
Tick Cell Culture Analysis of Growth Dynamics and Cellular Tropism of Rickettsia buchneri, an Endosymbiont of the Blacklegged Tick, Ixodes scapularis

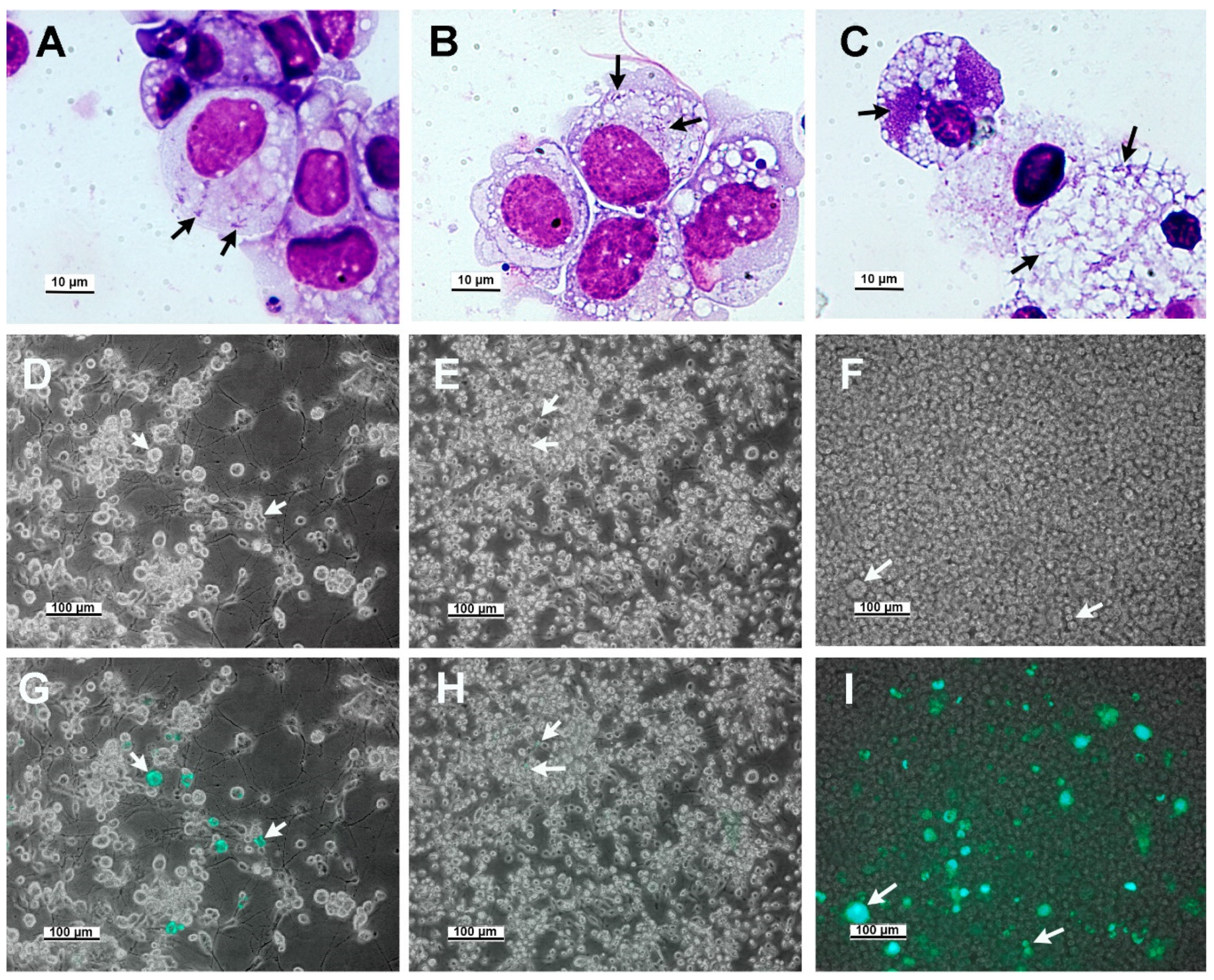{kind=link}
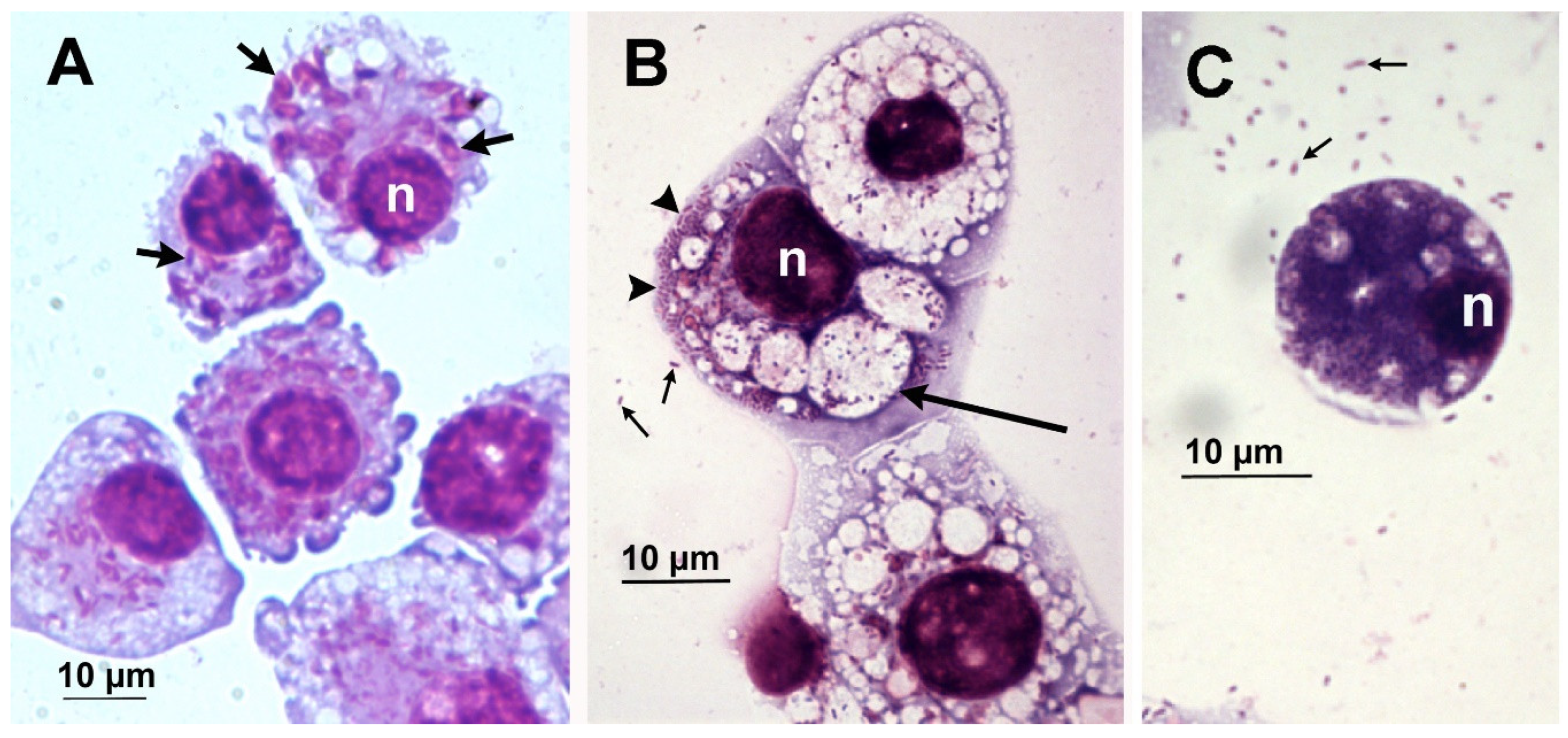{kind=link}
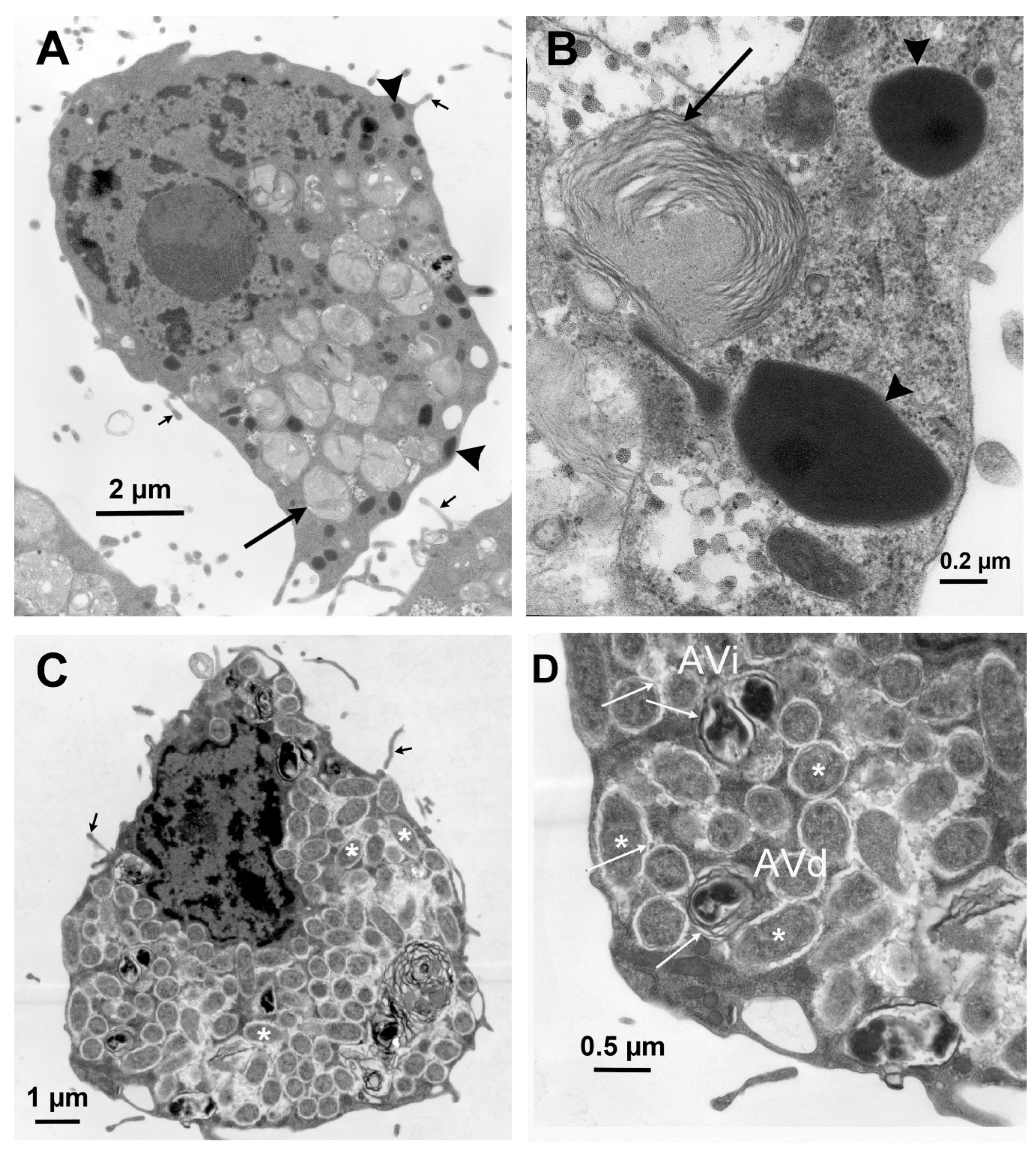{kind=link}
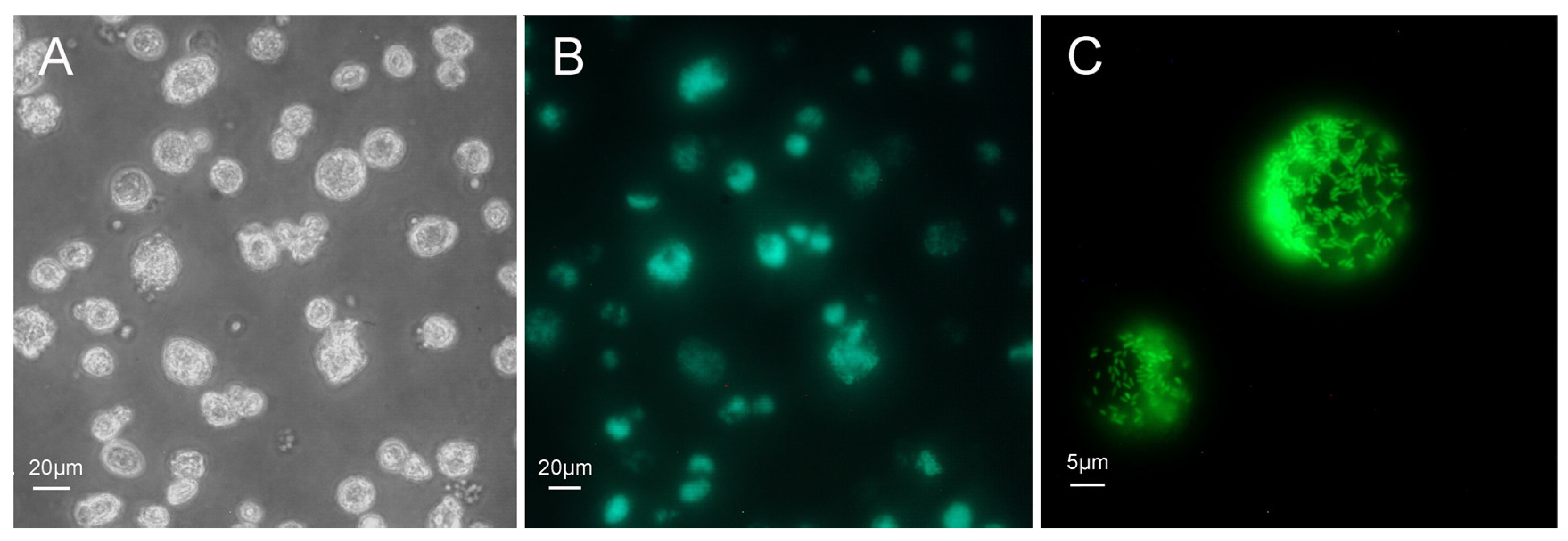{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction and History
2. Materials and Methods
2.1. Tick Cell Lines
2.2. Rickettsia buchneri
2.3. Challenge of Ixodes Cell Lines with Cell-Free R. buchneri
2.4. Microscopy of Cells Infected with Rickettsia buchneri
2.5. R. buchneri In Vitro Growth Dynamics: qPCR Analysis
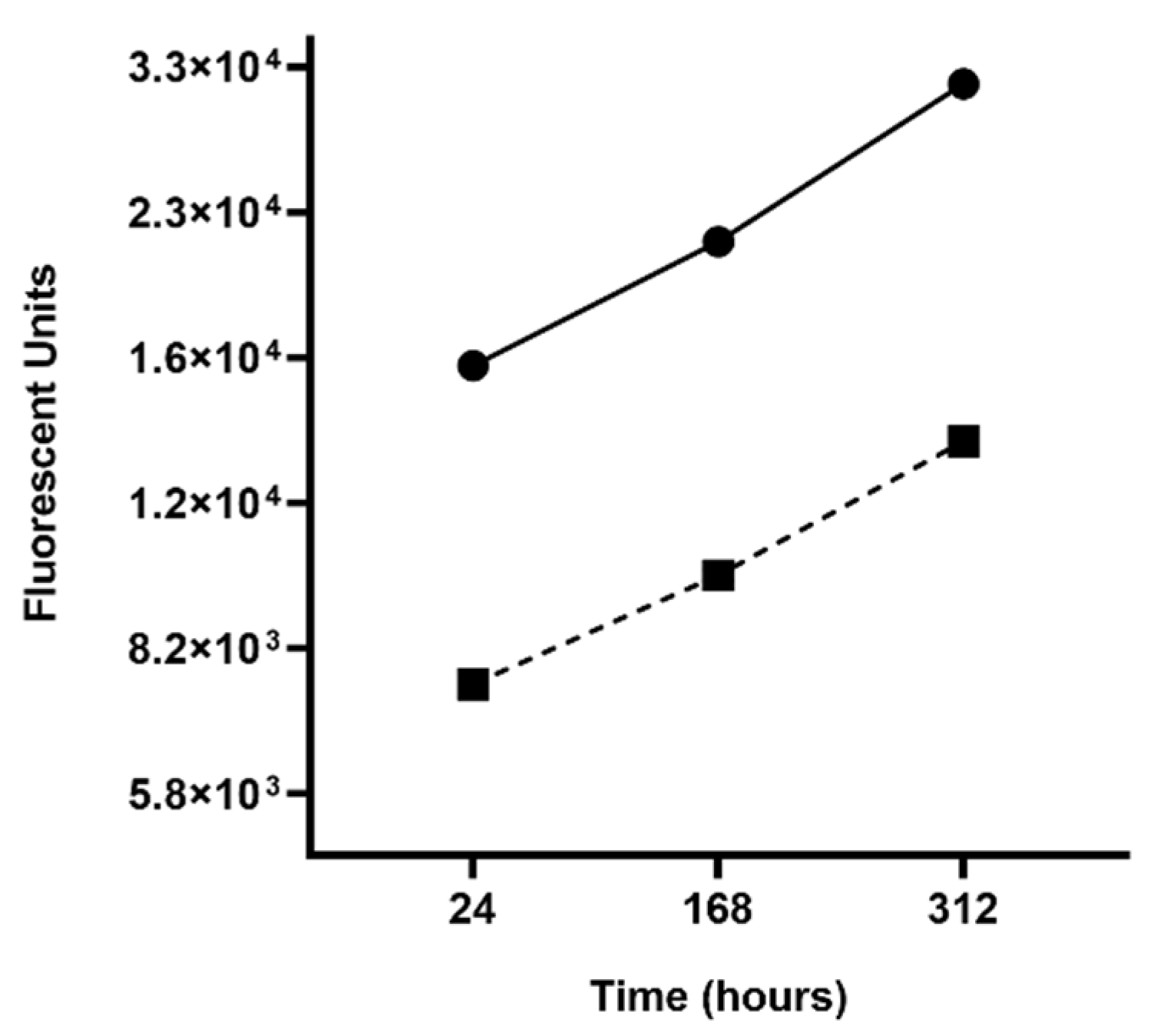
2.6. Fluorescence Plate Reader Analysis of R. buchneri Growth
2.7. RNA FISH: 23S rRNA Fast Red-Labeled Rickettsial Probe Design
2.8. In Vitro RNA FISH Assay
2.9. Tick Collection and Maintenance
2.10. RNA FISH Assay Dissected Whole Organs
3. Results
3.1. Challenge of Ixodes Cell Lines with R. buchneri
3.2. Microscopic Features of R. buchneri Growth in the Ixodes ricinus Cell Line IRE11
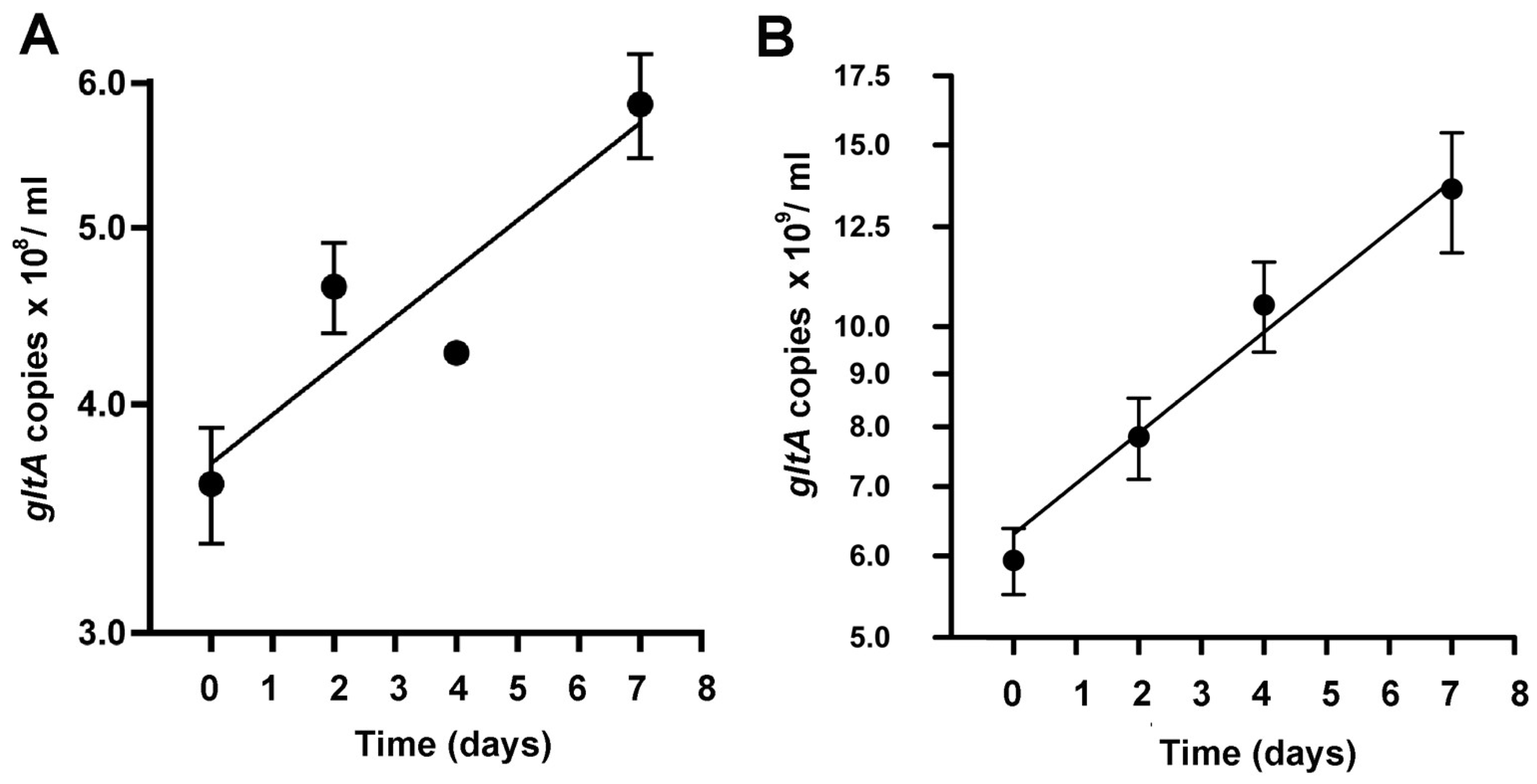
3.3. Growth Dynamics of R. buchneri in IRE11 Cells by qPCR
