
Generic Pheromones Identified from Northern Hemisphere Cerambycidae (Coleoptera) Are Attractive to Native Longhorn Beetles from Central-Southern Chile
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Studies, Localities, Seasons, and Site Characteristics
2.2. Traps, Compounds, and Treatments
2.3. Experimental Design, Results, and Statistical Analysis
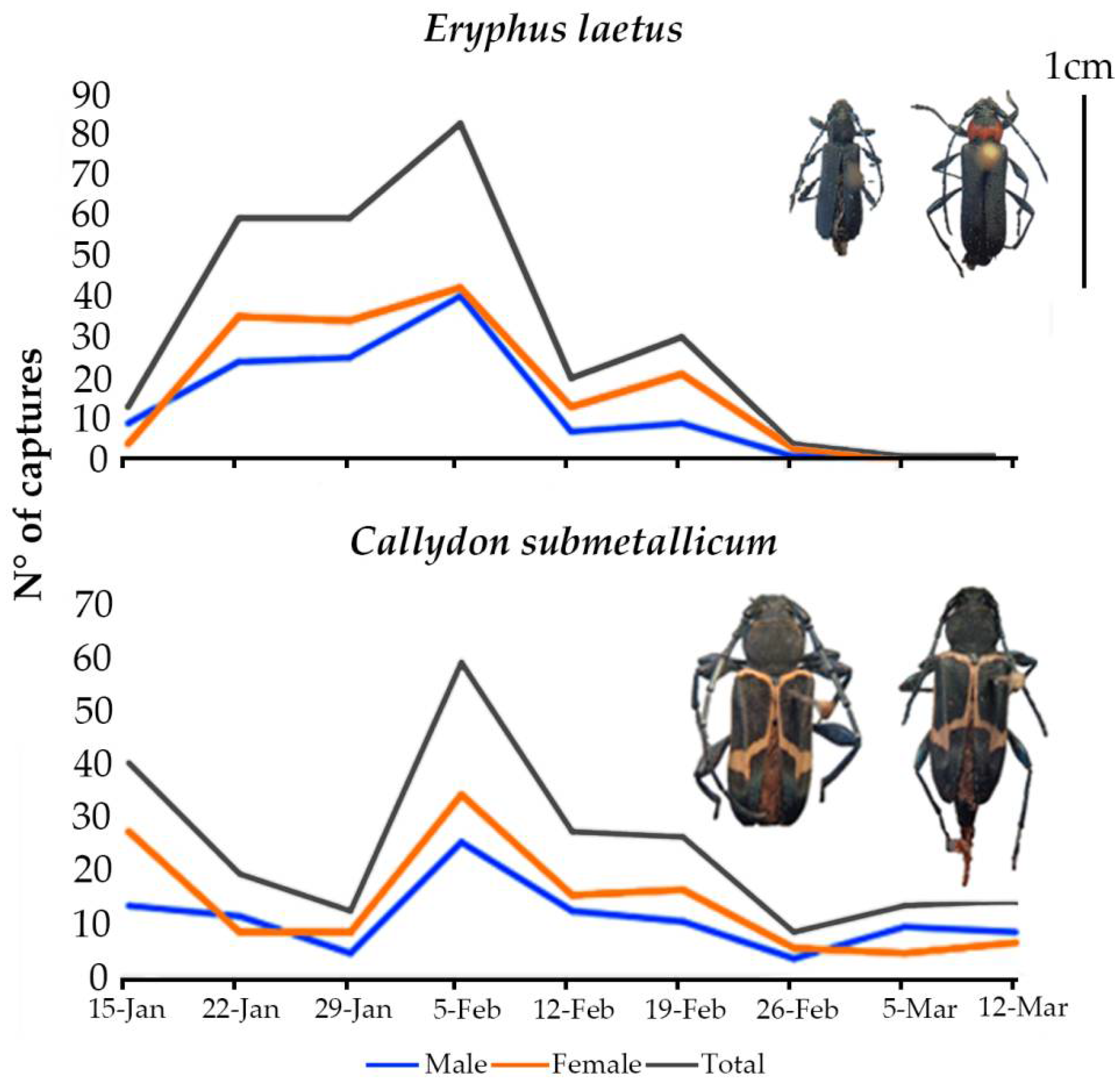
3. Results
3.1. Study 1
3.2. Study 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monnè, M.L.; Monnè, M.A.; Wang, Q. General morphology, classification, and biology of Cerambycidae. In Cerambycidae of the world: Biology and Pest Management, 1st ed.; Wang, Q., Ed.; CRC Press: Boca Raton, FL, USA, 2017; Chapter 1; pp. 1–70. [Google Scholar]
- Tavakilian, G.; Berkov, A.; Meurer-Grimes, B.; Mori, S. Neotropical tree species and their faunas of xylophagous longicorns (Coleoptera: Cerambycidae) in French Guiana. Bot. Rev. 1997, 63, 303–355. [Google Scholar] [CrossRef]
- Yasui, H.; Fujiwara-Tsujii, N.; Yasuda, T. Detection of volatile pheromone candidates from the white-spotted longicorn beetle, Anoplophora malasiaca (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2019, 54, 203–211. [Google Scholar] [CrossRef]
- Linsley, E.G. The Cerambycidae of North America: Part I—Introduction; University of California Press: Berkley, CA, USA, 1961; Volume 18, pp. 1–97. [Google Scholar]
- Iwabuchi, K. Mating behavior of Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae) 1. Behavioral sequences and existence of the male sex pheromone. Appl. Entomol. Zool. 1982, 17, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Fettköther, R.; Dettner, K.; Schröder, F.; Meyer, H.; Francke, W.; Noldt, U. The male pheromone of the old house borer Hylotrupes bajulus (L.) (Coleoptera: Cerambycidae): Identification and female response. Experientia 1995, 51, 270–277. [Google Scholar] [CrossRef]
- Allison, J.; Borden, J.; Seybold, S.A. A review of the chemical ecology of the Cerambycidae (Coleoptera). Evolutionary, mechanistic and environmental approaches to chemically-mediated interactions. Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Millar, J.G.; Hanks, L.M. Chemical Ecology of Cerambycids. In Cerambycidae of the World: Biology and Pest Management, 1st ed.; Wang, Q., Ed.; CRC Press: Boca Raton, FL, USA, 2017; Chapter 5; pp. 161–208. [Google Scholar]
- El-Sayed, A.M. The Pherobase: Database of Pheromones and Semiochemicals. 2022. Available online: https://www.pherobase.com (accessed on 30 September 2022).
- Cohen, C.; Liltved, W.R.; Colville, J.F.; Shuttleworth, A.; Weissflog, J.; Svatoš, A.; Bytebier, B.; Johnson, S.D. Sexual deception of a beetle pollinator through floral mimicry. Curr. Biol. 2021, 31, 1962–1969. [Google Scholar] [CrossRef]
