
Predicting Distribution of the Asian Longhorned Beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Natural Enemies in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Distribution Point Collection
2.2. Climate Data Collection and Environmental Variables
2.3. Optimization of Model Parameters
2.4. Species Distribution Model
2.5. Change of Suitable Area under Different Climates
3. Results
3.1. Reliability Analysis of Models Established for A. glabripennis, D. helophoroides, and D. major by Using MaxEnt
3.2. Potential Suitable Areas for A. glabripennis, D. helophoroides, and D. major under Current and Future Climate Conditions
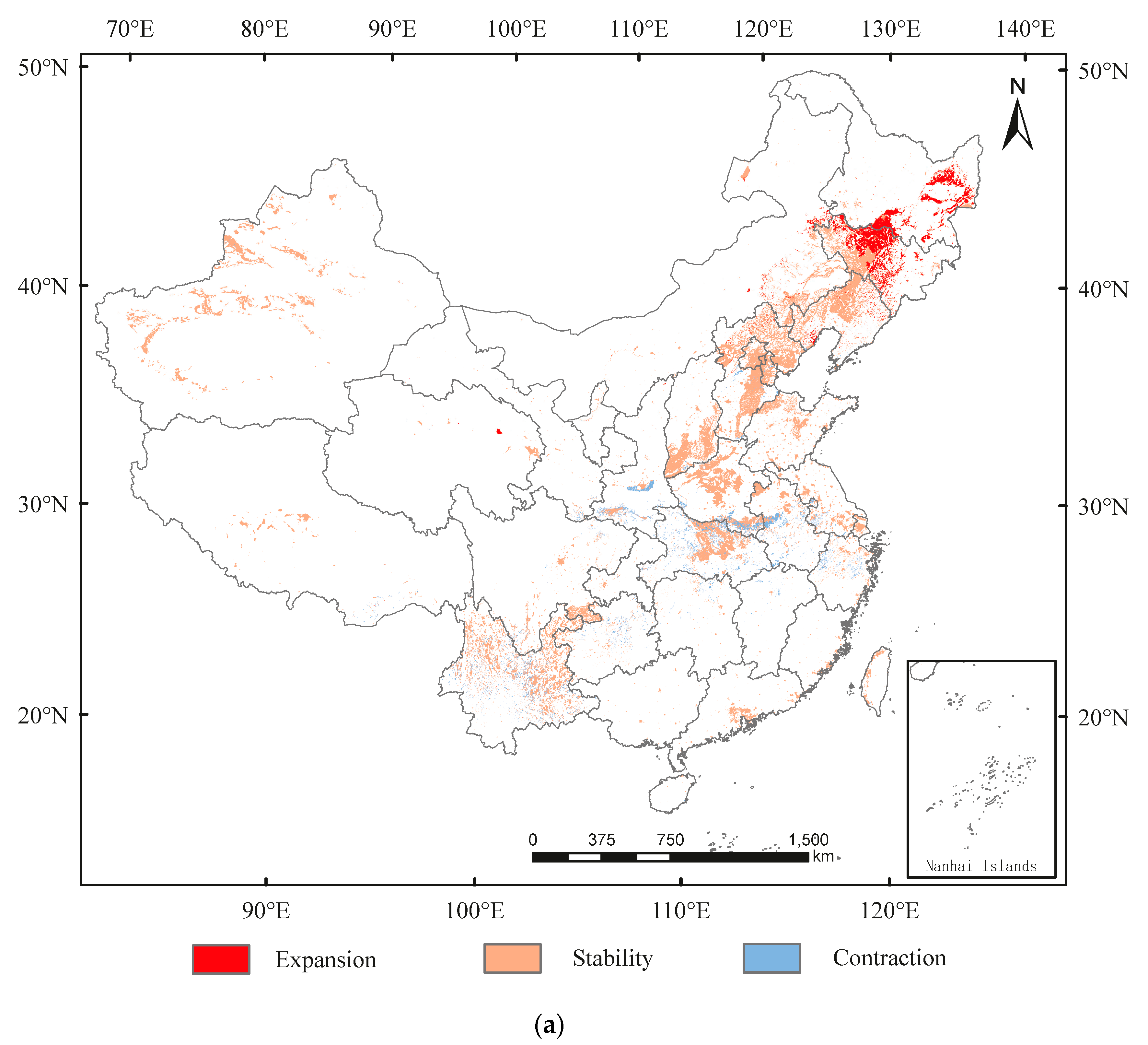
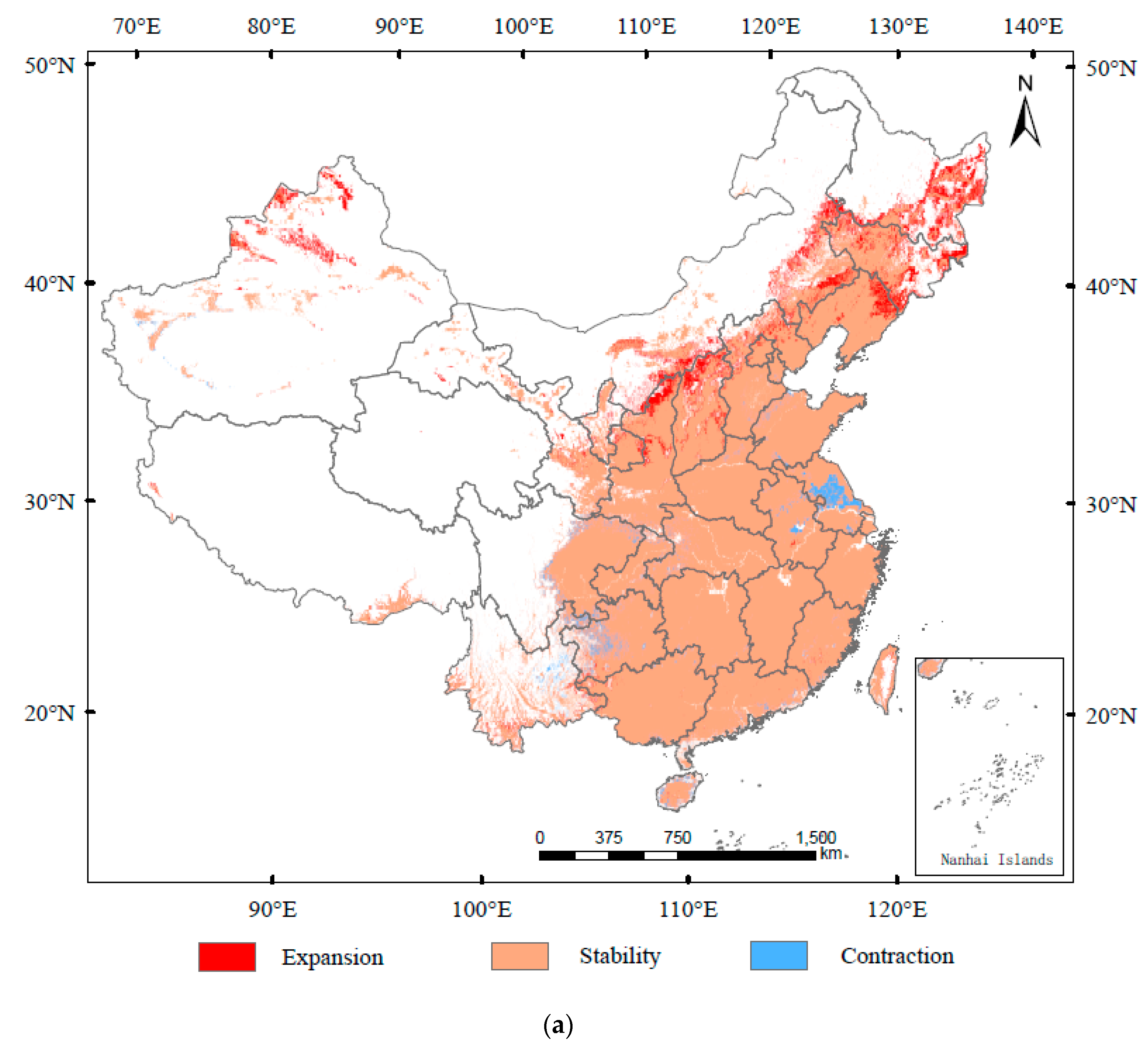
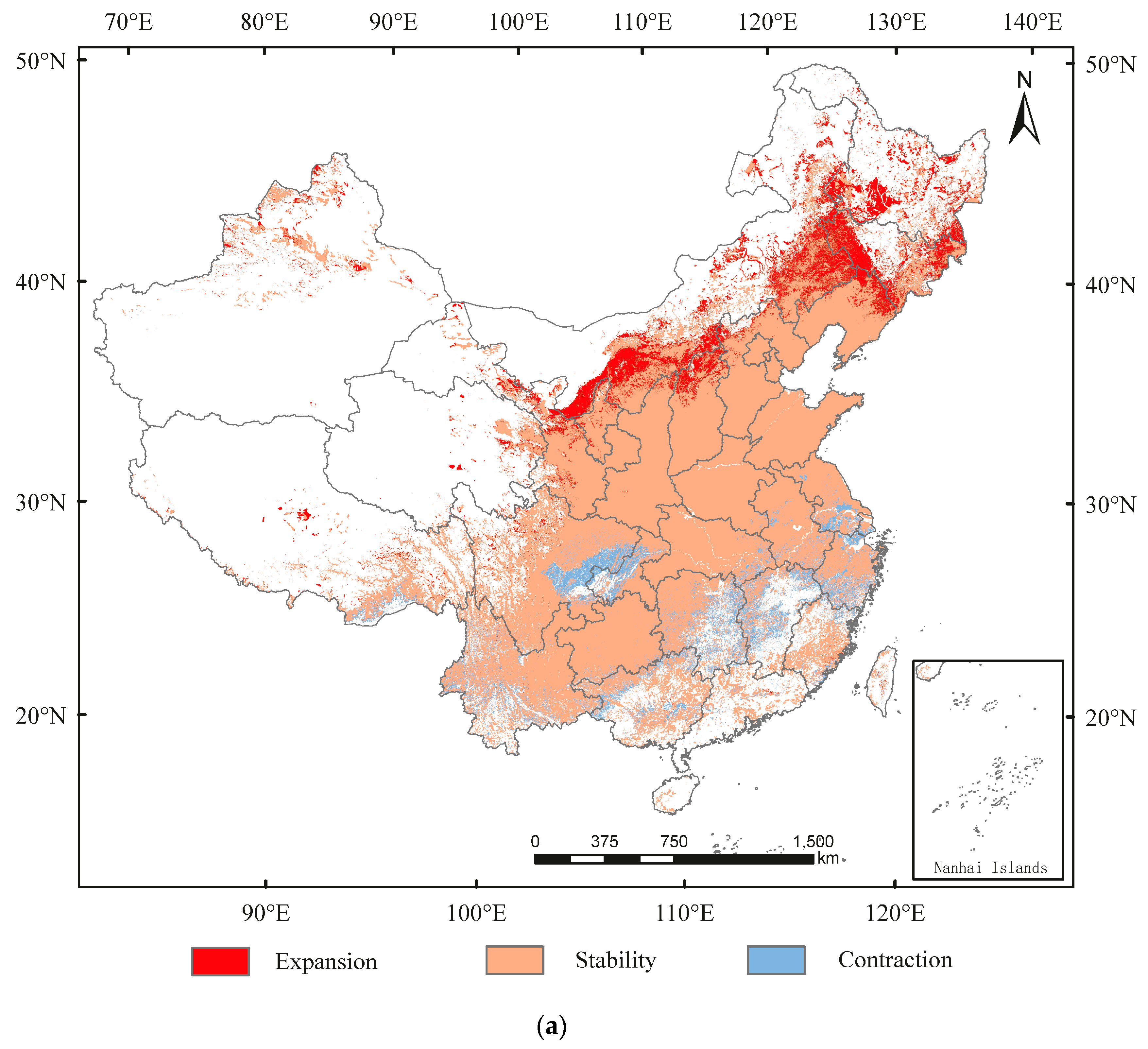
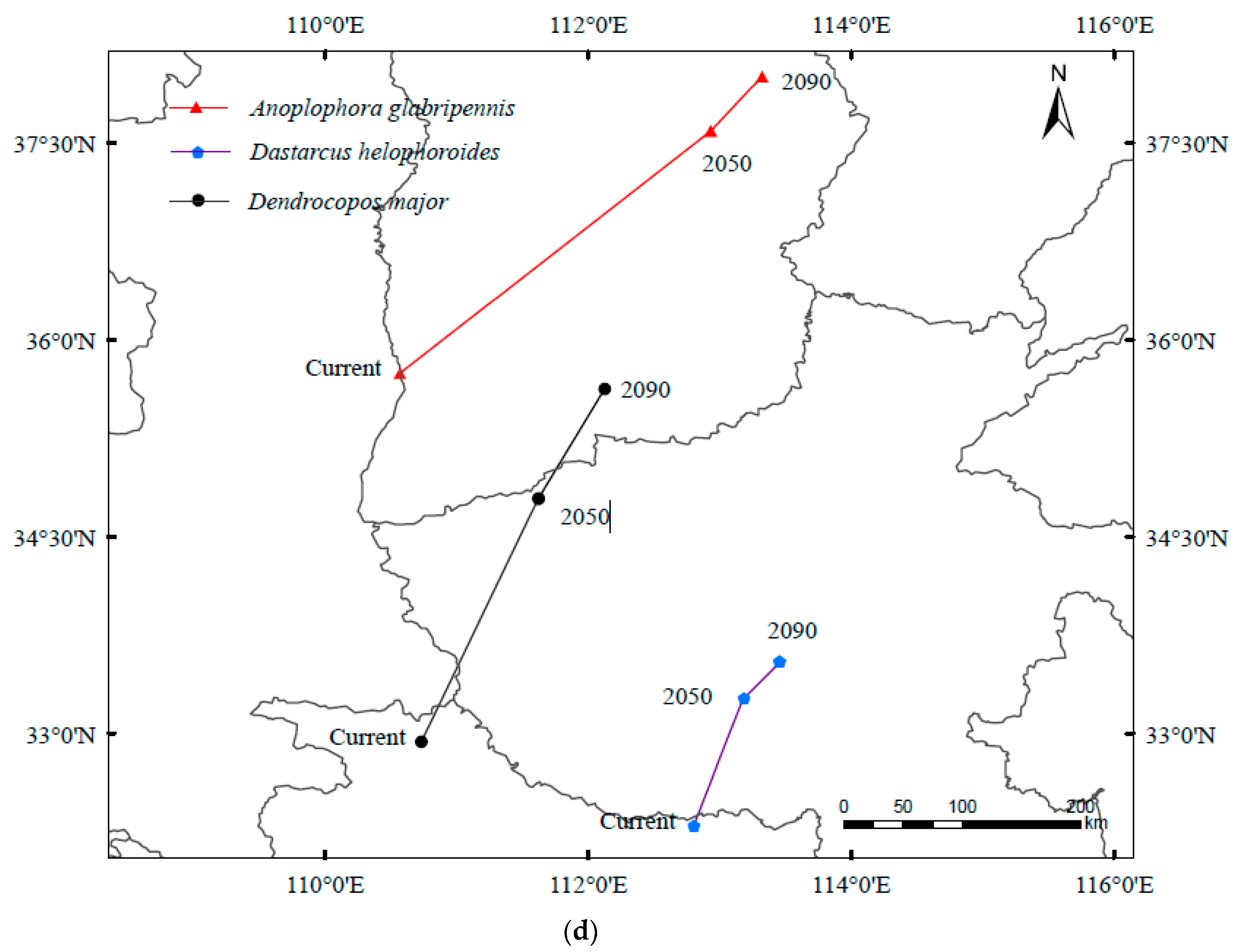
3.3. Spatial Transfer Characteristics of Suitable Areas for A. glabripennis, D. helophoroides, and D. major
3.4. The Distribution Overlap Region of A. glabripennis, D. helophoroides, and D. major
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haack, R.A.; Hérard, F.; Sun, J.H.; Turgeon, J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 2010, 55, 521–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingafelter, S.W.; Hoebeke, E.R. Variation and homology in elytral maculation in the Anoplophora malasiaca/macularia species complex (Coleoptera: Cerambycidae) of Japan and Taiwan. Proc. Entomol. Soc. Wash. 2001, 103, 757–769. [Google Scholar]
- Van der Gaag, D.; Loomans, A. Host plants of Anoplophora glabripennis, a review. EPPO Bull. 2014, 44, 518–528. [Google Scholar] [CrossRef]
- Haack, R.A.; Cavey, J.F.; Hoebeke, E.R.; Law, K. Anoplophora glabripennis: A new tree-infesting exotic cerambycid invades New York. Newsl. Mich. Entomol. Soc. 1996, 41, 1–3. [Google Scholar]
- Hérard, F.; Ciampitti, M.; Maspero, M.; Krehan, H.; Benker, U.; Boegel, C.; Schrage, R.; Bouhot-Delduc, L.; Bialooki, P. Anoplophora species in Europe: Infestations and management processes. EPPO Bull. 2006, 36, 470–474. [Google Scholar] [CrossRef]
- Turgeon, J.J.; Orr, M.; Grant, C.; Wu, Y.; Gasman, B. Decade-old satellite infestation of Anoplophora glabripennis Motschulsky (Coleoptera: Cerambycidae) found in Ontario, Canada outside regulated area of founder population. Coleopt. Bull. 2015, 69, 674–678. [Google Scholar] [CrossRef]
- Javal, M.; Lombaert, E.; Tsykun, T.; Courtin, C.; Kerdelhué, C.; Prospero, S.; Roques, A.; Roux, G. Deciphering the worldwide invasion of the Asian long-horned beetle: A recurrent invasion process from the native area together with a bridgehead effect. Mol. Ecol. 2019, 28, 951–967. [Google Scholar] [CrossRef]
