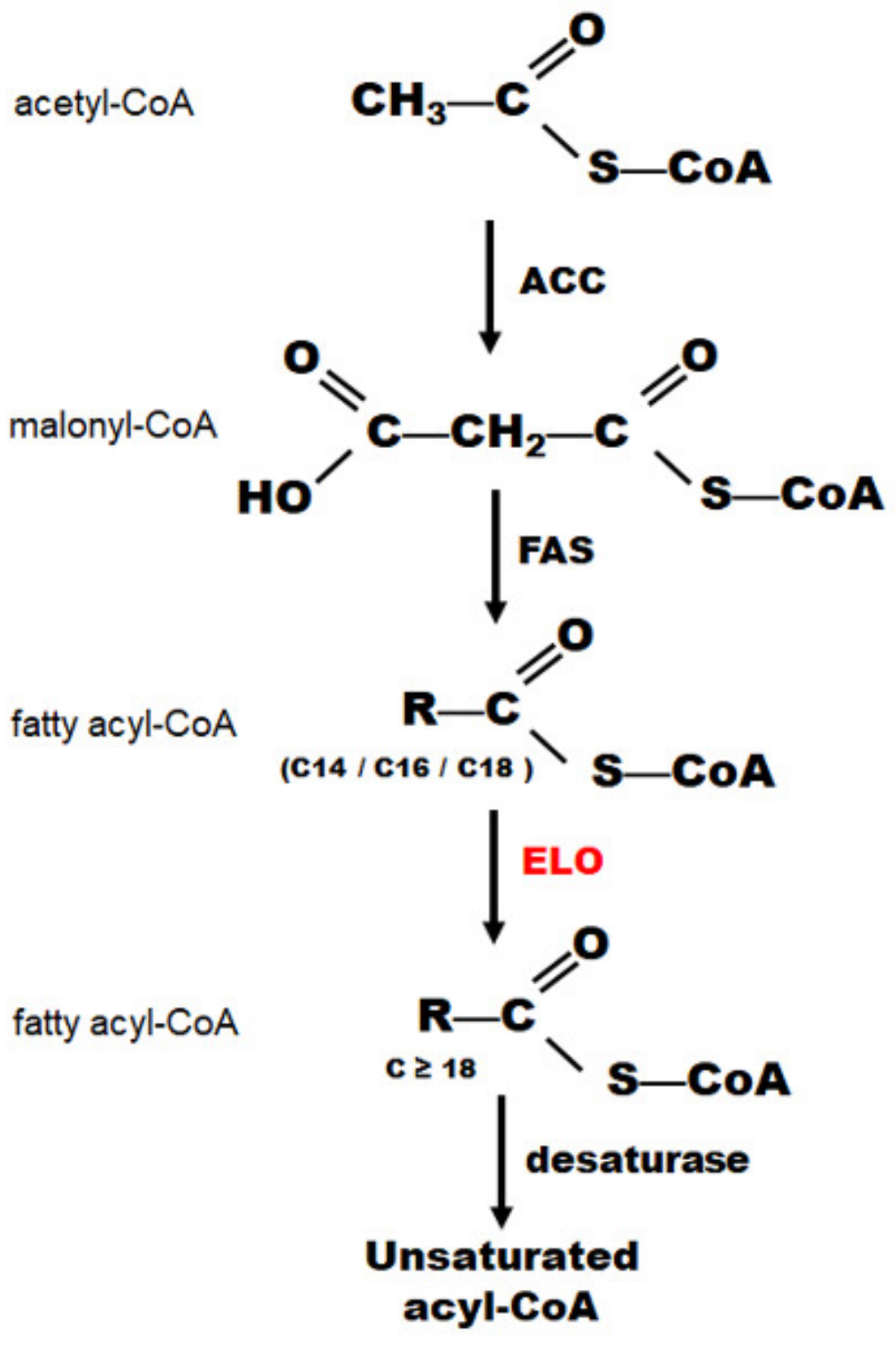
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Identification of AeELO2 and AeELO9
2.3. Quantitative Real Time PCR
2.4. RNA Interference in Larvae and Adult Mosquitoes
2.5. Cold Treatment of Mosquito Larvae and Adults
2.6. Statistical analyses
3. Results
3.1. Identification of AeELO2 and AeELO9
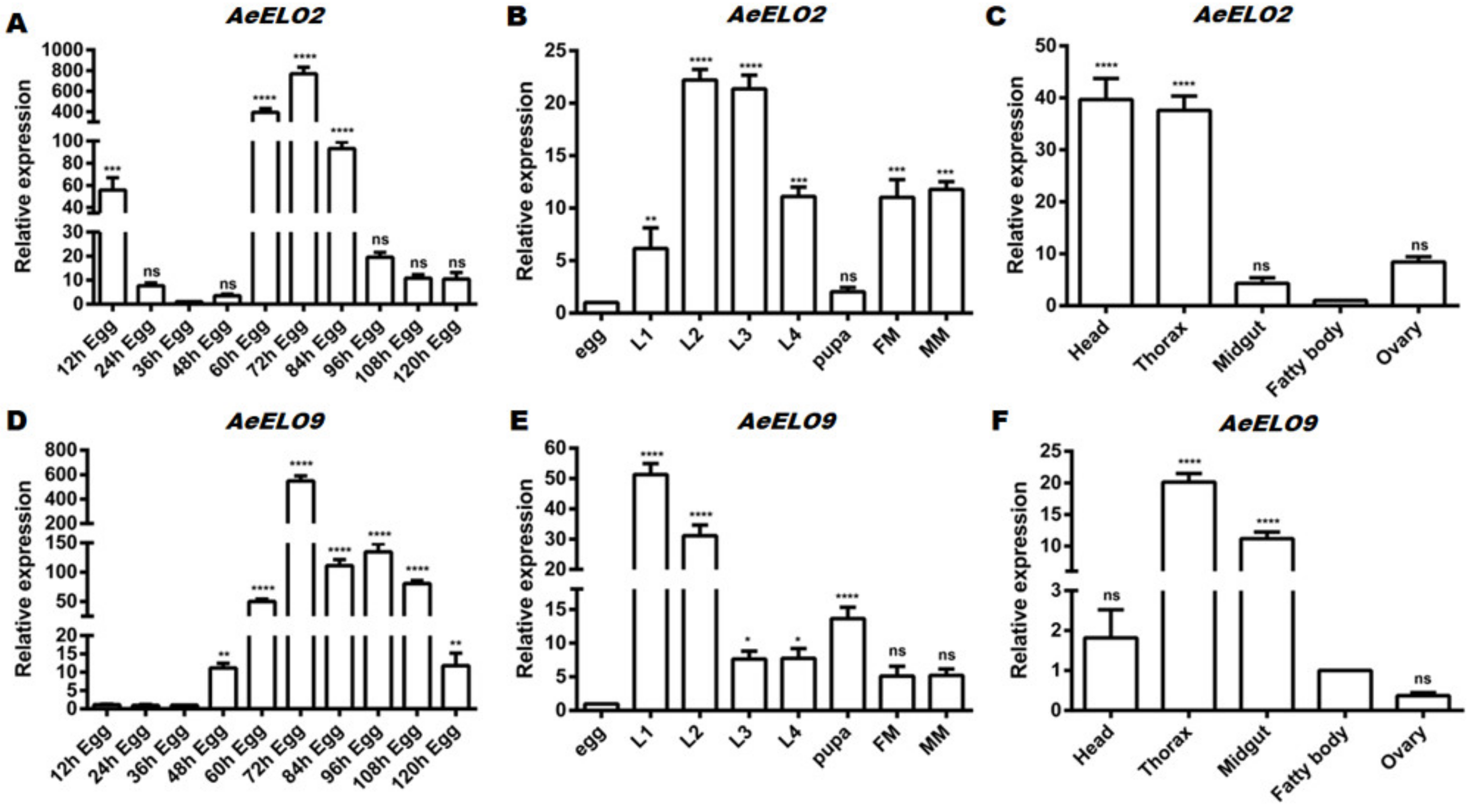
3.2. Spatiotemporal Expression Profiles of AeELO2 and AeELO9
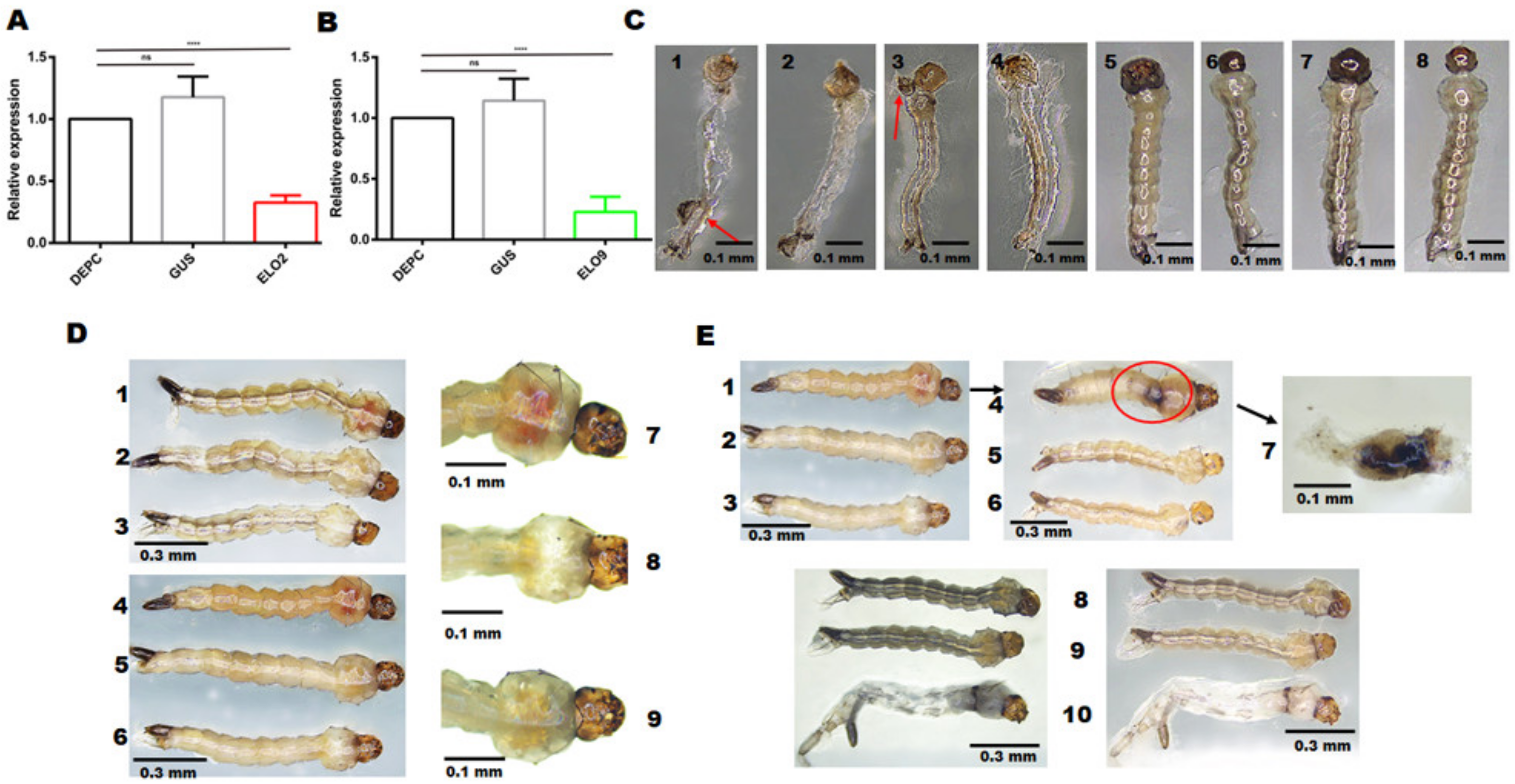
3.3. RNAi-Mediated Knockdown of AeELO2 Affects Larval Molting Behavior
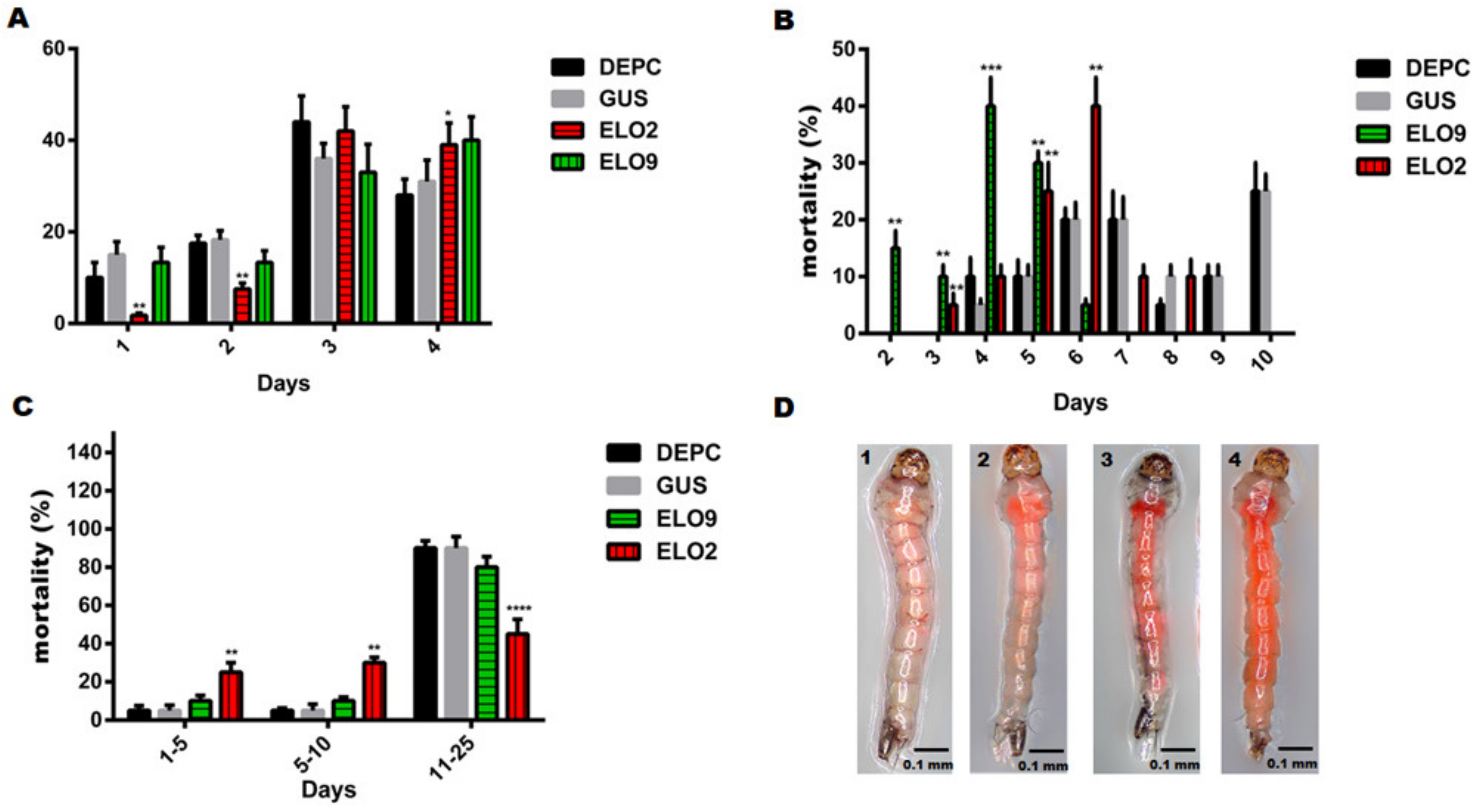
3.4. The Effects of AeELO2 and AeELO9 on Larval Development and Freezing Tolerance
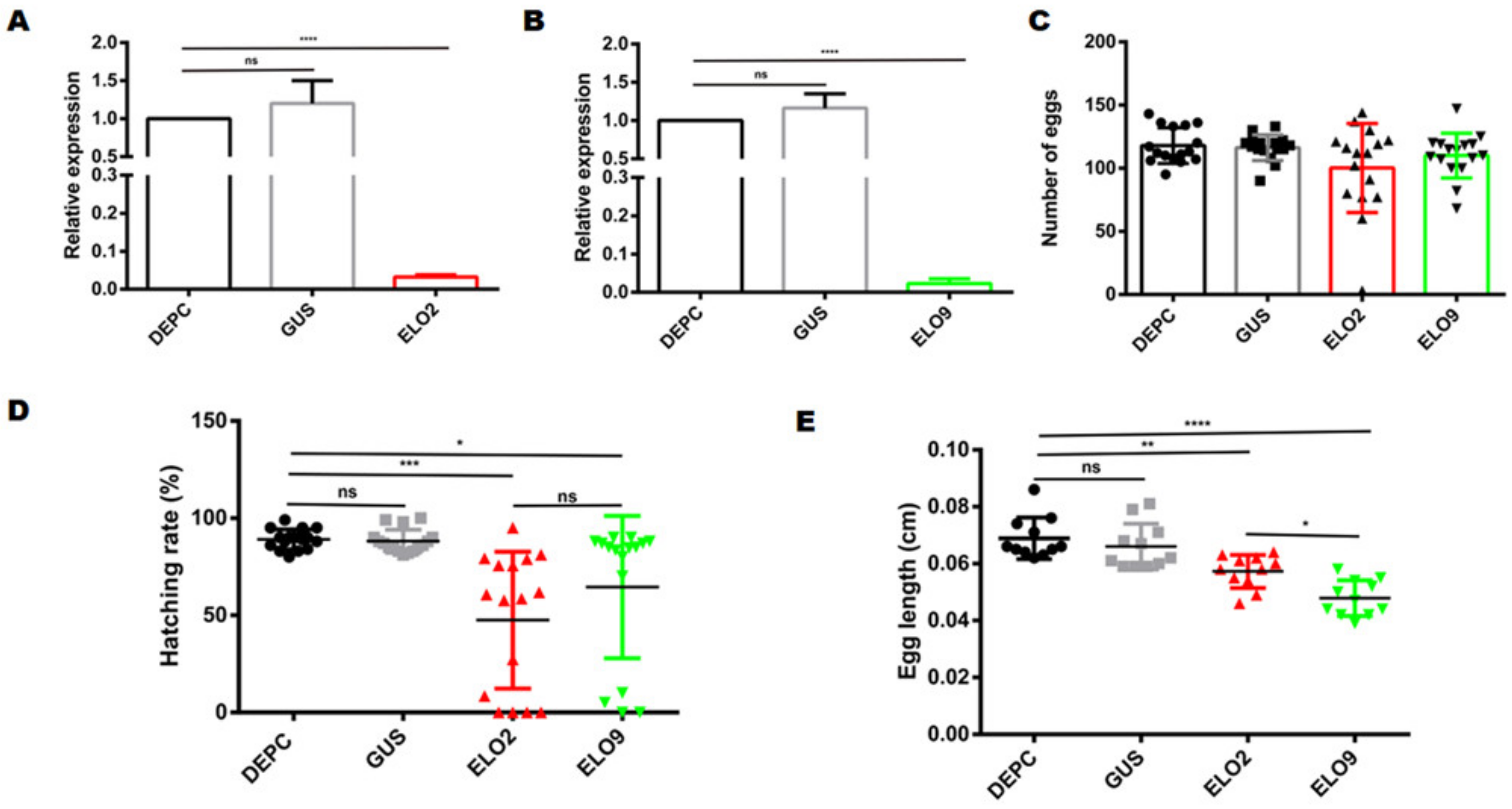
3.5. Decreased AeELO2 and AeELO9 Expression Inhibited Egg Hatching
3.6. Inhibiting the Expression of AeELO2 and AeELO9 Interfered with Cuticle Elasticity and Osmotic Pressure during Adult and Egg Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Qiao, J.W.; Fan, Y.L.; Wu, B.J.; Bai, T.T.; Wang, Y.H.; Zhang, Z.F.; Wang, D.; Liu, T.X. Downregulation of NADPH-cytochrome P450 reductase via RNA interference increases the susceptibility of Acyrthosiphon pisum to desiccation and insecticides. Insect Sci. 2021, 29, 1105–1119. [Google Scholar] [CrossRef]