- Hanks, L.M.; Millar, J.G. Field bioassays of cerambycid pheromones reveal widespread parsimony of pheromone structures, enhancement by host plant volatiles, and antagonism by components from heterospecifics. Chemoecology 2013, 23, 21–44. [Google Scholar] [CrossRef]
- Hanks, L.M.; Millar, J.G. Sex and aggregation-sex pheromones of cerambycid beetles: Basic science and practical applications. J. Chem. Ecol. 2016, 42, 631–654. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Reagel, P.F.; Wong, J.C.H.; Meier, L.R.; Silva, W.D.; Mongold-Diers, J.; Millar, J.G.; Hanks, L.M. Cerambycid beetle species with similar pheromones are segregated by phenology and minor pheromone components. J. Chem. Ecol. 2015, 41, 431–441. [Google Scholar] [CrossRef]
- Diesel, N.M.; Zou, Y.; Johnson, T.D.; Diesel, D.A.; Millar, J.G.; Mongold-Diers, J.A.; Hanks, L.M. The rare North American cerambycid beetle Dryobius sexnotatus shares a novel pyrrole pheromone component with species in Asia and South America. J. Chem. Ecol. 2017, 43, 739–744. [Google Scholar] [CrossRef]
- Yasui, H.; Fujiwara-Tsujii, N.; Yasuda, T.; Fukaya, M.; Kiriyama, S.; Nakano, A.; Watanabe, T.; Mori, K. Electroantennographic responses and field attraction of an emerging invader, the red-necked longicorn beetle Aromia bungii (Coleoptera: Cerambycidae), to the chiral and racemic forms of its male-produced aggregation-sex pheromone. Appl. Entomol. Zool. 2019, 54, 109–114. [Google Scholar] [CrossRef]
- Fonseca, M.G.; Vidal, D.M.; Zarbin, P.H. Male-produced sex pheromone of the cerambycid beetle Hedypathes betulinus: Chemical identification and biological activity. J. Chem. Ecol. 2010, 36, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Amorós, M.E.; Lagarde, L.; Carmo, H.D.; Heguaburu, V.; Monné, M.; Buenahora, J.; González, A. Trapping of Retrachydes thoracicus thoracicus (Olivier) and other neotropical cerambycid beetles in pheromone- and kairomone-baited traps. Neotrop. Entomol. 2022, 51, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Aguirre Gil, O.J.; Paredes-Espinosa, R.; Briones, R.A.; Alarcon, I.M.; Vejarano, T.E.G.; Monné, M.L.; Gonzalez, A.; Allison, J. Screening known Cerambycidae pheromones for activity with the Peruvian fauna. Agric. For. Entomol. 2021, 23, 506–511. [Google Scholar] [CrossRef]
- Curkovic, T.; Ferrera, C. Female calling and male flight orientation and searching behaviors in Callisphyris apicicornis: Evidence for a female-produced sex attractant pheromone. Int. J. Agric. Nat. Resour. 2012, 39, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Curkovic, T.; Rodríguez, D.; Huerta, A.; Bergmann, J.; Ceballos, R. Behavioral and physiological response of male Callisphyris apicicornis (Coleoptera: Cerambycidae) to virgin conspecific females’ extracts. Chil. J. Agric. Res. 2018, 78, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Barriga, J.E. Coleoptera Neotropical. 2022. Available online: http://coleoptera-neotropical.org/paginaprincipalhome.html (accessed on 30 September 2022).
