
Leaf Nutritional Content, Tree Richness, and Season Shape the Caterpillar Functional Trait Composition Hosted by Trees
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Region and Experimental Design
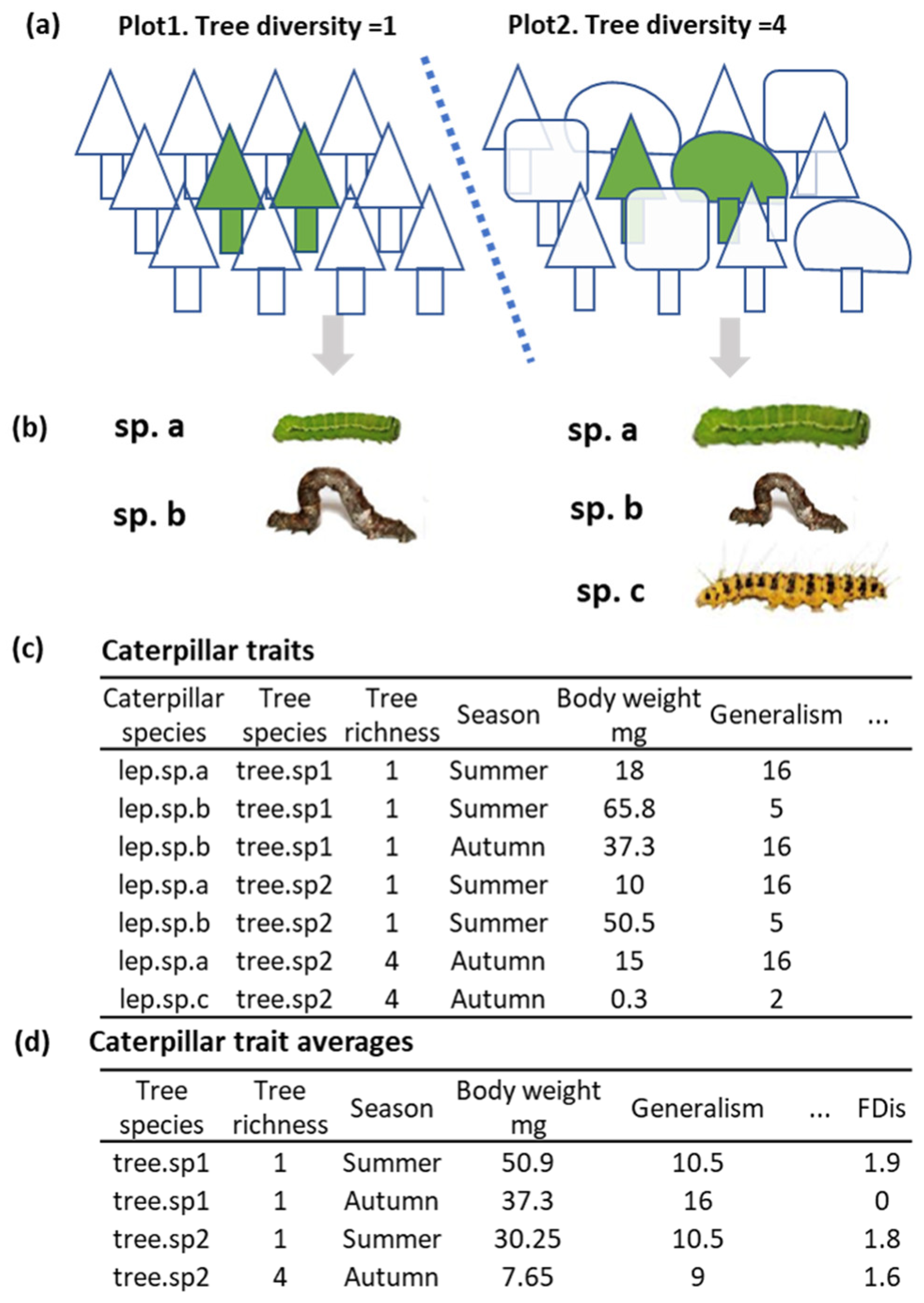
2.2. Sampling Strategy
2.3. Caterpillar Trait Measurements
2.4. Leaf Trait Selection
2.5. Statistical Analyses
2.5.1. Caterpillar Trait Data Processing and Linear Analysis
2.5.2. Caterpillar Functional Diversity within Tree Species
2.5.3. Caterpillar Trait Variation and Species-Sharing between Tree Species
2.5.4. Caterpillar Intra- and Inter-Specific Trait Variation
3. Results
3.1. Caterpillar Traits
3.1.1. Diversity and Generalism
3.1.2. Growth-Related Traits
3.1.3. Defensive Traits
3.1.4. Functional Diversity within Tree Species and Sensitivity Comparison
3.2. Functional Trait Variation and Species Sharing between Tree Species
4. Discussion
4.1. Tree Richness Effects on Caterpillar Trait Composition
4.2. Nutritional and Seasonal Effects on Caterpillar Traits
4.3. Inter- vs. Intra-Specific Trait Variation in Caterpillars
4.4. Methodological Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Caterpillar Species Richness SD | Abundance SD | |||||
|---|---|---|---|---|---|---|
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.996 ± 0.089 | 11.141 | <0.001 | 1.459 ± 0.097 | 15.026 | <0.001 |
| Tree replicates | 0.006 ± 0.130 | 0.047 | 0.963 | - | - | - |
| Caterpillar species richness | - | - | - | 0.451 ± 0.115 | 3.937 | 0.002 |
| Sampling season | −0.017 ± 0.093 | −0.184 | 0.857 | −0.117 ± 0.105 | −1.110 | 0.291 |
| Tree richness | −0.047 ± 0.130 | −0.363 | 0.724 | −0.004 ± 0.110 | −0.036 | 0.972 |
| Caterpillar FDis SD | Caterpillar generalism SD | |||||
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.662 ± 0.031 | 21.122 | <0.001 | 1.41 ± 0.042 | 33.862 | <0.001 |
| Caterpillar species richness | −0.132 ± 0.037 | −3.575 | 0.004 | −0.078 ± 0.049 | −1.580 | 0.142 |
| Sampling season | 0.019 ± 0.034 | 0.550 | 0.594 | 0.126 ± 0.045 | 2.772 | 0.018 |
| Tree species richness | −0.075 ± 0.036 | −2.115 | 0.058 | 0.030 ± 0.048 | 0.631 | 0.541 |
| Caterpillar body weight SD | Caterpillar head capsule width SD | |||||
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.550 ± 0.042 | 13.063 | <0.001 | 0.322 ± 0.020 | 15.998 | <0.001 |
| Caterpillar species richness | −0.129 ± 0.050 | −2.587 | 0.025 | −0.096 ± 0.024 | −4.039 | 0.002 |
| Sampling season | 0.016 ± 0.046 | 0.351 | 0.733 | −0.028 ± 0.022 | −1.291 | 0.223 |
| Tree species richness | −0.092 ± 0.048 | −1.930 | 0.080 | −0.067 ± 0.023 | −2.934 | 0.014 |
| Caterpillar relative head capsule width SD | ||||||
| Est ± SE | t | p | ||||
| (Intercept) | 0.096 ± 0.011 | 8.763 | <0.001 | |||
| Caterpillar species richness | −0.016 ± 0.016 | −1.007 | 0.337 | |||
| Caterpillar body weight | 0.004 ± 0.015 | 0.254 | 0.805 | |||
| Sampling season | <0.001 ± 0.012 | 0.007 | 0.995 | |||
| Tree species richness | 0.002 ± 0.014 | 0.169 | 0.869 | |||
| Caterpillar hair coverage SD | Caterpillar aposematism SD | |||||
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.615 ± 0.023 | 26.604 | <0.001 | 0.192 ± 0.011 | 17.872 | <0.001 |
| Caterpillar species richness | −0.132 ± 0.027 | −4.825 | 0.001 | −0.066 ± 0.013 | −5.177 | <0.001 |
| Sampling season | −0.006 ± 0.025 | −0.221 | 0.829 | 0.067 ± 0.012 | 5.737 | <0.001 |
| Tree species richness | −0.057 ± 0.026 | −2.161 | 0.054 | −0.026 ± 0.012 | −2.159 | 0.054 |
| Caterpillar Species Turnover | Caterpillar Species Nestedness | |||||
|---|---|---|---|---|---|---|
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.627 ± 0.005 | 126.823 | <0.001 | 0.043 ± 0.005 | 9.471 | <0.001 |
| Number of sampled plots | −0.012 ± 0.007 | −1.769 | 0.105 | 0.007 ± 0.006 | 1.165 | 0.269 |