- Yue, C.Y.; Zhang, X.P.; Liu, A.H.; Alimu, M. Risk analysis of the occurrence of Anoplophora glabripennis in Xinjiang. J. Northwest For. Univ. 2011, 26, 153–156. [Google Scholar]
- Thomson, L.J.; Macfadyen, S.; Hoffmann, A.A. Predicting the effects of climate change on natural enemies of agricultural pests. Biol. Control 2010, 52, 296–306. [Google Scholar] [CrossRef]
- Hu, J.; Angeli, S.; Schuetz, S.; Luo, Y.; Hajek, A.E. Ecology and management of exotic and endemic Asian longhorned beetle Anoplophora glabripennis. Agric. For. Entomol. 2009, 11, 359–375. [Google Scholar] [CrossRef]
- Marchioro, M.; Faccoli, M. Successful eradication of the Asian longhorn beetle, Anoplophora glabripennis, from north-eastern Italy: Protocol, techniques and results. Insects 2021, 12, 877. [Google Scholar] [CrossRef]
- Caltagirone, L.E. Landmark examples in classical biological control. Annu. Rev. Entomol. 1981, 26, 213–232. [Google Scholar] [CrossRef]
- Hufbauer, R.A.; Roderick, G.K. Microevolution in biological control: Mechanisms, patterns, and processes. Biol. Control 2005, 35, 227–239. [Google Scholar] [CrossRef]
- Luo, L.P.; Wang, X.Y.; Yang, Z.Q.; Zhao, J.X.; Tang, Y.L. Progress in biological control over Anoplophora glabripennis (Coleoptera: Cerambycidae). Biol. Dis. Sci. 2018, 41, 247–255. [Google Scholar]
- Yang, Z.Q.; Wang, X.Y.; Zhang, Y.N. Recent advances in biological control of important native and invasive forest pests in China. Biol. Control 2014, 68, 117–128. [Google Scholar] [CrossRef]
- Demidko, D.A.; Demidko, N.N.; Mikhaylov, P.V.; Sultson, S.M. Biological strategies of invasive bark beetles and borers species. Insects 2021, 12, 367. [Google Scholar] [CrossRef]
- Jiao, Z.B.; Wan, T.; Wen, J.B.; Hu, J.F.; Luo, Y.Q.; Zhang, L.S.; Fu, L.J. Functional response and numerical response of great spotted woodpecker Dendrocopos major on Asian longhorned beetle Anoplophora glabripennis larvae. Acta Zool. Sin. 2008, 54, 1106–1111. [Google Scholar]
- Yue, C.Y.; Zhang, X.P.; Zhang, J.W.; Jiao, S.P.; Nuergu; Kereman; Han, X.W.; Ji, L.L. A preliminary study on the effect of release of Dastarcus helophoroides to control Anoplophora glabripennis in forest land of Yanqi Basin. Xinjiang Agric. Sci. 2013, 50, 2085–2091. [Google Scholar]
- Wan, T.; Jiao, Z.B.; Wen, J.B.; Hu, J.F.; Luo, Y.Q.; Fu, L.J.; Zhang, L.S. Selective predation by the great spotted woodpecker Dendrocopos major on the Asian longhorned beetle Anoplophora glabripennis in winter. Acta Zool. Sin. 2008, 54, 555–560. [Google Scholar]