3.4. Growth Dynamics of R. buchneri in IRE11 Cells under Elevated Temperature Stress
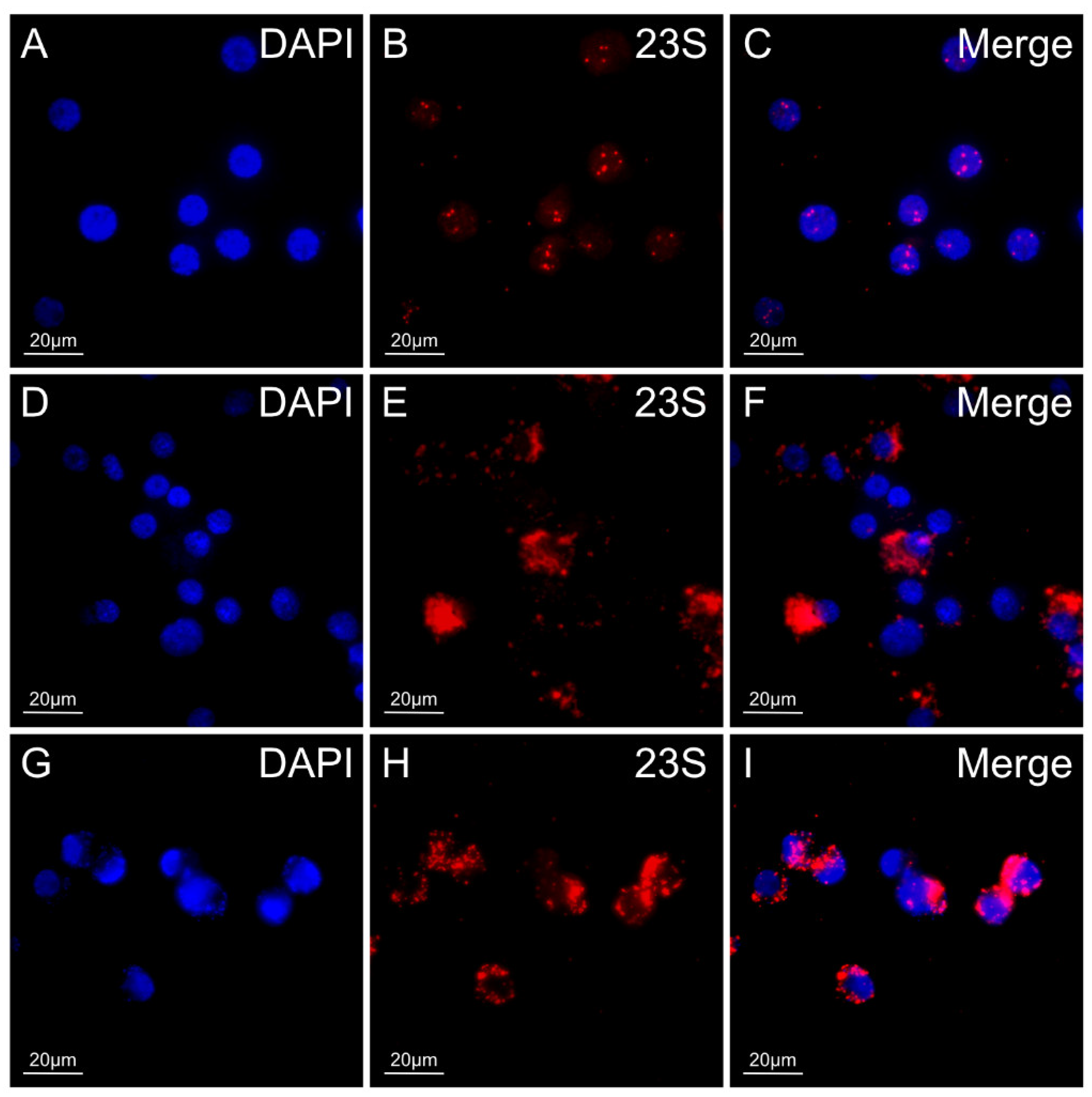
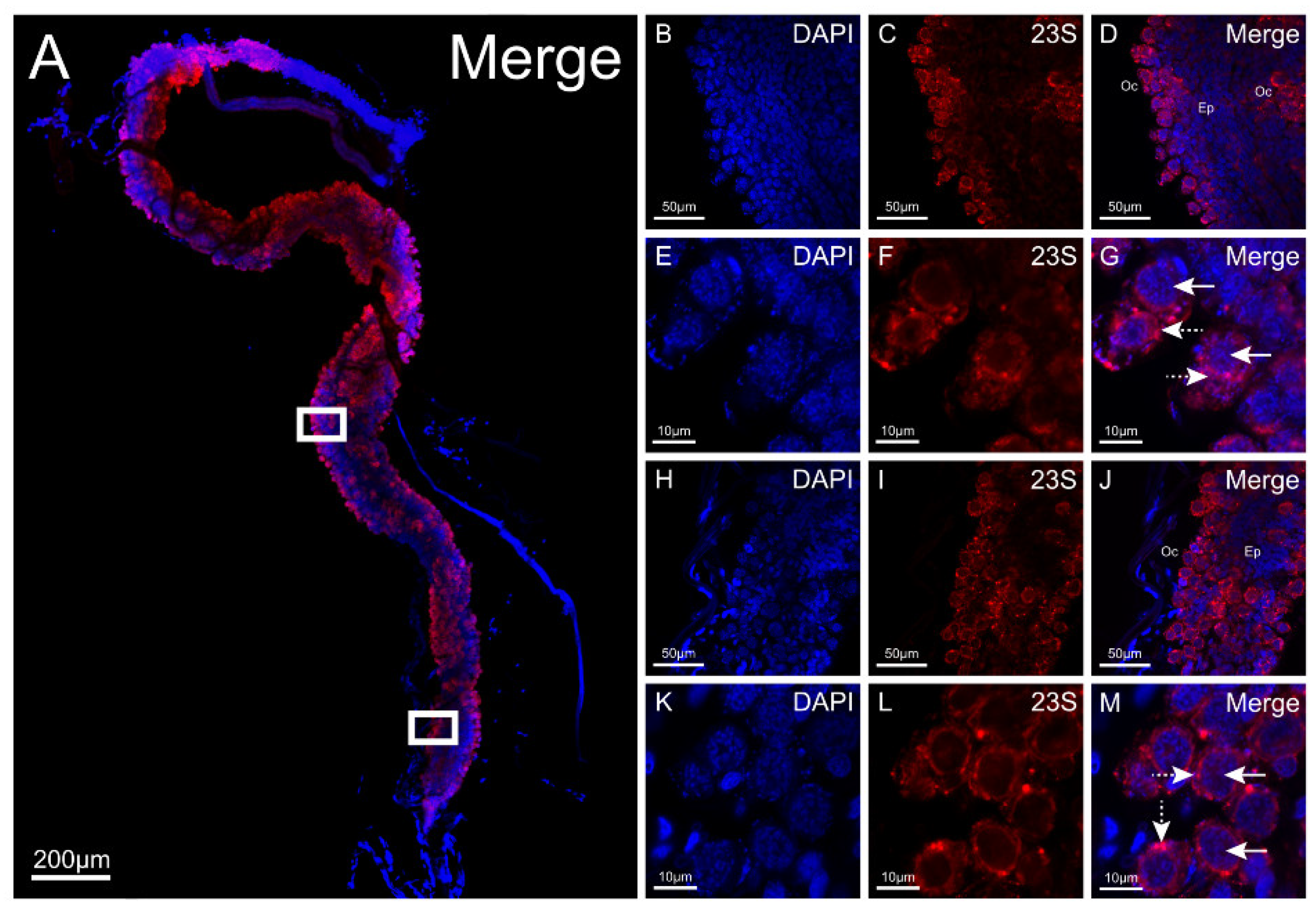
3.5. 23S rRNA FISH for Detection of R. buchneri in Ticks
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme disease—A tick-borne spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakken, J.S.; Dumler, J.S.; Chen, S.M.; Eckman, M.R.; Van Etta, L.L.; Walker, D.H. Human granulocytic ehrlichiosis in the upper Midwest United States. A new species emerging? JAMA 1994, 272, 212–218. [Google Scholar] [CrossRef]
- Piesman, J.; Spielman, A. Human babesiosis on Nantucket Island: Prevalence of Babesia microti in ticks. Am. J. Trop. Med. Hyg. 1980, 29, 742–746. [Google Scholar] [CrossRef]
- Telford, S.R., 3rd; Armstrong, P.M.; Katavolos, P.; Foppa, I.; Garcia, A.S.; Wilson, M.L.; Spielman, A. A new tick-borne encephalitis-like virus infecting New England deer ticks, Ixodes dammini. Emerg. Infect. Dis. 1997, 3, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Munderloh, U.G.; Liu, Y.; Wang, M.; Chen, C.; Kurtti, T.J. Establishment, maintenance and description of cell lines from the tick Ixodes scapularis. J. Parasitol. 1994, 80, 533–543. [Google Scholar] [CrossRef]