| Sampling season | 0.016 ± 0.005 | 3.183 | 0.009 | −0.009 ± 0.005 | −1.863 | 0.089 |
| Tree species richness | −0.023 ± 0.007 | −3.340 | 0.007 | 0.012 ± 0.006 | 1.827 | 0.095 |
| Caterpillar beta diversity | ||||||
| Est ± SE | t | p | ||||
| (Intercept) | 0.651 ± 0.003 | 225.127 | <0.001 | |||
| Number of sampled plots | −0.008 ± 0.004 | −1.983 | 0.073 | |||
| Sampling season | 0.011 ± 0.003 | 3.776 | 0.003 | |||
| Tree species richness | −0.016 ± 0.004 | −4.029 | 0.002 | |||
References
- Haddad, N.M.; Crutsinger, G.M.; Gross, K.; Haarstad, J.; Knops, J.M.H.; Tilman, D. Plant Species Loss Decreases Arthropod Diversity and Shifts Trophic Structure. Ecol. Lett. 2009, 12, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Schuldt, A.; Ebeling, A.; Kunz, M.; Staab, M.; Guimarães-Steinicke, C.; Bachmann, D.; Buchmann, N.; Durka, W.; Fichtner, A.; Fornoff, F.; et al. Multiple Plant Diversity Components Drive Consumer Communities across Ecosystems. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond Species: Functional Diversity and the Maintenance of Ecological Processes and Services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Mattson, W.J. Herbivory in Relation to Plant Nitrogen Content. Annu. Rev. Ecol. Syst. 1980, 11, 119–161. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Diaz, S.; Yendramini, F.; Cornelissen, J.H.C.; Gurvich, D.E.; Cabido, M. Leaf Traits and Herbivore Selection in the Field and in Cafeteria Experiments. Austral Ecol. 2008, 28, 642–650. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a Plant-Arthropod Association in Simple and Diverse Habitats: The Fauna of Collards (Brassica Oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Karban, R.; Karban, C.; Huntzinger, M.; Pearse, I.; Crutsinger, G. Diet Mixing Enhances the Performance of a Generalist Caterpillar, Platyprepia Virginalis. Ecol. Entomol. 2010, 35, 92–99. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Whalen, M.A.; Davenport, T.M.; Stone, J.P.; Duffy, J.E. Physiological Effects of Diet Mixing on Consumer Fitness: A Meta-Analysis. Ecology 2013, 94, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Simpson, S.J.; Simmonds, M.S.J.; Blaney, W.M. A Comparison of Dietary Selection Behaviour in Larval Locusta Migratoria and Spodoptera Littoralis. Physiol. Entomol. 1988, 13, 225–238. [Google Scholar] [CrossRef]
- Despland, E.; Noseworthy, M. How Well Do Specialist Feeders Regulate Nutrient Intake? Evidence from a Gregarious Tree-Feeding Caterpillar. J. Exp. Biol. 2006, 209, 1301–1309. [Google Scholar] [CrossRef]
- Bernays, E.A.; Minkenberg, O.P.J.M. Insect Herbivores: Different Reasons for Being a Generalist. Ecology 1997, 78, 1157. [Google Scholar] [CrossRef]
- Werner, E.E.; Hall, D.J. Optimal Foraging and the Size Selection of Prey by the Bluegill Sunfish (Lepomis Macrochirus). Ecology 1974, 55, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Belovsky, G.E. Herbivore Optimal Foraging: A Comparative Test of Three Models. Am. Nat. 1984, 124, 97–115. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Simpson, S.J.; Mayntz, D. Nutrition, Ecology and Nutritional Ecology: Toward an Integrated Framework. Funct. Ecol. 2009, 23, 4–16. [Google Scholar] [CrossRef]
- Novotny, V.; Drozd, P.; Miller, S.E.; Kulfan, M.; Janda, M.; Basset, Y.; Weiblen, G.D. Why Are There So Many Species of Herbivorous Insects in Tropical Rainforests? Science 2006, 313, 1115–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coley, P.D.; Bateman, M.L.; Kursar, T.A. The Effects of Plant Quality on Caterpillar Growth and Defense against Natural Enemies. Oikos 2006, 115, 219–228. [Google Scholar] [CrossRef]
- Scriber, J.M.; Slansky, F. The Nutritional Ecology of Immature Insects. Annu. Rev. Entomol. 1981, 26, 183–211. [Google Scholar] [CrossRef]