- Lv, F.; Hai, X.X.; Wang, Z.G.; Liu, B.X.; Yan, A.H.; Bi, Y.G. Research progress in Dastarcus helophoroides Fairmaire (Bothrideridae), an important natural enemy of the longhorn beetle pests. Sci. Sericul. 2014, 40, 1107–1113. [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [Green Version]
- MacLeod, A.; Evans, H.F.; Baker, R.H.A. An analysis of pest risk from an Asian longhorn beetle (Anoplophora glabripennis) to hardwood trees in the European community. Crop Prot. 2002, 21, 635–645. [Google Scholar] [CrossRef]
- Peterson, A.T.; Scachetti-Pereira, R. Potential geographic distribution of Anoplophora glabripennis (Coleoptera: Cerambycidae) in North America. Am. Midl. Nat. 2004, 151, 170–178. [Google Scholar] [CrossRef]
- Shatz, A.J.; Rogan, J.; Sangermano, F.; Miller, J.; Elmes, A. Modeling the risk of spread and establishment for Asian longhorned beetle (Anoplophora glabripennis) in Massachusetts from 2008–2009. Geocarto Int. 2016, 31, 813–831. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Yemshanov, D.; Hope, E.S. Potential economic impacts of the Asian longhorned beetle (Coleoptera: Cerambycidae) in eastern Canada. J. Econ. Entomol. 2020, 113, 839–850. [Google Scholar] [CrossRef]
- Byeon, D.; Kim, S.; Jung, J.; Jung, S.; Kim, K.; Lee, W. Climate-based ensemble modelling to evaluate the global distribution of Anoplophora glabripennis (Motschulsky). Agric. For. Entomol. 2021, 23, 569–583. [Google Scholar] [CrossRef]
- Zhou, Y.T.; Ge, X.Z.; Zou, Y.; Guo, S.W.; Wang, T.; Zong, S.X. Prediction of the potential global distribution of the Asian longhorned beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) under climate change. Agric. For. Entomol. 2021, 23, 557–568. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.W.; Lu, Y.X.; Fang, Y.J.; Xin, X.G.; Li, L.; Li, W.P.; Jie, W.H.; Zhang, J.; Liu, Y.M.; Zhang, L.; et al. The Beijing climate center climate system model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Li, D.X.; Li, Z.X.; Liu, Z.W.; Yang, Y.J.; Khoso, A.G.; Wang, L.; Liu, D. Climate change simulations revealed potentially drastic shifts in insect community structure and crop yields in China’s farmland. J. Pest Sci. 2022. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudik, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [Green Version]
- Aidoo, O.F.; Souza, P.G.C.; da Silva, R.S.; Santana, J.P.A.; Picanço, M.C.; Kyerematen, R.; Sètamou, M.; Ekesi, S.; Borgemeister, C. Climate-induced range shifts of invasive species (Diaphorina citri Kuwayama). Pest Manag. Sci. 2022, 78, 2534–2549. [Google Scholar] [CrossRef]