- Oliver, J.H., Jr.; Gregory, A.; Cummins, G.A.; Joiner, M.S. Immature Ixodes scapularis (Acari: Ixodidae) parasitizing lizards from the Southeastern, USA. J. Parasitol. 1993, 79, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Durden, L.A.; Oliver, J.H., Jr.; Banks, C.W.; Vogel, G.N. Parasitism of lizards by immature stages of the blacklegged tick, Ixodes scapularis (Acari, Ixodidae). Exp. Appl. Acarol. 2002, 26, 257–266. [Google Scholar] [CrossRef]
- Garvin, S.D.; Noden, B.H.; Dillwith, J.W.; Fox, S.F.; Payton, M.E.; Barker, R.W. Sylvatic Infestation of Oklahoma Reptiles with Immature Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2015, 52, 873–878. [Google Scholar] [CrossRef]
- Markowski, D.; Hyland, K.E.; Ginsberg, H.S.; Hu, R. Spatial distribution of larval Ixodes scapularis (Acari:Ixodidae) on Peromyscus leucopus and Microtus pennsylvanicus at two island sites. J. Parasitol. 1997, 83, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Cepeda, O.M.; Hazler, K.R.; Miller, M.C. Ecology of Lyme Disease: Habitat associations of ticks (Ixodes scapularis) in a rural landscape. Ecol. Applic. 1995, 5, 353–361. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Miller, M.C.; Hazler, K.R. Causes and consequences of tick (Ixodes scapularis) burdens on white-footed mice (Peromyscus leucopus). J. Mammal. 