- Scheirs, J.; De Bruyn, L. Integrating Optimal Foraging and Optimal Oviposition Theory in Plant-Insect Research. Oikos 2002, 96, 187–191. [Google Scholar] [CrossRef]
- Mitchell, R.G. Dispersal of Early Instars of the Douglas-Fir Tussock Moth1,2,3. Ann. Entomol. Soc. Am. 1979, 72, 291–297. [Google Scholar] [CrossRef]
- White, S.A.; Deacon, A.E. Where One Species Leads, Another Follows: Interspecies Processions in Tropical Caterpillars. Ecology 2021, 102, 2. [Google Scholar] [CrossRef]
- Janz, N. Evolutionary Ecology of Oviposition Strategies. In Chemoecology of Insect Eggs and Egg Deposition; Blackwell Publishing Ltd.: Oxford, UK, 2003; pp. 349–376. [Google Scholar]
- Behmer, S.T. Insect Herbivore Nutrient Regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P.; Cohen, R.W.; Friedman, S. Self-Selection of an Optimal Nutrient Mix from Defined Diets by Larvae of the Corn Earworm, Heliothis Zea (Boddie). Physiol. Zool. 1984, 57, 590–597. [Google Scholar] [CrossRef]
- Lee, K.P.; Cory, J.S.; Wilson, K.; Raubenheimer, D.; Simpson, S.J. Flexible Diet Choice Offsets Protein Costs of Pathogen Resistance in a Caterpillar. Proc. R. Soc. B Biol. Sci. 2006, 273, 823–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockhoff, B.A. Ontogenetic Change in Dietary Selection for Protein and Lipid by Gypsy Moth Larvae. J. Insect Physiol. 1993, 39, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J.; Reavey, D.; Valladares, G.R. Changes in Feeding Habit as Caterpillars Grow. Ecol. Entomol. 1991, 16, 339–344. [Google Scholar] [CrossRef]
- Gentry, G.L.; Dyer, L.A. On the Conditional Nature of Neotropical Caterpillar Defenses against Their Natural Enemies. Ecology 2002, 83, 3108. [Google Scholar] [CrossRef]
- Ohsaki, N.; Sato, Y. Food Plant Choice of Pieris Butterflies as a Trade-Off between Parasitoid Avoidance and Quality of Plants. Ecology 1994, 75, 59–68. [Google Scholar] [CrossRef]
- Singer, M.S.; Carrière, Y.; Theuring, C.; Hartmann, T. Disentangling Food Quality from Resistance against Parasitoids: Diet Choice by a Generalist Caterpillar. Am. Nat. 2004, 164, 423–429. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional Constraints in Terrestrial and Freshwater Food Webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- Chapman, R.F. The Insects: Structure and Function, 5th ed.; Simpson, S.J., Douglas, A.E., Eds.; Cambridge University Press: Cambridge, UK, 2013; ISBN 9780521113892. [Google Scholar]
- Raubenheimer, D.; Simpson, S.J. Nutrient Balancing in Grasshoppers: Behavioural and Physiological Correlates of Dietary Breadth. J. Exp. Biol. 2003, 206, 1669–1681. [Google Scholar] [CrossRef]
- Bernays, E.A. Evolution of Feeding Behavior in Insect Herbivores. Bioscience 1998, 48, 35–44. [Google Scholar] [CrossRef]
- Bernays, E.A.; Janzen, D.H. Saturniid and Sphingid Caterpillars: Two Ways to Eat Leaves. Ecology 1988, 69, 1153–1160. [Google Scholar] [CrossRef]
- Brousseau, P.-M.; Gravel, D.; Handa, I.T. On the Development of a Predictive Functional Trait Approach for Studying Terrestrial Arthropods. J. Anim. Ecol. 2018, 87, 1209–1220. [Google Scholar] [CrossRef]
- Hochuli, D.F. Insect Herbivory and Ontogeny: How Do Growth and Development Influence Feeding Behaviour, Morphology and Host Use? Austral Ecol. 2001, 26, 563–570. [Google Scholar] [CrossRef]
- Chavana-Bryant, C.; Malhi, Y.; Wu, J.; Asner, G.P.; Anastasiou, A.; Enquist, B.J.; Cosio Caravasi, E.G.; Doughty, C.E.; Saleska, S.R.; Martin, R.E.; et al. Leaf Aging of Amazonian Canopy Trees as Revealed by Spectral and Physiochemical Measurements. New Phytol. 2017, 214, 1049–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes, J.; Garcia, S.; Grandis, A.; Nascimento, H.; Domingues, T.F.; Guedes, A.V.; Aleixo, I.; Camargo, P.; Campos, J.; Damasceno, A.; et al. Changes in Leaf Functional Traits with Leaf Age: When Do Leaves Decrease Their Photosynthetic Capacity in Amazonian Trees? Tree Physiol. 2022, 42, 922–938. [Google Scholar] [CrossRef]