- Hayes, R.A.; Griffiths, M.W.; Nahrung, H.F.; Arnold, P.A.; Hanks, L.M.; Millar, J.G. Optimizing generic cerambycid pheromone lures for Australian biosecurity and biodiversity monitoring. J. Econ. Entomol. 2016, 109, 1741–1749. [Google Scholar] [CrossRef]
- Brodie, B.S.; Popescu, V.D.; Iosif, R.; Ciocanea, C.; Manolache, S.; Vanau, G.; Gavrilidis, A.A.; Serafim, R.; Rozylowicz, L. Non-lethal monitoring of longicorn beetle communities using generic pheromone lures and occupancy models. Ecol. Indic. 2019, 101, 330–340. [Google Scholar] [CrossRef]
- Wickham, J.D.; Harrison, R.D.; Lu, W.; Chen, Y.; Hanks, L.M.; Millar, J.G. Rapid assessment of cerambycid beetle biodiversity in a tropical rainforest in Yunnan province, China, using a multicomponent pheromone lure. Insects 2021, 12, 277. [Google Scholar] [CrossRef]
- Rassati, D.; Marchioro, M.; Flaherty, L.; Poloni, R.; Edwards, S.; Faccoli, M.; Sweeney, J. Response of native and exotic longhorn beetles to common pheromone components provides partial support for the pheromone-free space hypothesis. Insect Sci. 2021, 28, 793–810. [Google Scholar] [CrossRef]
- Millar, J.G.; Richards, A.B.; Halloran, S.; Zou, Y.; Boyd, E.A.; Quigley, K.N.; Hanks, L.M. Pheromone identification by proxy: Identification of aggregation-sex pheromones of North American cerambycid beetles as a strategy to identify pheromones of invasive Asian congeners. J. Pest Sci. 2019, 92, 213–220. [Google Scholar] [CrossRef]
- Tapia, A.; Zuñiga, A.A.; Rebolledo, R. Carábidos (Coleoptera: Cerabidae) presentes en un relicto vegetacional del llano central de la Araucanía. Idesia 2011, 29, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Morrone, J.J. Biogeographic areas and transition zones of Latin America and the Caribbean islands based on panbiogeographic and cladistic analyses of the entomofauna. Ann. Rev. Entomol. 2006, 51, 467–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, J.G.; Mitchell, R.F.; Mongold-Diers, J.A.; Zou, Y.; Bográn, C.E.; Fierke, M.K.; Ginzel, M.D.; Johnson, C.W.; Meeker, J.R.; Poland, T.M.; et al. Identifying possible pheromones of cerambycid beetles by field testing known pheromone components in four widely separated regions of the United States. J. Econ. Entomol. 2018, 111, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.D.; Millar, J.G.; Hanks, L.M.; Costa, C.M.; Leite, M.O.G.; Tonelli, M.; Bento, J.M.S. Interspecific cross-attraction between the South American cerambycid beetle Cotyclytus curvatus and Megacyllene acuita is averted by minor pheromone components. J. Chem. Ecol. 2018, 44, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Graham, E.E.; Mitchell, R.F.; Reagel, P.F.; Barbour, J.D.; Millar, J.G.; Hanks, L.M. Treating panel traps with a fluoropolymer enhances their efficiency in capturing cerambycid beetles. J. Econ. Entomol. 2010, 103, 641–647. [Google Scholar] [CrossRef]