1996, 77, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Yabsley, M.J.; Nims, T.N.; Savage, M.Y.; Durden, L.A. Ticks and tick-borne pathogens and putative symbionts of black bears (Ursus americanus floridanus) from Georgia and Florida. J. Parasitol. 2009, 95, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Kodner, C.; Jarnefeld, J.; Eck, D.K.; Xu, Y. Agents of human anaplasmosis and Lyme disease at Camp Ripley, Minnesota. Vector Borne Zoonotic Dis. 2011, 11, 1529–1534. [Google Scholar] [CrossRef]
- Jara, R.F.; Wydeven, A.P.; Samuel, M.D. Gray wolf exposure to emerging vector-borne diseases in Wisconsin with comparison to domestic dogs and humans. PLoS ONE 2016, 11, e0165836. [Google Scholar] [CrossRef] [PubMed]
- Coipan, E.C.; Jahfari, S.; Fonville, M.; Maassen, C.B.; van der Giessen, J.; Takken, W.; Takumi, K.; Sprong, H. Spatiotemporal dynamics of emerging pathogens in questing Ixodes ricinus. Front. Cell Infect. Microbiol. 2013, 3, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef]
- Comer, K.M. Rocky Mountain spotted fever. Vet. Clin. N. Am. Small Anim. Pract. 1991, 21, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Beard, C.B.; Cordon-Rosales, C.; Durvasula, R.V. Bacterial symbionts of the triatominae and their potential use in control of Chagas disease transmission. Annu. Rev. Entomol. 2002, 47, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Utarini, A.; Indriani, C.; Ahmad, R.A.; Tantowijoyo, W.; Arguni, E.; Ansari, M.R.; Supriyati, E.; Wardana, D.S.; Meitika, Y.; Ernesia, I.; et al. Efficacy of Wolbachia-infected mosquito deployments for the control of Dengue. N. Engl. J. Med. 2021, 384, 2177–2186. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.L.; Maltz, M.; Aksoy, S. Obligate symbionts activate immune system development in the tsetse fly. J. Immunol. 2012, 188, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Loving, S.M.; Smith, A.B.; DiSalvo, A.F.; Burgdorfer, W. Distribution and prevalence of spotted fever group rickettsiae in ticks from South Carolina, with an epidemiological survey of persons bitten by infected ticks. Am. J. Trop. Med. Hyg. 1978, 27, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Magnarelli, L.A.; Anderson, J.F.; Philip, R.N.; Burgdorfer, W. Antibodies to spotted fever-group rickettsiae in dogs and prevalence of infected ticks in southern Connecticut. Am. J. Vet. Res. 1982, 43, 656–659. [Google Scholar] [PubMed]