- Chung, H.; Loehlin, D.W.; Dufour, H.D.; Vaccarro, K.; Millar, J.G.; Carroll, S.B. A single gene affects both ecological divergence and mate choice in Drosophila. Science 2014, 343, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; Le Goff, G.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, G.J.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef]
- Guillou, H.; Zadravec, D.; Martin, P.G.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Chertemps, T.; Duportets, L.; Labeur, C.; Ueda, R.; Takahashi, K.; Saigo, K.; Wicker-Thomas, C. A female-biased expressed elongase involved in long-chain hydrocarbon biosynthesis and courtship behavior in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2007, 104, 4273–4278. [Google Scholar] [CrossRef]
- Parvy, J.P.; Napal, L.; Rubin, T.; Poidevin, M.; Perrin, L.; Wicker-Thomas, C.; Montagne, J. Drosophila melanogaster Acetyl-CoA-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system. PLoS Genet. 2012, 8, e1002925. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.; Hollmann, M.; Schafer, M.A. The fatty acid elongase NOA is necessary for viability and has a somatic role in Drosophila sperm development. J. Cell Sci. 2007, 120, 2924–2934. [Google Scholar] [CrossRef]
- Zheng, T.; Li, H.; Han, N.; Wang, S.; Hackney Price, J.; Wang, M.; Zhang, D. Functional Characterization of Two Elongases of Very Long-Chain Fatty Acid from Tenebrio molitor L. (Coleopt: Tenebrionidae). Sci. Rep. 2017, 7, 10990. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.J.; Tong, Z.; Yang, Z.; Kaminoh, J.; Kamiyah, S.; Chen, H.; Zeng, J.; Chen, Y.; Luo, L.; Zhang, K. Essential role of Elovl4 in very long chain fatty acid synthesis, skin permeability barrier function, and neonatal survival. Int. J. Biol. Sci. 2007, 3, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, J.; Wilson, E.H.; Masek, K.; Striepen, B. Apicoplast fatty acid synthesis is essential for organelle biogenesis and parasite survival in Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 2006, 103, 13192–13197. [Google Scholar] [CrossRef] [Green Version]
- Li, D.T.; Chen, X.; Wang, X.Q.; Moussian, B.; Zhang, C.X. The fatty acid elongase gene family in the brown planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2019, 108, 32–43. [Google Scholar] [CrossRef]
- Rohrig, F.; Schulze, A. The multifaceted roles of fatty acid synthesis in cancer. Nat. Rev. Cancer 2016, 16, 732–749. [Google Scholar] [CrossRef]
- Xie, J.; Peng, G.; Wang, M.; Zhong, Q.; Song, X.; Bi, J.; Tang, J.; Feng, F.; Gao, H.; Li, B. RR-1 cuticular protein TcCPR69 is required for growth and metamorphosis in Tribolium castaneum. Insect Sci. 2022, 29, 1612–1628. [Google Scholar] [CrossRef]
- Vannini, L.; Willis, J.H. Localization of RR-1 and RR-2 cuticular proteins within the cuticle of Anopheles gambiae. Arthropod. Struct. Dev. 2017, 46, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lu, H.R.; Zhang, L.; Liao, C.H.; Han, Q. RNA interference-mediated knockdown of 3, 4-dihydroxyphenylacetaldehyde synthase affects larval development and adult survival in the mosquito Aedes aegypti. Parasit. Vectors 2019, 12, 311. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Upadhyay, A.; Liang, J.; Han, Q.; Li, J. 3,4-Dihydroxyphenylacetaldehyde synthase and cuticle formation in insects. Dev. Comp. Immunol. 2018, 83, 44–50. [Google Scholar] [CrossRef]
- Mun, S.; Noh, M.Y.; Dittmer, N.T.; Muthukrishnan, S.; Kramer, K.J.; Kanost, M.R.; Arakane, Y. Cuticular protein with a low complexity sequence becomes cross-linked during insect cuticle sclerotization and is required for the adult molt. Sci. Rep. 2015, 5, 10484. [Google Scholar] [CrossRef]
- Plotegher, N.; Berti, G.; Ferrari, E.; Tessari, I.; Zanetti, M.; Lunelli, L.; Greggio, E.; Bisaglia, M.; Veronesi, M.; Girotto, S.; et al. DOPAL derived alpha-synuclein oligomers impair synaptic vesicles physiological function. Sci. Rep. 2017, 7, 40699. [Google Scholar] [CrossRef] [PubMed]
- Vavricka, C.; Han, Q.; Huang, Y.; Erickson, S.M.; Harich, K.; Christensen, B.M.; Li, J. From L-dopa to dihydroxyphenylacetaldehyde: A toxic biochemical pathway plays a vital physiological function in insects. PLoS ONE 2011, 6, e16124. [Google Scholar] [CrossRef]