- Pajares, J.A.; Álvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef]
- Silk, P.J.; Sweeney, J.; Wu, J.; Price, J.; Gutowski, J.M.; Kettela, E.G. Evidence for a male-produced pheromone in Tetropium fuscum (F.) and Tetropium cinnamopterum (Kirby) (Coleoptera: Cerambycidae). Naturwissenschaften 2007, 94, 697–701. [Google Scholar] [CrossRef]
- Hanks, L.M.; Millar, J.G.; Moreira, J.A.; Barbour, J.D.; Lacey, E.S.; McElfresh, J.S.; Reuter, F.R.; Ray, A.M. Using generic pheromone lures to expedite identification of aggregation pheromones for the cerambycid beetles Xylotrechus nauticus, Phymatodes lecontei, and Neoclytus modestus modestus. J. Chem. Ecol. 2007, 33, 889–907. [Google Scholar] [CrossRef]
- Lacey, E.S.; Ginzel, M.D.; Millar, J.G.; Hanks, L.M. Male-produced aggregation pheromone of the cerambycid beetle Neoclytus acuminatus acuminatus. J. Chem. Ecol. 2004, 30, 1493–1507. [Google Scholar] [CrossRef]
- Miller, D.R.; Crowe, C.M.; Mayo, P.D.; Reid, L.S.; Silk, P.J.; Sweeney, J.D. Interactions between ethanol, syn-2, 3-hexanediol, 3-hydroxyhexan-2-one, and 3-hydroxyoctan-2-one lures on trap catches of hardwood longhorn beetles in southeastern United States. J. Econ. Entomol. 2017, 110, 2119–2128. [Google Scholar] [CrossRef] [PubMed]
- DMC (Dirección Meteorológica de Chile). Anuario Metereológico 2019; DMC (Dirección Meteorológica de Chile): Santiago, Chile, 2020; 131p. [Google Scholar]
- Friedmann, M. The use of ranks to avoid the assumptions of normality implicit in the analysis of variance. J. Am. Stat. Assoc. 1937, 32, 675–701. [Google Scholar] [CrossRef]
- Conover, W.J. Practical Nonparametric Statistics; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 1999. [Google Scholar]
- Monné, M.A. Catalogue of the Cerambycidae (Coleoptera) of the Neotropical Region. Part I. Subfamily Cerambycinae. Zootaxa 2005, 946, 1–765. [Google Scholar] [CrossRef]
- Monné, M.A. Catalogue of the Cerambycidae (Coleoptera) of the Neotropical Region. Part II. Subfamily Lamiinae. Zootaxa 2005, 1023, 1–759. [Google Scholar] [CrossRef]
- Monné, M.A. Catalogue of the type-species of the genera of the Cerambycidae, Disteniidae, Oxypeltidae and Vesperidae (Coleoptera) of the Neotropical Region. Zootaxa 2012, 3213, 1–183. [Google Scholar] [CrossRef]
- Fierke, M.K.; Skabeikis, D.D.; Millar, J.G.; Teale, S.A.; McElfresh, J.S.; Hanks, L.M. Identification of a male-produced aggregation pheromone for Monochamus scutellatus scutellatus and an attractant for the congener Monochamus notatus (Coleoptera: Cerambycidae). J. Econ. Entomol. 2012, 105, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Nakamuta, K.; Leal, W.S.; Nakashima, T.; Tokoro, M.; Ono, M.; Nakanishi, M. Increase of trap catches by a combination of male sex pheromones and floral attractant in longhorn beetle, Anaglyptus subfasciatus. J. Chem. Ecol. 1997, 23, 1635–1640. [Google Scholar] [CrossRef]