- Stamp, N.E.; Bowers, M.D. Phenology of Nutritional Differences between New and Mature Leaves and Its Effect on Caterpillar Growth. Ecol. Entomol. 1990, 15, 447–454. [Google Scholar] [CrossRef]
- Čížek, L. Diet Composition and Body Size in Insect Herbivores: Why Do Small Species Prefer Young Leaves? Eur. J. Entomol. 2005, 102, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Hauer-Jákli, M.; Tränkner, M. Critical Leaf Magnesium Thresholds and the Impact of Magnesium on Plant Growth and Photo-Oxidative Defense: A Systematic Review and Meta-Analysis From 70 Years of Research. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Ji, M.; Jin, G.; Liu, Z. Effects of Ontogenetic Stage and Leaf Age on Leaf Functional Traits and the Relationships between Traits in Pinus Koraiensis. J. For. Res. 2021, 32, 2459–2471. [Google Scholar] [CrossRef]
- Wallace, A.; Mueller, R.T. Calcium Uptake and Distribution in Plants. J. Plant Nutr. 1980, 2, 247–256. [Google Scholar] [CrossRef]
- Wyatt, G.R. The Biochemistry of Insect Hemolymph. Annu. Rev. Entomol. 1961, 6, 75–102. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, M.; Chen, Y.; Yin, D.; He, D.; Fang, S.; Ellwood, M.D.F.; Chu, C. Plant-caterpillar Food Web: Integrating Leaf Stoichiometry and Phylogeny. Ecol. Entomol. 2021, 46, 1026–1035. [Google Scholar] [CrossRef]
- Prather, R.M.; Castillioni, K.; Kaspari, M.; Souza, L.; Prather, C.M.; Reihart, R.W.; Welti, E.A.R. Micronutrients Enhance Macronutrient Effects in a Meta-analysis of Grassland Arthropod Abundance. Glob. Ecol. Biogeogr. 2020, 29, 2273–2288. [Google Scholar] [CrossRef]
- Usherwood, P.N.R. Insect Neuromuscular Mechanisms. Am. Zool. 1967, 7, 553–582. [Google Scholar] [CrossRef] [Green Version]
- Loader, C.; Damman, H. Nitrogen Content of Food Plants and Vulnerability of Pieris Rapae to Natural Enemies. Ecology 1991, 72, 1586–1590. [Google Scholar] [CrossRef]
- Bowers, M.D. Aposematic Caterpillars: Life-Styles of the Warningly Colored and Unpalatable. In Caterpillars: Ecological and Evolutionary Constraints on Foraging; Stamp, N.E., Casey, T.M., Eds.; Chapman & Hall: New York, NY, USA, 1993; pp. 331–371. [Google Scholar]
- Sugiura, S.; Yamazaki, K. Caterpillar Hair as a Physical Barrier against Invertebrate Predators. Behav. Ecol. 2014, 25, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Higginson, A.D.; Ruxton, G.D. Optimal Defensive Coloration Strategies during the Growth Period of Prey. Evolution 2010, 64, 53–67. [Google Scholar] [CrossRef]
- Grant, J.B. Ontogenetic Colour Change and the Evolution of Aposematism: A Case Study in Panic Moth Caterpillars. J. Anim. Ecol. 2007, 76, 439–447. [Google Scholar] [CrossRef]
- Mappes, J.; Kokko, H.; Ojala, K.; Lindström, L. Seasonal Changes in Predator Community Switch the Direction of Selection for Prey Defences. Nat. Commun. 2014, 5, 5016. [Google Scholar] [CrossRef]
- Bruelheide, H.; Nadrowski, K.; Assmann, T.; Bauhus, J.; Both, S.; Buscot, F.; Chen, X.; Ding, B.; Durka, W.; Erfmeier, A.; et al. Designing Forest Biodiversity Experiments: General Considerations Illustrated by a New Large Experiment in Subtropical China. Methods Ecol. Evol. 2014, 5, 74–89. [Google Scholar] [CrossRef] [Green Version]
- Davrinche, A.; Haider, S. Intra-Specific Leaf Trait Responses to Species Richness at Two Different Local Scales. Basic Appl. Ecol. 2021, 55, 20–32. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) Documentation of the of the BLASTCLUST-Algorithm. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 1 December 2019).