- Alabaster, A.; Isoe, J.; Zhou, G.; Lee, A.; Murphy, A.; Day, W.A.; Miesfeld, R.L. Deficiencies in acetyl-CoA carboxylase and fatty acid synthase 1 differentially affect eggshell formation and blood meal digestion in Aedes aegypti. Insect Biochem. Mol. Biol. 2011, 41, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Mysore, K.; Andrews, E.; Li, P.; Duman-Scheel, M. Chitosan/siRNA nanoparticle targeting demonstrates a requirement for single-minded during larval and pupal olfactory system development of the vector mosquito Aedes aegypti. BMC Dev. Biol. 2014, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Chertemps, T.; Duportets, L.; Labeur, C.; Wicker-Thomas, C. A new elongase selectively expressed in Drosophila male reproductive system. Biochem. Biophys. Res. Commun. 2005, 333, 1066–1072. [Google Scholar] [CrossRef] [PubMed]
- Schackmann, M.J.; Ofman, R.; Dijkstra, I.M.; Wanders, R.J.; Kemp, S. Enzymatic characterization of ELOVL1, a key enzyme in very long-chain fatty acid synthesis. Biochim. Biophys. Acta 2015, 1851, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.S.; Toke, D.A.; Mandala, S.; Martin, C.E. ELO2 and ELO3, homologues of the Saccharomyces cerevisiae ELO1 gene, function in fatty acid elongation and are required for sphingolipid formation. J. Biol. Chem. 1997, 272, 17376–17384. [Google Scholar] [CrossRef]