- Bobadoye, B.; Torto, B.; Fombong, A.; Zou, Y.; Adlbauer, K.; Hanks, L.M.; Millar, J.G. Evidence of aggregation-sex pheromone use by longhorned beetle (Coleoptera: Cerambycidae) species native to Africa. Environ. Entomol. 2019, 48, 189–192. [Google Scholar] [CrossRef]
- Botero, J.P.; Monné, M.L. Cladistic analysis of the tribe Eburiini Blanchard, 1945 and revalidation of the tribe Dychophyiini Gistel, 1848 (Coleoptera: Cerambycidae). Zool. J. Linn. Soc. 2018, 20, 1095–1122. [Google Scholar] [CrossRef]
- Sweeney, J.D.; Silk, P.J.; Gutowski, J.M.; Wu, J.; Lemay, M.A.; Mayo, P.D.; Magee, D.I. Effect of chirality, release rate, and host volatiles on response of Tetropium fuscum (F.), Tetropium cinnamopterum Kirby, and Tetropium castaneum (L.) to the aggregation pheromone, fuscumol. J. Chem. Ecol. 2010, 36, 1309–1321. [Google Scholar] [CrossRef]
- Ray, A.M.; Barbour, J.D.; McElfresh, J.S.; Moreira, J.A.; Swift, I.; Wright, I.M.; Žunič, A.; Mitchell, R.F.; Graham, E.E.; Alten, R.L.; et al. 3-Hexanediols as sex attractants and a female-produced sex pheromone for cerambycid beetles in the prionine genus Tragosoma. J. Chem. Ecol. 2012, 38, 1151–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, A.M.; Millar, J.G.; Moreira, J.A.; McElfresh, J.S.; Mitchell, R.F.; Barbour, J.D.; Hanks, L.M. North American species of cerambycid beetles in the genus Neoclytus share a common hydroxyhexanone-hexanediol pheromone structural motif. J. Econ. Entomol. 2015, 108, 1860–1868. [Google Scholar] [CrossRef] [PubMed]
- Molander, M.A.; Winde, I.B.; Burman, J.; Nyabuga, F.N.; Lindblom, T.U.T.; Hanks, L.M.; Millar, J.G.; Larsson, M.C. Common cerambycid pheromone components as attractants for longhorn beetles (Cerambycidae) breeding in ephemeral oak substrates in northern Europe. J. Chem. Ecol. 2019, 45, 537–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrone, J.J. Biogeographical regions under track and cladistic scrutiny. J. Biogeogr. 2002, 29, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Elgueta, M. Coleoptera de Chile. In Hacia un Proyecto Ciencia y Tecnología Para el Desarrollo (CYTED) Para el Inventario y Estimación de la Diversidad Entomológica en Iberoamérica; PRIBES-2000; Martín-Piera, F., Morrone, J., Melic, A., Eds.; Sociedad Entomológica Aragonesa: Zaragoza, Spain, 2000; pp. 145–154. [Google Scholar]
- Linsley, E.G. Ecology of Cerambycidae. Ann. Rev. Entomol. 1959, 4, 99–138. [Google Scholar] [CrossRef]
- Krahmer, E. Feromona de una especie de Cerambycidae atrae a los machos de otro género (Coleoptera). Rev. Chil. Entomol. 1990, 18, 95. [Google Scholar]