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Li, Y.; Chesters, D.; Anttonen, P.; Bruelheide, H.; Chen, J.; Durka, W.; Guo, P.; Härdtle, W.; Ma, K.; et al. Multiple Components of Plant Diversity Loss Determine Herbivore Phylogenetic Diversity in a Subtropical Forest Experiment. J. Ecol. 2019, 107, 2697–2712. [Google Scholar] [CrossRef] [Green Version]
- Ahrens, D.; Fujisawa, T.; Krammer, H.-J.; Eberle, J.; Fabrizi, S.; Vogler, A.P. Rarity and Incomplete Sampling in DNA-Based Species Delimitation. Syst. Biol. 2016, 65, 478–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyar, H.G. The Number of Molts of Lepidopterous Larvae. Psyche A J. Entomol. 1890, 5, 420–422. [Google Scholar] [CrossRef]
- Hutchinson, J.M.C.; McNamara, J.M.; Houston, A.I.; Vollrath, F. Dyar’s Rule and the Investment Principle: Optimal Moulting Strategies If Feeding Rate Is Size–Dependent and Growth Is Discontinuous. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 1997, 352, 113–138. [Google Scholar] [CrossRef]
- Hunter, A.F.; Elkinton, J.S. Effects of Synchrony with Host Plant on Populations of a Spring-Feeding Lepidopteran. Ecology 2000, 81, 1248. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New Handbook for Standardised Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2016, 64, 715. [Google Scholar] [CrossRef] [Green Version]
- Muiruri, E.W.; Barantal, S.; Iason, G.R.; Salminen, J.-P.; Perez-Fernandez, E.; Koricheva, J. Forest Diversity Effects on Insect Herbivores: Do Leaf Traits Matter? New Phytol. 2019, 221, 2250–2260. [Google Scholar] [CrossRef] [PubMed]
- Stamp, N.E.; Bowers, M.D. Variation in Food Quality and Temperature Constrain Foraging of Gregarious Caterpillars. Ecology 1990, 71, 1031–1039. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; ISBN 9781544336473. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A Protocol for Data Exploration to Avoid Common Statistical Problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95364-9. [Google Scholar]
- Laliberté, E.; Legendre, P. A Distance-Based Framework for Measuring Functional Diversity from Multiple Traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; Logez, M. Betapart: Partitioning Beta Diversity into Turnover and Nestedness Components. 2021. Available online: https://cran.r-project.org/web/packages/betapart/betapart.pdf (accessed on 11 November 2022).
- Wang, M.; Li, Y.; Chesters, D.; Bruelheide, H.; Ma, K.; Guo, P.; Zhou, Q.; Staab, M.; Zhu, C.; Schuldt, A. Host Functional and Phylogenetic Composition Rather than Host Diversity Structure Plant–Herbivore Networks. Mol. Ecol. 2020, 29, 2747–2762. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yan, C.; Luo, A.; Li, Y.; Chesters, D.; Qiao, H.; Chen, J.; Zhou, Q.; Ma, K.; Bruelheide, H.; et al. Phylogenetic Relatedness, Functional Traits, and Spatial Scale Determine Herbivore Co-occurrence in a Subtropical Forest. Ecol. Monogr. 2022, 92, e01492. [Google Scholar] [CrossRef]
- Bihn, J.H.; Gebauer, G.; Brandl, R. Loss of Functional Diversity of Ant Assemblages in Secondary Tropical Forests. Ecology 2010, 91, 782–792. [Google Scholar] [CrossRef]