- Lemke, P.; Ren, J.F.; Alley, R.B.; Allison, I.; Carrasco, J.F.; Flato, G.; Fujii, Y.; Kaser, G.; Mote, P.; Thomas, R.H.; et al. IPCC: Contribution of Working Groups I, II, III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Wang, C.; Hawthorne, D.; Qin, Y.J.; Pan, X.B.; Li, Z.H.; Zhu, S.F. Impact of climate and host availability on future distribution of Colorado potato beetle. Sci. Rep. 2017, 7, 4489. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.P.; Li, H.Q.; Zhao, L. Incorporating anthropogenic variables into ecological niche modeling to predict areas of invasion of Popillia japonica. J. Pest Sci. 2017, 90, 151–160. [Google Scholar] [CrossRef]
- Lecheta, M.C.; Corrêa, R.C.; Moura, M.O. Climate shapes the geographic distribution of the blowfly Sarconesia chlorogaster (Diptera: Calliphoridae): An environmental niche modeling approach. Environ. Entomol. 2017, 46, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.Z.; Zong, S.X.; He, S.Y.; Liu, Y.T.; Kong, X.Q. Areas of China predicted to have a suitable climate for Anoplophora chinensis under a climate-warming scenario. Entomol. Exp. Appl. 2014, 153, 256–265. [Google Scholar] [CrossRef]
- Li, Z.H.; Qin, Y.J. Review on the quantitative assessment models for pest risk analysis and their comparison. Plant Prot. 2018, 44, 134–145. [Google Scholar]
- Jimenez-Valverd, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discriminationmeasure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distribut. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial bias in the GBIF database and its effect on modeling species’ geographicdistributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- Hickling, R.; David, B.R.; Jane, K.H.; Chris, D.T. A northward shift of range margins in British Odonata. Glob. Chang. Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Speight, M.R.; Hunter, M.D.; Watt, A.D. Ecology of Insects: Concepts and Applications; Blackwell Science Ltd.: Oxford, UK, 1999. [Google Scholar]
- Broadmeadow, M. Climate Change: Impacts on UK Forests; Bulletin 125; Forestry Commission: Edinburgh, UK, 2002. [Google Scholar]
- Morimoto, N.; Imura, O.; Kiura, T. Potential effects of global warming on the occurrence of Japanese pest insects. Appl. Entomol. Zool. 1998, 33, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Umina, P.A.; Weeks, A.R.; Kearney, M.R.; McKechnie, S.W.; Hoffmann, A.A. A rapid shift in a classic clinal pattern in Drosophila reflecting climate change. Science 2005, 308, 691–693. [Google Scholar] [CrossRef]
- Musolin, D.L. Insects in a warmer world: Ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change. Glob. Change Biol. 2007, 13, 1565–1585. [Google Scholar] [CrossRef]