- Billings, A.N.; Teltow, G.J.; Weaver, S.C.; Walker, D.H. Molecular characterization of a novel Rickettsia species from Ixodes scapularis in Texas. Emerg. Infect. Dis. 1998, 4, 305–309. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Joardar, V.; Williams, K.P.; Driscoll, T.; Hostetler, J.B.; Nordberg, E.; Shukla, M.; Walenz, B.; Hill, C.A.; Nene, V.M.; et al. A Rickettsia genome overrun by mobile genetic elements provides insight into the acquisition of genes characteristic of an obligate intracellular lifestyle. J. Bacteriol. 2012, 194, 376–394. [Google Scholar] [CrossRef] [Green Version]
- Kurtti, T.J.; Felsheim, R.F.; Burkhardt, N.Y.; Oliver, J.D.; Heu, C.C.; Munderloh, U.G. Rickettsia buchneri sp. nov., a rickettsial endosymbiont of the blacklegged tick Ixodes scapularis. Int. J. Syst. Evol. Microbiol. 2015, 65, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Cull, B.; Burkhardt, N.Y.; Wang, X.R.; Thorpe, C.J.; Oliver, J.D.; Kurtti, T.J.; Munderloh, U.G. Antibiotic gene clusters in the Ixodes scapularis symbiont Rickettsia buchneri and their potential role in exclusion of pathogenic bacteria from the tick. Front. Vet. Sci. under review.