- Brousseau, P.; Gravel, D.; Handa, I.T. Traits of Litter-dwelling Forest Arthropod Predators and Detritivores Covary Spatially with Traits of Their Resources. Ecology 2019, 100, 1–13. [Google Scholar] [CrossRef]
- Guariento, E.; Strutzenberger, P.; Truxa, C.; Fiedler, K. The Trinity of Ecological Contrasts: A Case Study on Rich Insect Assemblages by Means of Species, Functional and Phylogenetic Diversity Measures. BMC Ecol. 2020, 20, 29. [Google Scholar] [CrossRef]
- Zhang, J.; Bruelheide, H.; Chen, X.; Eichenberg, D.; Kröber, W.; Xu, X.; Xu, L.; Schuldt, A. Tree Diversity Promotes Generalist Herbivore Community Patterns in a Young Subtropical Forest Experiment. Oecologia 2017, 183, 455–467. [Google Scholar] [CrossRef]
- Schoonhoven, L.M. What Makes a Caterpillar Eat? The Sensory Code Underlying Feeding Behavior. In Perspectives in Chemoreception and Behavior. Proceedings in Life Sciences; Chapman, R.F., Bernays, E.A., Stoffolano, J.J.G., Eds.; Springer: New York, NY, USA, 1987; pp. 69–97. [Google Scholar]
- Fiedler, K. Diet Breadth and Host Plant Diversity of Tropical- vs. Temperate-Zone Herbivores: South-East Asian and West Palaearctic Butterflies as a Case Study. Ecol. Entomol. 1998, 23, 285–297. [Google Scholar] [CrossRef]
- Novotny, V.; Basset, Y.; Miller, S.E.; Weiblen, G.D.; Bremer, B.; Cizek, L.; Drozd, P. Low Host Specificity of Herbivorous Insects in a Tropical Forest. Nature 2002, 416, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.R.; Morrow, P.A. Specialization: Species Property or Local Phenomenon? Science 1981, 211, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Russell, E.P. Enemies Hypothesis: A Review of the Effect of Vegetational Diversity on Predatory Insects and Parasitoids. Environ. Entomol. 1989, 18, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Staab, M.; Schuldt, A. The Influence of Tree Diversity on Natural Enemies—A Review of the “Enemies” Hypothesis in Forests. Curr. For. Rep. 2020, 6, 243–259. [Google Scholar] [CrossRef]
- Dyer, L.A. Tasty Generalists and Nasty Specialists? Antipredator Mechanisms in Tropical Lepidopteran Larvae. Ecology 1995, 76, 1483–1496. [Google Scholar] [CrossRef]
- Jactel, H.; Moreira, X.; Castagneyrol, B. Tree Diversity and Forest Resistance to Insect Pests: Patterns, Mechanisms, and Prospects. Annu. Rev. Entomol. 2021, 66, 277–296. [Google Scholar] [CrossRef]
- Teder, T.; Esperk, T.; Remmel, T.; Sang, A.; Tammaru, T. Counterintuitive Size Patterns in Bivoltine Moths: Late-Season Larvae Grow Larger despite Lower Food Quality. Oecologia 2010, 162, 117–125. [Google Scholar] [CrossRef]
- Seifert, C.L.; Jorge, L.R.; Volf, M.; Wagner, D.L.; Lamarre, G.P.A.; Miller, S.E.; Gonzalez-Akre, E.; Anderson-Teixeira, K.J.; Novotný, V. Seasonality Affects Specialisation of a Temperate Forest Herbivore Community. Oikos 2021, 130, 1450–1461. [Google Scholar] [CrossRef]
- Kaplan, I.; Denno, R.F. Interspecific Interactions in Phytophagous Insects Revisited: A Quantitative Assessment of Competition Theory. Ecol. Lett. 2007, 10, 977–994. [Google Scholar] [CrossRef]