- Stephens, A.E.A.; Kriticos, D.J.; Leriche, A. The current and future potential geographical distribution of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Bull. Entomol. Res. 2007, 97, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Ma, C.S. The impacts of extreme high temperature on insect populations under climate change: A review. Sci. Sin. Vitae 2016, 46, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.P.; Dang, Y.Q.; Wang, X.Y.; Li, F.; Yang, Z.Q. The potential for using Dastarcus helophoroides as a biological control agent against Anoplophora glabripennis in Hunchun, Jilin province. Chin. J. Appl. Entomol. 2021, 58, 187–194. [Google Scholar]
- Keena, M.A.; Moore, P.M. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) larvae and pupae. Environ. Entomol. 2010, 39, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; He, H.; Lu, C.; Liang, Y.; Wu, H.; Zheng, L.; Wang, X.; Liang, G. Comparative transcriptome analysis of two populations of Dastarcus helophoroides (Fairmaire) under high temperature stress. Forests 2022, 13, 13. [Google Scholar] [CrossRef]
- Luo, N.G.; Guo, Y.N.; Gao, Z.B.; Chen, K.X.; Chou, J.M. Assessment of CMIP6 and CMIP5 model performance for extreme temperature in China. Atmos. Ocean. Sci. Lett. 2020, 13, 589–597. [Google Scholar] [CrossRef]
- Wei, J.R.; Yang, Z.Q.; Liu, Y.L.; Zhao, H.B.; Tang, H. Distribution and ecological biology of Dastarcus helophoroides. For. Pest Dis. 2009, 28, 16–18. [Google Scholar]
- Xie, J.L. Dastarcus helophoroides against Monochamus alternatus in pine forest. Biol. Disaster Sci. 2019, 42, 66–69. [Google Scholar]
- Zhang, Z.M. Studies on control effect of Dastarcus helophoroides on Monochamus alternates. For. Prospect. Des. 2019, 39, 42–45. [Google Scholar]
- Gao, S.K.; Zahng, Y.L.; Tang, Y.L.; Yang, Z.Q.; Wang, X.Y.; Lu, J.F.; Wang, J.; Situ, C.N.; Fu, F.Y. Overwintering characteristics and cold-hardiness of biotype of Dastarcus helophoroides (Coleoptera: Bothrideridae) on Monochamus alternatus (Coleoptera: Cerambycidae). Sci. Silvae Sin. 2016, 52, 68–74. [Google Scholar]
- Huang, H.H.; Xu, Z.F.; Yang, Z.Q.; Wang, X.D.; Fan, J.X.; Qian, M.H.; Te, L.K.; Chen, J.J. Important natural enemy of the Monochamus alternatus Hope—Dastarcus helophoroides. Guangdong For. Sci. Technol. 2003, 19, 76–77, 82. [Google Scholar]
- Zheng, Y.N.; Jang, S.W.; Huang, F.Y.; Yang, Z.Q.; Gao, G.P.; Liu, P.X.; Shi, Y.; Fan, L.C.; Zhang, Y.L. Dastarcus helophoroides, natural enemy of Monochamus saltuarius was found in pine tree infected by Bursaphelenchus xylophilus in Liaoning, China. Chin. J. Biol. Control 2020, 36, 315–318. [Google Scholar]