- Magnarelli, L.A.; Andreadis, T.G.; Stafford, K.C., III; Holland, C.J. Rickettsiae and Borrelia burgdorferi in ixodid ticks. J. Clin. Microbiol. 1991, 29, 2798–2804. [Google Scholar] [CrossRef] [Green Version]
- Noda, H.; Munderloh, U.G.; Kurtti, T.J. Endosymbionts of ticks and their relationship to Wolbachia spp. and tick- borne pathogens of human and animals. Appl. Environ. Microbiol. 1997, 63, 3926–3932. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.D.; Price, L.D.; Burkhardt, N.Y.; Heu, C.C.; Khoo, B.S.; Thorpe, C.J.; Kurtti, T.J.; Munderloh, U.G. Growth dynamics and antibiotic elimination of symbiotic Rickettsia buchneri in the tick Ixodes scapularis (Acari: Ixodidae). Appl. Environ. Microbiol. 2021, 87, e01672-20. [Google Scholar] [CrossRef]
- Palomar, A.M.; Premchand-Branker, S.; Alberdi, P.; Belova, O.A.; Moniuszko-Malinowska, A.; Kahl, O.; Bell-Sakyi, L. Isolation of known and potentially pathogenic tick-borne microorganisms from European ixodid ticks using tick cell lines. Ticks Tick-Borne Dis. 2019, 10, 628–638. [Google Scholar] [CrossRef] [PubMed]
- Beliavskaia, A.; Hönig, V.; Erhart, J.; Vyhlidalova, T.; Palus, M.; Cerny, J.; Kozlova, I.; Ruzek, D.; Palomar, A.M.; Bell-Sakyi, L. Spiroplasma isolated from third-generation laboratory colony Ixodes persulcatus ticks. Front. Vet. Sci. 2021, 8, 659786. [Google Scholar] [CrossRef] [PubMed]
- Al-Rofaai, A.; Bell-Sakyi, L. Tick cell lines in research on tick control. Front. Physiol. 2020, 11, 152. [Google Scholar] [CrossRef] [Green Version]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a spotted fever group Rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Environ. Microbiol. 2002, 68, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.D.; Burkhardt, N.Y.; Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Motility characteristics are altered for Rickettsia bellii transformed to overexpress a heterologous rickA gene. Appl. Environ. Microbiol. 2014, 80, 1170–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, D.E.; Klompen, J.S.; Keirans, J.E.; Black, W.C., 4th. Population genetics of Ixodes scapularis (Acari: Ixodidae) based on mitochondrial 16S and 12S genes. J. Med. Entomol. 1996, 33, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef]
- Kurtti, T.J.; Burkhardt, N.Y.; Heu, C.C.; Munderloh, U.G. Fluorescent protein expressing Rickettsia buchneri and Rickettsia peacockii for tracking symbiont-tick cell interactions. Vet. Sci. 2016, 3, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkhardt, N.Y.; Baldridge, G.D.; Williamson, P.C.; Billingsley, P.M.; Heu, C.C.; Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Development of shuttle vectors for transformation of diverse Rickettsia species. PLoS ONE 2011, 6, e29511. [Google Scholar] [CrossRef]
- Kurtti, T.J.; Simser, J.A.; Baldridge, G.D.; Palmer, A.T.; Munderloh, U.G. Factors influencing in vitro infectivity and growth of Rickettsia peacockii (Rickettsiales: Rickettsiaceae) an endosymbiont of the Rocky Mountain wood tick, Dermacentor andersoni (Acari, Ixodidae). J. Invert. Pathol. 2005, 90, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtti, T.J.; Munderloh, U.G.; Johnson, R.C.; Ahlstrand, G.G. Colony formation and morphology in Borrelia burgdorferi. J. Clin. Microbiol. 1987, 25, 2054–2058. [Google Scholar] [CrossRef] [Green Version]
- Munderloh, U.G.; Hayes, S.F.; Cummings, J.; Kurtti, T.J. Microscopy of spotted fever rickettsia movement through tick cells. Microsc. Microanal. 1998, 4, 115–121. [Google Scholar] [CrossRef]
- Kurtti, T.J.; Munderloh, U.G.; Hayes, S.F.; Krueger, D.E.; Ahlstrand, G.G. Ultrastructural analysis of the invasion of tick cells by Lyme disease spirochetes (Borrelia burgdorferi) in vitro. Can. J. Zool. 1994, 72, 977–994. [Google Scholar] [CrossRef]
- Lynn, G.E.; Oliver, J.D.; Nelson, C.M.; Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Tissue distribution of the Ehrlichia muris-like agent in a tick vector. PLoS ONE 2015, 10, e0122007. [Google Scholar] [CrossRef]
- Stenos, J.; Graves, S.R.; Unsworth, N.B. A highly sensitive and specific real-time PCR assay for the detection of spotted fever and typhus group Rickettsiae. Am. J. Trop. Med. Hyg. 2005, 73, 1083–1085. [Google Scholar] [CrossRef]
- Hagen, R.; Verhoeve, V.I.; Gillespie, J.J.; Driscoll, T.P. Conjugative transposons and their cargo genes vary across natural populations of Rickettsia buchneri infecting the tick Ixodes scapularis. Genome Biol. Evol. 2018, 10, 3218–3229. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, J.M.; Nilsson, O.R.; Fischer, E.R.; Long, D.; Offerdahl, D.K.; Park, Y.; Scott, D.P.; Bloom, M.E. Dissecting Flavivirus biology in salivary gland cultures from fed and unfed Ixodes scapularis (Black-Legged Tick). mBio 2019, 10, e02628-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattila, J.T.; Munderloh, U.G.; Kurtti, T.J. Phagocytosis of the Lyme disease spirochete, Borrelia burgdorferi, by cells from the ticks, Ixodes scapularis and Dermacentor andersoni, infected with an endosymbiont, Rickettsia peacockii. J. Insect Sci. 2007, 7, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, K.H.; Haug, T. Ultrastructural, cytochemical, and immunocytochemical characterization of haemocytes of the hard tick Ixodes ricinus (Acari; Chelicerata). Cell Tissue Res. 1994, 277, 493–504. [Google Scholar] [CrossRef]