- Denno, R.F.; McClure, M.S.; Ott, J.R. Interspecific Interactions in Phytophagous Insects: Competition Reexamined and Resurrected. Annu. Rev. Entomol. 1995, 40, 297–331. [Google Scholar] [CrossRef]
- Calvo, D.; Molina, J.M. Head Capsule Width and Instar Determination for Larvae of Streblote Panda (Lepidoptera: Lasiocampidae). Ann. Entomol. Soc. Am. 2008, 101, 881–886. [Google Scholar] [CrossRef]
- Jobin, L.; Hebért, C.; Bourassa, J.-P. Head Capsule Width of Larval Populations of the Gypsy Moth (Lepidoptera: Lymantriidae) in Quebec, with Reference to Dyar’s Hypothesis. Environ. Entomol. 1992, 21, 89–93. [Google Scholar] [CrossRef]
- Belovsky, G.E.; Slade, J.B. Insect Herbivory Accelerates Nutrient Cycling and Increases Plant Production. Proc. Natl. Acad. Sci. USA 2000, 97, 14412–14417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiss, J.; Bridle, J.R.; Montoya, J.M.; Woodward, G. Emerging Horizons in Biodiversity and Ecosystem Functioning Research. Trends Ecol. Evol. 2009, 24, 505–514. [Google Scholar] [CrossRef]
- Schmitz, O.J.; Buchkowski, R.W.; Burghardt, K.T.; Donihue, C.M. Functional Traits and Trait-Mediated Interactions. In Trait-Based Ecology—From Structure to Function; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; pp. 319–343. [Google Scholar]



| Caterpillar Species Richness | Caterpillar Abundance | |||||
|---|---|---|---|---|---|---|
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 1.382 ± 0.035 | 39.759 | <0.001 | 1.592 ± 0.018 | 88.861 | <0.001 |
| Tree replicates | 0.178 ± 0.039 | 4.607 | <0.001 | - | - | - |
| Caterpillar species richness | - | - | - | 0.656 ± 0.018 | 35.668 | <0.001 |
| Tree species richness | 0.101 ± 0.039 | 2.610 | 0.010 | - | - | - |
| Sampling season | - | - | - | 0.016 ± 0.018 | 0.863 | 0.390 |
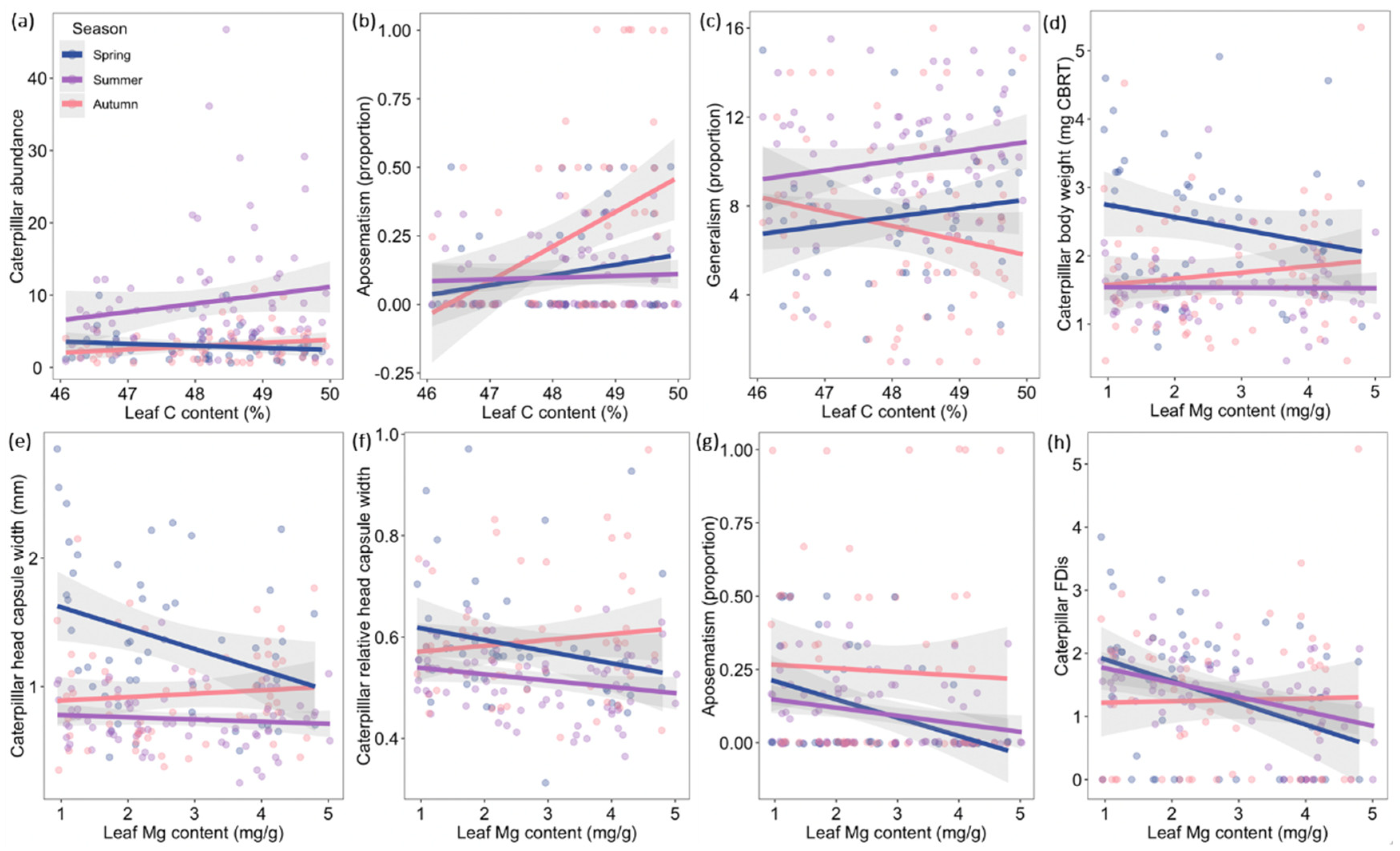
| Leaf C content | - | - | - | 0.030 ± 0.021 | 1.450 | 0.149 |
| Leaf Mg content | −0.097 ± 0.035 | −2.774 | 0.006 | 0.044 ± 0.021 | 2.073 | 0.040 |
| Season: leaf C content | - | - | - | 0.056 ± 0.018 | 3.070 | 0.002 |
| Caterpillar FDis | Caterpillar generalism | |||||
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.747 ± 0.025 | 29.911 | <0.001 | 2.164 ± 0.040 | 53.703 | <0.001 |
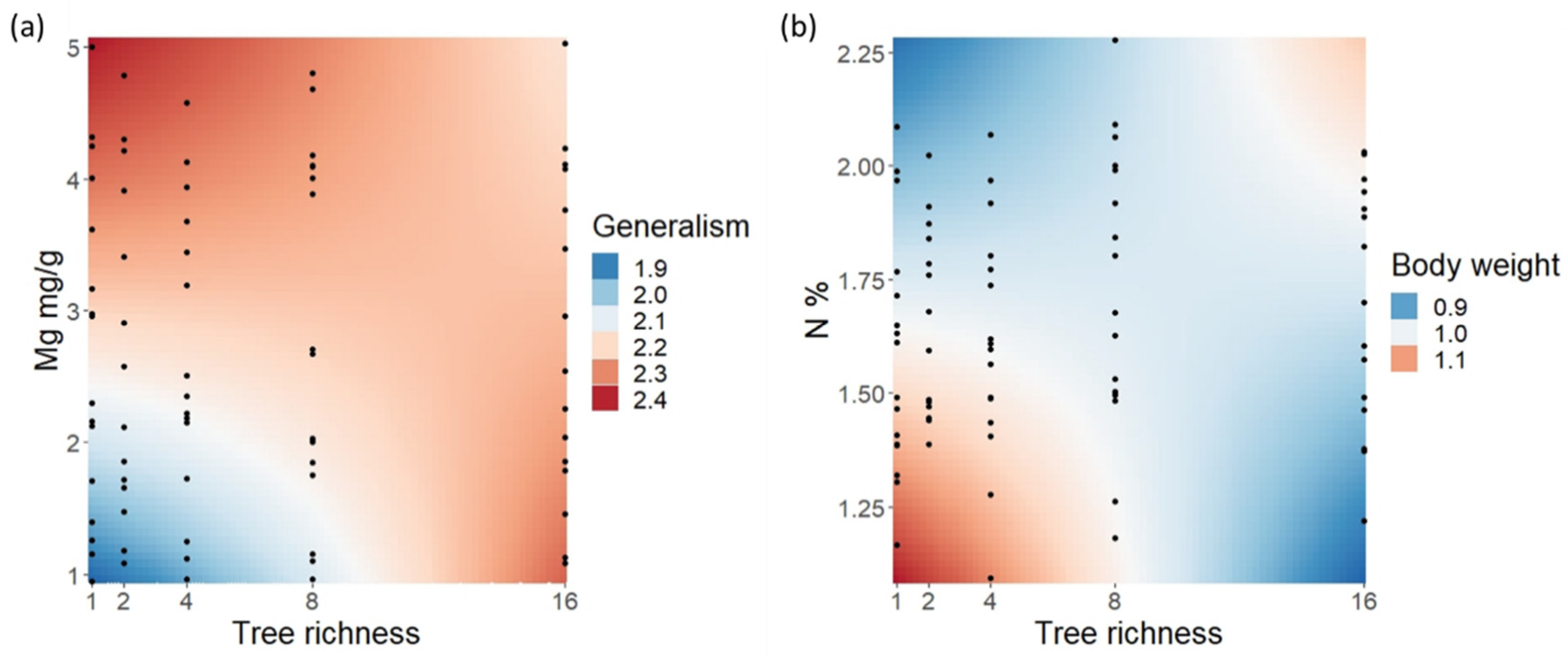
| Caterpillar species richness | 0.298 ± 0.026 | 11.640 | <0.001 | 0.098 ± 0.031 | 3.135 | 0.002 |
| Caterpillar body weight | - | - | - | −0.085 ± 0.032 | −2.643 | 0.009 |
| Sampling season | −0.004 ± 0.025 | −0.149 | 0.882 | −0.092 ± 0.032 | −2.890 | 0.004 |
| Tree species richness | - | - | - | 0.040 ± 0.030 | 1.328 | 0.186 |
| Leaf C content | - | - | - | 0.060 ± 0.044 | 1.359 | 0.187 |
| Leaf Mg content | −0.048 ± 0.026 | −1.885 | 0.061 | 0.099 ± 0.045 | 2.216 | 0.036 |
| Season: C | - | - | - | −0.074 ± 0.030 | −2.421 | 0.017 |
| Season: Mg | 0.063 ± 0.025 | 2.468 | 0.015 | - | - | - |
| Tree richness: Mg | - | - | - | −0.072 ± 0.032 | −2.276 | 0.024 |
| Caterpillar body weight | Caterpillar head capsule width | |||||