- Hao, C.F.; Wang, H.P.; Zahng, Z.Q.; Chang, Y.; Li, M.L. Characterization and expression analysis of HSP70 gene in Dastarcus helophoroides. J. Northeast. For. Univ. 2016, 44, 108–115, 124. [Google Scholar]
- Qiu, L.F.; Zhong, L.; Shao, J.L.; Che, S.C.; Li, G. Influence of environmental temperature and adult body size on the mortality and fecundity of Dastarcus helophoroides. Chin. J. Appl. Entomol. 2021, 58, 959–965. [Google Scholar]
- Wei, J.R.; Wang, S.Y.; Niu, Y.L.; Tang, Y.L. Cold tolerance of Dastarcus helophoroides. For. Pest Dis. 2010, 29, 19–20, 46. [Google Scholar]
- Wei, K.; Zhang, Y.N.; Yang, Z.Q.; Wang, X.Y.; Han, Y.Y.; Liu, Y.; Qu, M.Q. Overwintering and cold tolerance of a parasitic natural enemy, Dastarcus helophoroides (Coleoptera: Bothrideridae), in Gansu Province. For. Res. 2015, 28, 588–592. [Google Scholar]
- Zhou, Y.J.; Lu, C.D.; Shen, H.Y.; Liang, Y.; Chen, Z.H.; Zhong, J.H.; Tan, J.J.; Wang, X.X.; Liang, G.H. Adaptation and physiological response of Dastarcus helophoroides (Fairmaire) to high temperature stress. Chin. J. Biol. Control 2021, 37, 1179–1188. [Google Scholar]
- Zhang, D.R.; Zheng, J.; Fan, J.L.; Fang, Z.C.; Ji, Q.Y.; Yuan, Y.Z.; Liu, W.F. Spatiotemporal variations of extreme temperature indices in different climatic zones of China over the past 60 years. Chin. J. Agrometeorol. 2019, 40, 422–434. [Google Scholar]
- Michalek, K.G.; Miettinen, J. Dendrocopos major great spotted woodpecker. BWP Update 2003, 5, 101–184. [Google Scholar]
- Sun, M.R.; Li, K.Q.; Zhu, J.J.; Kao, C.C.; Sun, X.J.; Zhou, Z.S. Reproduction habits of three species of woodpeckers and their prey on insects. For. Pest Dis. 2002, 21, 12–14. [Google Scholar]
- Koenig, W.D.; Walters, E.L.; Walters, J.R.; Kellam, J.S.; Michalek, K.G.; Schrader, M.S. Seasonal body weight variation in five species of woodpeckers. Condor 2005, 107, 810–822. [Google Scholar] [CrossRef]
- McNab, B.K. Energetics of toucans, abarbet, andahornbill: Implications for avian frugivory. Auk 2001, 11, 8916–8933. [Google Scholar]
- Hitch, A.T.; Leberg, P.L. Breeding distributions of North American bird species moving north as result of climate change. Conserv. Biol. 2007, 21, 534–539. [Google Scholar] [CrossRef]
- Leech, D.I.; Crick, H.Q.P. Influence climate change on the abundance, distribution phenology of woodland bird species in temperate regions. Ibis 2007, 149, 128–145. [Google Scholar] [CrossRef]
- Lu, J.F.; Cai, J.Y.; Zhan, M.K.; Fu, F.Y.; Wang, J.; Situ, C.N. Research advances on biological characteristics and application of Dastarcus helophoroides. Guizhou For. Sci. Technol. 2016, 44, 43–48. [Google Scholar]