- Dzaki, N.; Ramli, K.N.; Azlan, A.; Ishak, I.H.; Azzam, G. Evaluation of reference genes at different developmental stages for quantitative real-time PCR in Aedes aegypti. Sci. Rep. 2017, 7, 43618. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, Y.; Niu, N.; Zhao, Y.; Liu, W.; Ma, E.; Moussian, B.; Zhang, J. The fatty acid elongase gene LmELO7 is required for hydrocarbon biosynthesis and cuticle permeability in the migratory locust, Locusta migratoria. J. Insect Physiol. 2020, 123, 104052. [Google Scholar] [CrossRef]
- Noh, M.Y.; Muthukrishnan, S.; Kramer, K.J.; Arakane, Y. Development and ultrastructure of the rigid dorsal and flexible ventral cuticles of the elytron of the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2017, 91, 21–33. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Niu, N.; Zhao, Y.; Liu, W.; Moussian, B.; Zhang, J. Two fatty acid synthase genes from the integument contribute to cuticular hydrocarbon biosynthesis and cuticle permeability in Locusta migratoria. Insect Mol. Biol. 2020, 29, 555–568. [Google Scholar] [CrossRef]
- Laws, K.M.; Drummond-Barbosa, D. Control of Germline Stem Cell Lineages by Diet and Physiology. Results Probl. Cell Differ. 2017, 59, 67–99. [Google Scholar]
- Almeida-Oliveira, F.; Tuthill, B.F., II; Gondim, K.C.; Majerowicz, D.; Musselman, L.P. dHNF4 regulates lipid homeostasis and oogenesis in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2021, 133, 103569. [Google Scholar] [CrossRef]