- Hanks, L.M.; Mongold-Diers, J.A.; Mitchell, R.F.; Zou, Y.; Wong, J.C.H.; Meier, L.R.; Johnson, T.D.; Millar, J.G. The role of minor pheromone components in segregating 14 species of longhorned beetles (Coleoptera: Cerambycidae) of the subfamily Cerambycinae. J. Econ. Entomol. 2019, 112, 2236–2252. [Google Scholar] [CrossRef]
- Artigas, J.N. Familia Cerambycidae. In Entomología Económica: Insectos de Interés Agrícola, Forestal, Médico y Veterinario, 1st ed.; Ediciones Universidad de Concepción: Concepción, Chile, 1994; Chapter 27.6; pp. 120–160. [Google Scholar]
- Ide, S. Barrenadores del floema. In Entomología Forestal en Chile; Lanfranco, D., Ruiz, C., Eds.; Ediciones Universidad Austral de Chile: Valdivia, Chile, 2010; pp. 141–161. [Google Scholar]
- Barriga, J.E.; Curkovic, T.; Fichet, T.; Henríquez, J.; Macaya, J. Nuevos antecedentes de coleopteros xilófagos y plantas hospederas en Chile, con una recopilación de citas previas. Rev. Chil. Entomol. 1993, 20, 65–91. [Google Scholar]
- Pizarro, J.; Alfaro, F.M.; Muñoz, R.A.; Barriga, J.E.; Letelier, L. Arthropods of forestry and medical-veterinary importance in the Limarí basin (Coquimbo region, Chile). Int. J. Agric. Nat. Resour. 2019, 46, 40–49. [Google Scholar] [CrossRef]
- Huerta, A.; Navarrete, W.; Aaya, J.E.; Muñoz, F. Composición y clasificación de daño de insectos en Nothofagus glauca (Fagaceae) en la Región del Maule, Chile. Rev. Colomb. Entomol. 2011, 37, 56–61. [Google Scholar] [CrossRef]
- Wong, J.C.H.; Mitchell, R.F.; Striman, B.; Millar, J.G.; Hanks, L.M. Blending synthetic pheromones of cerambycid beetles to develop trap lures that simultaneously attract multiple species. J. Econ. Entomol. 2012, 105, 906–915. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Treatments | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3-OH-6-2-Kt | R*S*- Diol | R*R*- Diol | Monocha- mol | Fuscumol | Fuscumol Acetate | Control | T2 | p | ||
| E. laetus | 0.35 ± 0.29 | 0.50 ± 0.50 | 0.10 ± 0.10 | 0.05 ± 0.05 | 0 | 0.10 ± 0.10 | 0.05 ± 0.05 | 23 | ||
| C. submetallicum | 0.80 ± 0.22 a 1 | 0 b | 0 b | 0 b | 0 b | 0 b | 0 b | 1 × 1030 | <0.0001 | 16 |
| H. corralensis | 0 | 0 | 0 | 0.05 ± 0.05 | 0 | 0 | 0 | 1 | ||
| Species | Treatments | Total | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3-OH-6-2-Kt | R*S*-diol | R*R*-diol | Monochamol | Fuscumol | Fuscumol acetate | 2-Methyl | Geranyl | Control | T2 | p | ||
| Butanol | Acetone | |||||||||||
| E. laetus | 0.625 ± 0.358 a 1 | 0.304 ± 0.241 abc | 0.220 ± 0.167 abcd | 0.161 ± 0.109 cde | 0.089 ± 0.063 defg | 0.089 ± 0.031 bcde | 0.012 ± 0.006 g | 0.029 ± 0.012 efg | 0.071 ± 0.063 fg | 7.3 | 0.0004 | 269 |
| C. submetallicum | 1.286 ± 0.537 a | 0 b | 0 b | 0 b | 0 b | 0.006 ± 0.006 b | 0.006 ± 0.006 b | 0 b | 0 b | 7.8 | 0.0003 | 218 |
| C. testaceus | 0.024 ± 0.006 | 0.024 ± 0.006 | 0 | 0.024 ± 0.006 | 0.029 ± 0.029 | 0.012 ± 0.012 | 0 | 0 | 0.006 ± 0.006 | 2.4 | 0.0645 | 20 |