- Hu, C.H.; Zha, Y.P.; Chen, J.Y. Research progress of controlling longhorn beetle by Dastarcus helophoroides. Hubei For. Sci. Technol. 2012, 176, 48–50. [Google Scholar]
- Gao, Y.; Xie, C.X.; Liu, Y.P.; Zheng, H.Y.; Xu, M. Study on parasitic efficiency of releasing Dastarcus helophoroides to control Anoplophora glabripennis in park willows. J. Southwest For. Univ. 2013, 33, 104–106. [Google Scholar]
- Li, F. Study on the Key Control Factors of Asian Longhorned Beetle (Anoplophora glabripennis) Natural Populations; Shenyang Agricultural University: Shenyang, China, 2017. [Google Scholar]
- Hu, J.F.; Wen, J.B.; Luo, Y.Q. Review of study on great spotted woodpecker, Picoides major. J. Anhui Agric. Univ. 2008, 35, 405–410. [Google Scholar]
- Jiao, Z.B. The Control Effect of Picoides Maior to Anoplophora Glabripennis; Beijing Forestry University: Beijing, China, 2008. [Google Scholar]
- Wang, Z.; Yuan, K.Y.; Wang, X.M. Investigation of damages and annual life history of Anoplophora glabripennis (Motsch.) in Baotou in the Inner Mongolia Autonomous region. J. Inner Mongolia Agric. Univ. 2016, 37, 87–96. [Google Scholar]
- Ma, L.L. Study on the insect monitoring and control techniques of Anoplophora glabripennis and Myzus persicae in Lanzhou Lanshan area. Gansu For. 2021, 1, 43–44. [Google Scholar]
- Ogura, N.; Tabata, K.; Wang, W. Rearing of the colydiid beetle predator, Dastarcus helophoroides, on artificial diet. BioControl 1999, 44, 291–299. [Google Scholar] [CrossRef]
- Togashi, K.; Itabashi, M. Maternal size dependency of ovariole number in Dastarcus helophoroides (Coleoptera: Colydiidae). J. For. Res. 2005, 10, 373–376. [Google Scholar] [CrossRef]
- Lim, J.; Oh, H.; Park, S.; Koh, S.; Lee, S. First record of the family Bothrideridae (Coleoptera) in Korea represented by the wood-boring beetle ectoparasite, Dastarcus helophoroides. J. Asia-Pac. Entomol. 2012, 15, 273–275. [Google Scholar] [CrossRef]
- Gould, J.R.; Aflague, B.; Murphy, T.C.; McCartin, L.; Elkinton, J.S.; Rim, K.; Duan, J.J. Collecting nontarget wood-boring insects for host-specificity testing of natural enemies of Cerambycids: A case study of Dastarcus helophoroides (Coleoptera: Bothrideridae), a parasitoid of the Asian longhorned beetle (Coleoptera: Cerambycidae). Environ. Entomol. 2018, 47, 1440–1450. [Google Scholar] [CrossRef] [Green Version]
- Rim, K.; Golec, J.R.; Duan, J.J. Host selection and potential non-target risk of Dastarcus helophoroides, a larval parasitoid of the Asian longhorned beetle, Anoplophora glabripennis. Biol. Control 2018, 123, 120–126. [Google Scholar] [CrossRef]