- Speake, B.K.; Murray, A.M.; Noble, R.C. Transport and transformations of yolk lipids during development of the avian embryo. Prog. Lipid Res. 1998, 37, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Speake, B.K.; Thompson, M.B. Lipids of the eggs and neonates of oviparous and viviparous lizards. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2000, 127, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.C.; Cocchi, M. Lipid metabolism and the neonatal chicken. Prog. Lipid Res. 1990, 29, 107–140. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Wu, Y.-C.; Chen, J.-K.; Zhu, X.-J.; Merkler, D.; Liao, C.-H.; Han, Q. Elongases of Long-Chain Fatty Acids ELO2 and ELO9 Are Involved in Cuticle Formation and Function in Fecundity in the Yellow Fever Mosquito, Aedes aegypti. Insects 2023, 14, 189. https://doi.org/10.3390/insects14020189
Chen J, Wu Y-C, Chen J-K, Zhu X-J, Merkler D, Liao C-H, Han Q. Elongases of Long-Chain Fatty Acids ELO2 and ELO9 Are Involved in Cuticle Formation and Function in Fecundity in the Yellow Fever Mosquito, Aedes aegypti. Insects. 2023; 14(2):189. https://doi.org/10.3390/insects14020189
Chicago/Turabian StyleChen, Jing, Yu-Chen Wu, Jiu-Kai Chen, Xiao-Jing Zhu, David Merkler, Cheng-Hong Liao, and Qian Han. 2023. "Elongases of Long-Chain Fatty Acids ELO2 and ELO9 Are Involved in Cuticle Formation and Function in Fecundity in the Yellow Fever Mosquito, Aedes aegypti" Insects 14, no. 2: 189. https://doi.org/10.3390/insects14020189