| H. corralensis | 0.006 ± 0.006 | 0.012 ± 0.012 | 0 | 0 | 0.006 ± 0.006 | 0.012 ± 0.012 | 0 | 0.029 ± 0.029 | 0 | 11 | ||
| X. flavonitida | 0.006 ± 0.006 | 0.006 ± 0.006 | 0.006 ± 0.006 | 0 | 0.012 ± 0.012 | 0.006 ± 0.006 | 0 | 0.012 ± 0.012 | 0 | 8 | ||
| A. cristatipennis | 0 | 0 | 0 | 0 | 0.006 ± 0.006 | 0.006 ± 0.006 | 0 | 0.006 ± 0.006 | 0.006 ± 0.006 | 4 | ||
| N. humeralis | 0 | 0.006 ± 0.006 | 0 | 0.006 ± 0.006 | 0 | 0 | 0 | 0 | 0.006 ± 0.006 | 3 | ||
| M. quadrimaculatus | 0 | 0.006 ± 0.006 | 0 | 0 | 0 | 0.006 ± 0.006 | 0 | 0 | 0 | 2 | ||
| C. sulcicorne | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 ± 0.006 | 0 | 0 | 1 | ||
| X. semipolita | 0 | 0 | 0 | 0 | 0 | 0.006 ± 0.006 | 0 | 0 | 0 | 1 | ||
| S. virescens | 0 | 0 | 0 | 0 | 0 | 0.006 ± 0.006 | 0 | 0 | 0 | 1 | ||
| E. laetus | C. submetallicum | C. testaceus | ||||
|---|---|---|---|---|---|---|
| Treatments | Males | Females | Males | Females | Males | Females |
| 3-OH-6-2-Kt | 0.345 ± 0.221 a 1 | 0.280 ± 0.137 a | 0.554 ± 0.206 a | 0.732 ± 0.331 a | 0.024 ± 0.010 | 0 |
| R*S*-diol | 0.089 ± 0.071 abcd | 0.214 ± 0.170 abcd | 0 b | 0 b | 0.024 ± 0.027 | 0 |
| R*R*-diol | 0.089 ± 0.071 abcd | 0.131 ± 0.096 abcde | 0 b | 0 b | 0 | 0 |
| Monochamol | 0.065 ± 0.049 cde | 0.095 ± 0.060 bcdef | 0 b | 0 b | 0.024 ± 0.027 | 0 |
| Fuscumol | 0.048 ± 0.039 defg | 0.042 ± 0.024 efg | 0 b | 0 b | 0.024 ± 0.041 | 0.006 ± 0.006 |
| Fuscumol acetate | 0.030 ± 0.012 bcd | 0.060 ± 0.021 cdef | 0.006 ± 0.006 b | 0 b | 0.024 ± 0.021 | 0 |
| 2-Methylbutanol | 0 g | 0.012 ± 0.006 g | 0.006 ± 0.006 b | 0 b | 0 | 0 |
| Geranylacetone | 0 g | 0.030 ± 0.012 defg | 0 b | 0 b | 0 | 0 |
| Control | 0.024 ± 0.024 efg | 0.048 ± 0.039 fg | 0 b | 0 b | 0 | 0.006 ± 0.006 |
| T2 | 6.89 | 4.47 | 6.2 | 1 × 1030 | 2.46 | |
| p | 0.0006 | 0.0053 | 0.001 | <0.0001 | 0.0599 | |
| Total | 116 | 153 | 95 | 123 | 18 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curkovic, T.; Arraztio, D.; Huerta, A.; Rebolledo, R.; Cheuquel, A.; Contreras, A.; Millar, J.G. Generic Pheromones Identified from Northern Hemisphere Cerambycidae (Coleoptera) Are Attractive to Native Longhorn Beetles from Central-Southern Chile. Insects 2022, 13, 1067. https://doi.org/10.3390/insects13111067
Curkovic T, Arraztio D, Huerta A, Rebolledo R, Cheuquel A, Contreras A, Millar JG. Generic Pheromones Identified from Northern Hemisphere Cerambycidae (Coleoptera) Are Attractive to Native Longhorn Beetles from Central-Southern Chile. Insects. 2022; 13(11):1067. https://doi.org/10.3390/insects13111067
Chicago/Turabian StyleCurkovic, Tomislav, Diego Arraztio, Amanda Huerta, Ramón Rebolledo, Arly Cheuquel, Américo Contreras, and Jocelyn G. Millar. 2022. "Generic Pheromones Identified from Northern Hemisphere Cerambycidae (Coleoptera) Are Attractive to Native Longhorn Beetles from Central-Southern Chile" Insects 13, no. 11: 1067. https://doi.org/10.3390/insects13111067