- Li, H.P.; Lv, F.; Bi, Y.G.; Wang, Z.G. Reviews on the serious wood-boring pest Anoplophora glabripennis in forestry of China. For. Ecol. Sci. 2020, 35, 1–9. [Google Scholar]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment Variable | Variable Type |
|---|---|
| Bio_1 | Annual mean temperature/°C |
| Bio_2 | Mean diurnal range/°C |
| Bio_3 | Isothermally [(Bio2/Bio7) × 100] |
| Bio_4 | Temperature seasonality |
| Bio_5 | Maximum temperature of the warmest month/°C |
| Bio_6 | Minimum temperature of the coldest month/°C |
| Bio_7 | Temperature annual range (Bio5–Bio6)/°C |
| Bio_8 | Mean temperature of the wettest quarter/°C |
| Bio_9 | Mean temperature of the driest quarter/°C |
| Bio_10 | Mean temperature of the warmest quarter/°C |
| Bio_11 | Mean temperature of the coldest quarter/°C |
| Bio_12 | Annual precipitation/mm |
| Bio_13 | Precipitation of the wettest period/mm |
| Bio_14 | Precipitation of the driest period/mm |
| Bio_15 | Precipitation seasonality (CV) |
| Bio_16 | Precipitation of the wettest quarter/mm |
| Bio_17 | Precipitation of the driest quarter/mm |
| Bio_18 | Precipitation of the warmest quarter/mm |
| Bio_19 | Precipitation of the coldest quarter/mm |
| Bio_20 | Aspect |
| Bio_21 | Elevation/m |
| Bio_22 | Slope |
| Bio_23 | GlobalMaps°LandCover v3° |
| Bio_24 | GlobalMaps°Vegetation v2 |
| Bio_25 | Vegetation |
| Parameter | Species | FC | RM | Training AUC | Test AUC | AICc |
|---|---|---|---|---|---|---|
| Default | Anoplophora glabripennis | LQPH | 1 | 0.9933 | 0.8962 | 237.61 |
| Dastarcus_helophoroides | 0.9669 | 0.8192 | 160.69 | |||
| Dendrocopos_major | 0.9273 | 0.8282 | 372.74 | |||
| Optimized | Anoplophora glabripennis | LQ | 0.1 | 0.9937 | 0.9179 | 0 |
| Dastarcus_helophoroides | QT | 1.8 | 0.9800 | 0.8238 | 0 | |
| Dendrocopos_major | QP | 3.8 | 0.9321 | 0.8504 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.-C.; Wang, J.-G.; Lei, Y.-H. Predicting Distribution of the Asian Longhorned Beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Natural Enemies in China. Insects 2022, 13, 687. https://doi.org/10.3390/insects13080687
Zhang Q-C, Wang J-G, Lei Y-H. Predicting Distribution of the Asian Longhorned Beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Natural Enemies in China. Insects. 2022; 13(8):687. https://doi.org/10.3390/insects13080687
Chicago/Turabian StyleZhang, Quan-Cheng, Jun-Gang Wang, and Yong-Hui Lei. 2022. "Predicting Distribution of the Asian Longhorned Beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Natural Enemies in China" Insects 13, no. 8: 687. https://doi.org/10.3390/insects13080687