- Borovicková, B.; Hypsa, V. Ontogeny of tick hemocytes: A comparative analysis of Ixodes ricinus and Ornithodoros moubata. Exp. Appl. Acarol. 2005, 35, 317–333. [Google Scholar] [CrossRef]
- Chen, C.; Munderloh, U.G.; Kurtti, T.J. Cytogenetic characteristics of cell lines from Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1994, 31, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Kotsarenko, K.; Vetchova, P.; Lieskovska, J.; Füssy, Z.; Cabral-de-Mello, D.C.; Ryan, O.M.; Rego, R.O.M.; Alberdi, P.; Collins, M.; Bell-Sakyi, L.; et al. Karyotype changes in long-term cultured tick cell lines. Sci. Rep. 2020, 10, 13443. [Google Scholar] [CrossRef] [PubMed]
- Loginov, D.S.; Loginova, Y.F.; Dycka, F.; Böttinger, K.; Vechtova, P.; Sterba, J. Tissue-specific signatures in tick cell line MS profiles. Parasites Vectors 2019, 12, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, J.; Riedel, N.; Grittner, U.; Endres, M.; Banneke, S.; Emmrich, J.V. Body temperature measurement in mice during acute illness: Implantable temperature transponder versus surface infrared thermometry. Sci. Rep. 2018, 8, 3526. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The essential role of tick salivary glands and saliva in tick feeding and pathogen transmission. Front. Cell Infect. Microbiol. 2017, 7, 281. [Google Scholar] [CrossRef]
- da Silva Costa, L.F.; Nunes, P.H.; Soares, J.F.; Labruna, M.B.; Camargo-Mathias, M.I. Distribution of Rickettsia rickettsii in ovary cells of Rhipicephalus sanguineus (Latreille1806) (Acari: Ixodidae). Parasites Vectors 2011, 4, 222. [Google Scholar] [CrossRef] [Green Version]
- Niebylski, M.L.; Schrumpf, M.E.; Burgdorfer, W.; Fischer, E.R.; Gage, K.L.; Schwan, T.G. Rickettsia peacockii sp. nov., a new species infecting wood ticks, Dermacentor andersoni, in western Montana. Int. J. Syst. Bacteriol. 1997, 47, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Burgdorfer, W.; Hayes, S.G.; Mavros, A.J. Nonpathogenic rickettsiae in Dermacentor andersoni: A limiting factor for the distribution of Rickettsia rickettsia. In Rickettsiae and Rickettsial Diseases; Burgdorfer, W., Anacker, R.L., Eds.; Academic Press: New York, NY, USA, 1981; pp. 585–594. [Google Scholar]
- Al-Khafaji, A.M.; Armstrong, S.D.; Varotto Boccazzi, I.; Gaiarsa, S.; Sinha, A.; Li, Z.; Sassera, D.; Carlow, C.K.S.; Epis, S.; Makepeace, B.L. Rickettsia buchneri, symbiont of the deer tick Ixodes scapularis, can colonise the salivary glands of its host. Ticks Tick Borne Dis. 2020, 11, 101299. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Stafford, K.C., 3rd; Mather, T.N.; Yeh, M.T.; Horn, K.D.; Dumler, J.S. Hemocytic rickettsia-like organisms in ticks: Serologic reactivity with antisera to Ehrlichiae and detection of DNA of agent of human granulocytic ehrlichiosis by PCR. J. Clin. Microbiol. 1995, 33, 2710–2714. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.J.; Gawronski, J.D.; Eveleigh, D.E.; Benson, D.R. Intracellular symbionts and other bacteria associated with deer ticks (Ixodes scapularis) from Nantucket and Wellfleet, Cape Cod, Massachusetts. Appl. Environ. Microbiol. 2004, 70, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, C.X.; Moy, F.; Daniels, T.J.; Godfrey, H.P.; Cabello, F.C. Molecular analysis of microbial communities identified in different developmental stages of Ixodes scapularis ticks from Westchester and Dutchess Counties, New York. Environ. Microbiol. 2006, 8, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Trout Fryxell, R.T.; Steelman, C.D.; Szalanski, A.L.; Billingsley, P.M.; Williamson, P.C. Molecular detection of Rickettsia species within ticks (Acari: Ixodidae) collected from Arkansas United States. J. Med. Entomol. 2015, 52, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Nelder, M.P.; Russell, C.B.; Sheehan, N.J.; Sander, B.; Moore, S.; Li, Y.; Johnson, S.; Patel, S.N.; Sider, D. Human pathogens associated with the blacklegged tick Ixodes scapularis: A systematic review. Parasites Vectors 2016, 9, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thorpe, C.J.; Wang, X.-R.; Munderloh, U.G.; Kurtti, T.J. Tick Cell Culture Analysis of Growth Dynamics and Cellular Tropism of Rickettsia buchneri, an Endosymbiont of the Blacklegged Tick, Ixodes scapularis. Insects 2021, 12, 968. https://doi.org/10.3390/insects12110968
Thorpe CJ, Wang X-R, Munderloh UG, Kurtti TJ. Tick Cell Culture Analysis of Growth Dynamics and Cellular Tropism of Rickettsia buchneri, an Endosymbiont of the Blacklegged Tick, Ixodes scapularis. Insects. 2021; 12(11):968. https://doi.org/10.3390/insects12110968
Chicago/Turabian StyleThorpe, Cody J., Xin-Ru Wang, Ulrike G. Munderloh, and Timothy J. Kurtti. 2021. "Tick Cell Culture Analysis of Growth Dynamics and Cellular Tropism of Rickettsia buchneri, an Endosymbiont of the Blacklegged Tick, Ixodes scapularis" Insects 12, no. 11: 968. https://doi.org/10.3390/insects12110968