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.997 ± 0.019 | 51.275 | <0.001 | 0.655 ± 0.007 | 99.980 | <0.001 |
| Caterpillar body weight | - | - | - | 0.189 ± 0.006 | 30.412 | <0.001 |
| Caterpillar species richness | - | - | - | −0.023 ± 0.006 | −3.834 | <0.001 |
| Sampling season | −0.087 ± 0.019 | −4.537 | <0.001 | −0.007 ± 0.006 | −1.197 | 0.233 |
| Tree species richness | −0.018 ± 0.020 | −0.909 | 0.365 | - | - | - |
| Leaf C content | - | - | - | −0.019 ± 0.008 | −2.394 | 0.030 |
| Leaf N content | −0.033 ± 0.021 | −1.551 | 0.123 | −0.020 ± 0.007 | −2.641 | 0.015 |
| Leaf Mg content | −0.004 ± 0.021 | −0.204 | 0.839 | −0.025 ± 0.008 | −3.155 | 0.007 |
| Season: Mg | 0.043 ± 0.020 | 2.178 | 0.031 | 0.017 ± 0.006 | 2.950 | 0.004 |
| Tree richness: N | 0.040 ± 0.020 | 1.998 | 0.047 | - | - | - |
| Caterpillar relative head capsule width | ||||||
| Est ± SE | t | p | ||||
| (Intercept) | 0.440 ± 0.005 | 82.459 | <0.001 | |||
| Caterpillar species richness | −0.010 ± 0.005 | −2.082 | 0.039 | |||
| Caterpillar body weight | −0.017 ± 0.005 | −3.430 | <0.001 | |||
| Sampling season | −0.001 ± 0.005 | −0.216 | 0.829 | |||
| Leaf Mg content | −0.012 ± 0.005 | −2.298 | 0.033 | |||
| Season: Mg | 0.014 ± 0.005 | 2.902 | 0.004 | |||
| Caterpillar hair coverage | Caterpillar aposematism | |||||
| Est ± SE | t | p | Est ± SE | t | p | |
| (Intercept) | 0.565 ± 0.029 | 19.980 | <0.001 | 0.145 ± 0.014 | 11.245 | <0.001 |
| Caterpillar body weight | - | - | - | 0.038 ± 0.015 | 2.766 | 0.006 |
| Aposematism | 0.203 ± 0.030 | 6.796 | <0.001 | - | - | - |
| Hair coverage | - | - | - | 0.091 ± 0.015 | 6.430 | <0.001 |
| Sampling season | −0.142 ± 0.029 | −4.883 | <0.001 | 0.084 ± 0.015 | 5.510 | <0.001 |
| Leaf C content | −0.091 ± 0.033 | −2.775 | 0.006 | 0.054 ± 0.016 | 3.095 | 0.002 |
| Leaf N content | −0.074 ± 0.032 | −2.297 | 0.023 | - | - | - |
| Leaf Mg content | - | - | - | −0.003 ± 0.016 | −0.577 | 0.564 |
| Season: C | - | - | - | 0.058 ± 0.017 | 3.133 | 0.002 |
| Season: Mg | - | - | - | 0.043 ± 0.017 | 2.313 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anttonen, P.; Li, Y.; Chesters, D.; Davrinche, A.; Haider, S.; Bruelheide, H.; Chen, J.-T.; Wang, M.-Q.; Ma, K.-P.; Zhu, C.-D.; et al. Leaf Nutritional Content, Tree Richness, and Season Shape the Caterpillar Functional Trait Composition Hosted by Trees. Insects 2022, 13, 1100. https://doi.org/10.3390/insects13121100
Anttonen P, Li Y, Chesters D, Davrinche A, Haider S, Bruelheide H, Chen J-T, Wang M-Q, Ma K-P, Zhu C-D, et al. Leaf Nutritional Content, Tree Richness, and Season Shape the Caterpillar Functional Trait Composition Hosted by Trees. Insects. 2022; 13(12):1100. https://doi.org/10.3390/insects13121100
Chicago/Turabian StyleAnttonen, Perttu, Yi Li, Douglas Chesters, Andréa Davrinche, Sylvia Haider, Helge Bruelheide, Jing-Ting Chen, Ming-Qiang Wang, Ke-Ping Ma, Chao-Dong Zhu, and et al. 2022. "Leaf Nutritional Content, Tree Richness, and Season Shape the Caterpillar Functional Trait Composition Hosted by Trees" Insects 13, no. 12: 1100. https://doi.org/10.3390/insects13121100