
New Cytogenetic Photomap and Molecular Diagnostics for the Cryptic Species of the Malaria Mosquitoes Anopheles messeae and Anopheles daciae from Eurasia

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
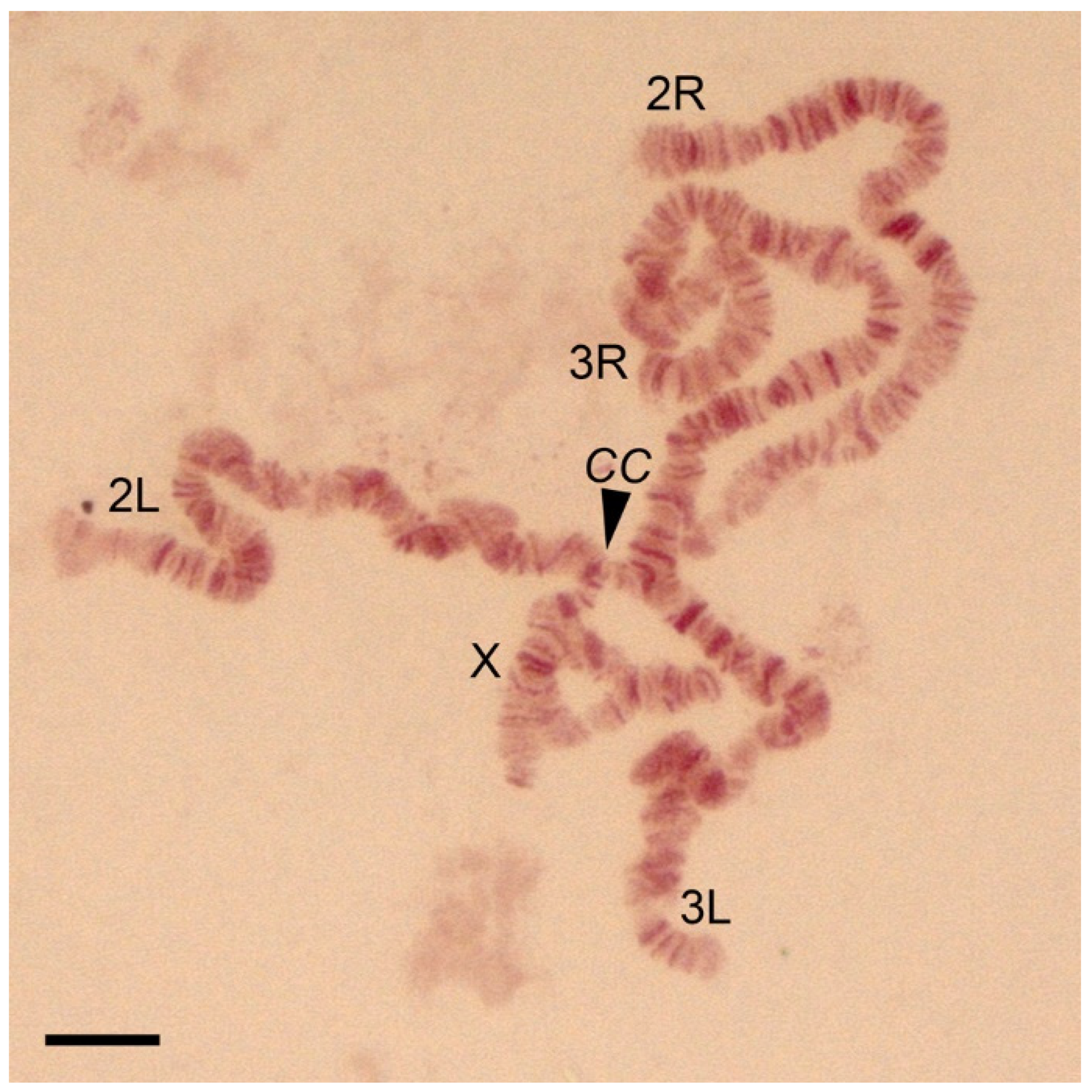
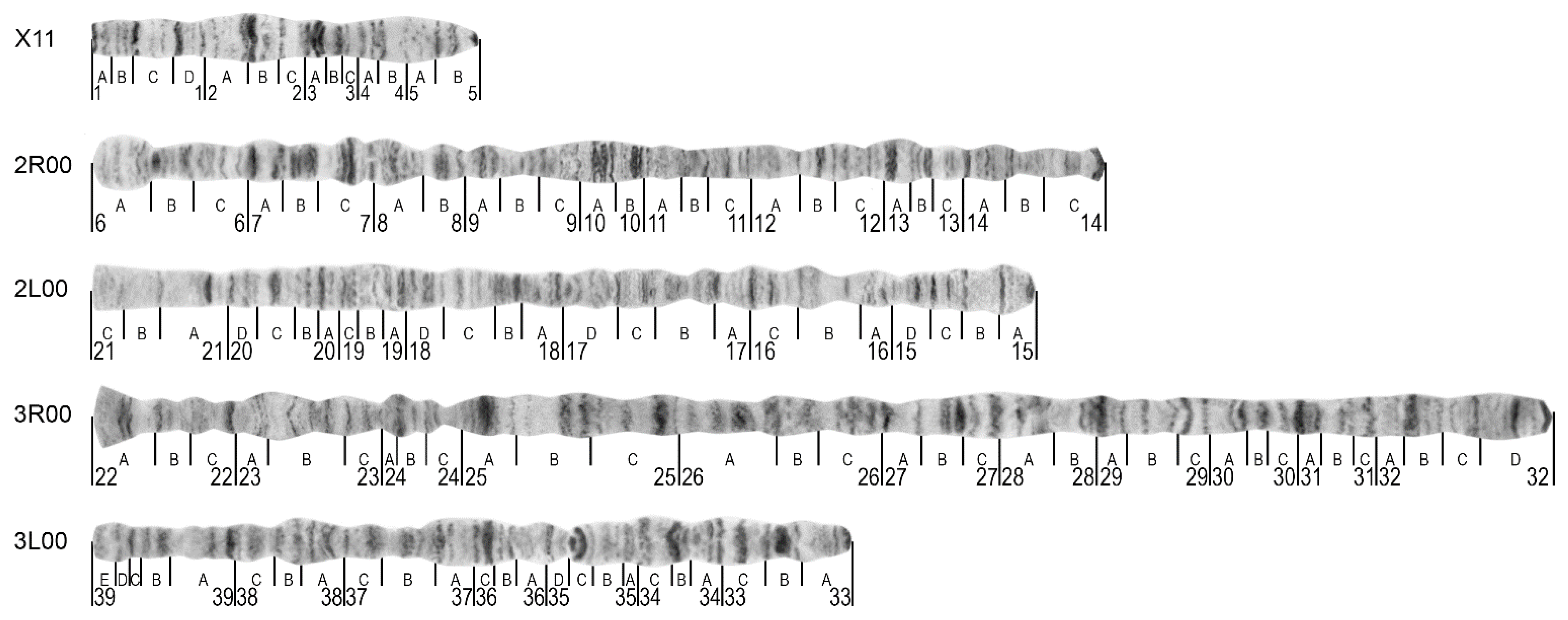
3.1. The Standard-Universal Cytogenetic Map for the Cryptic Species Anopheles messeae and Anopheles daciae
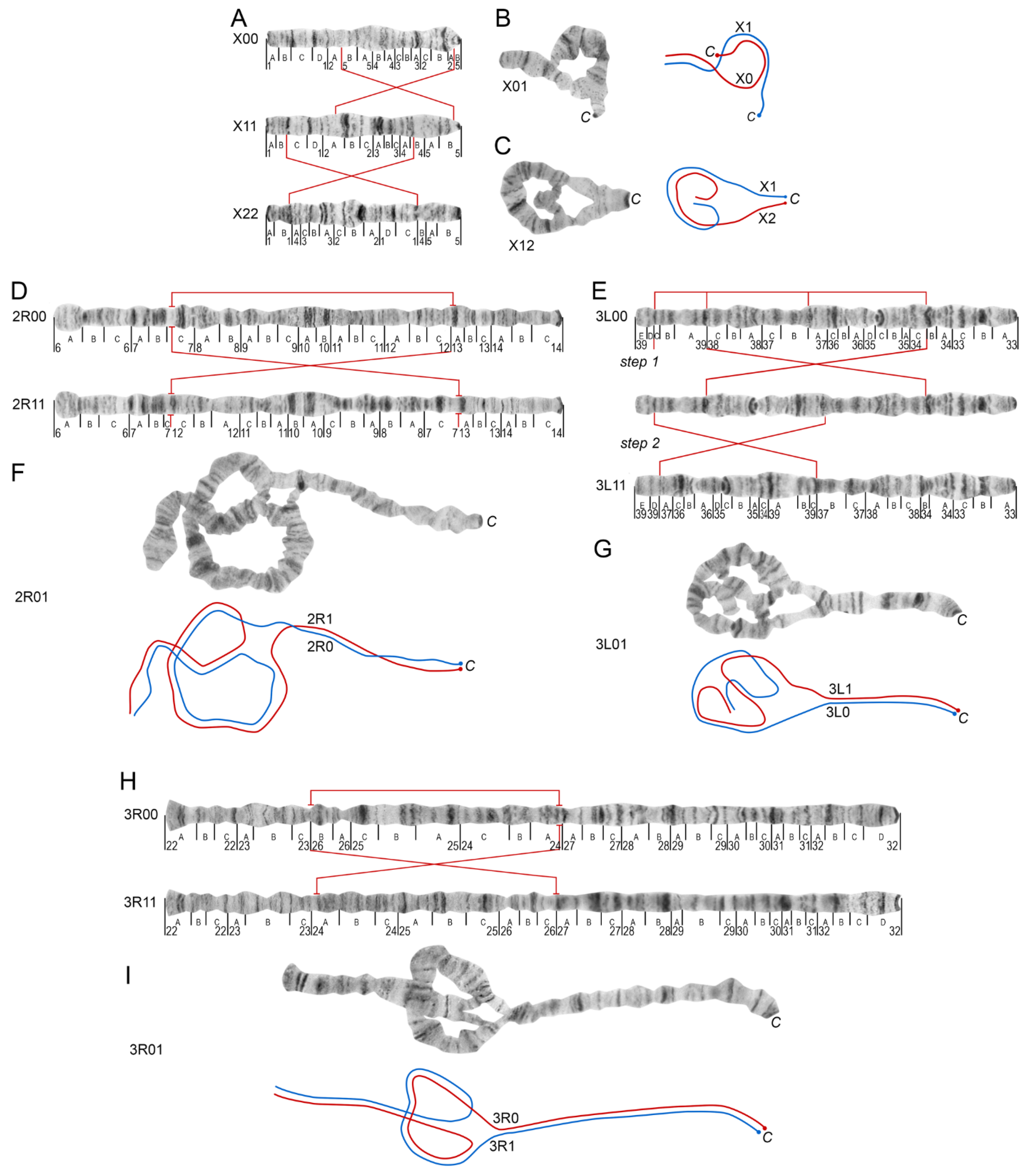
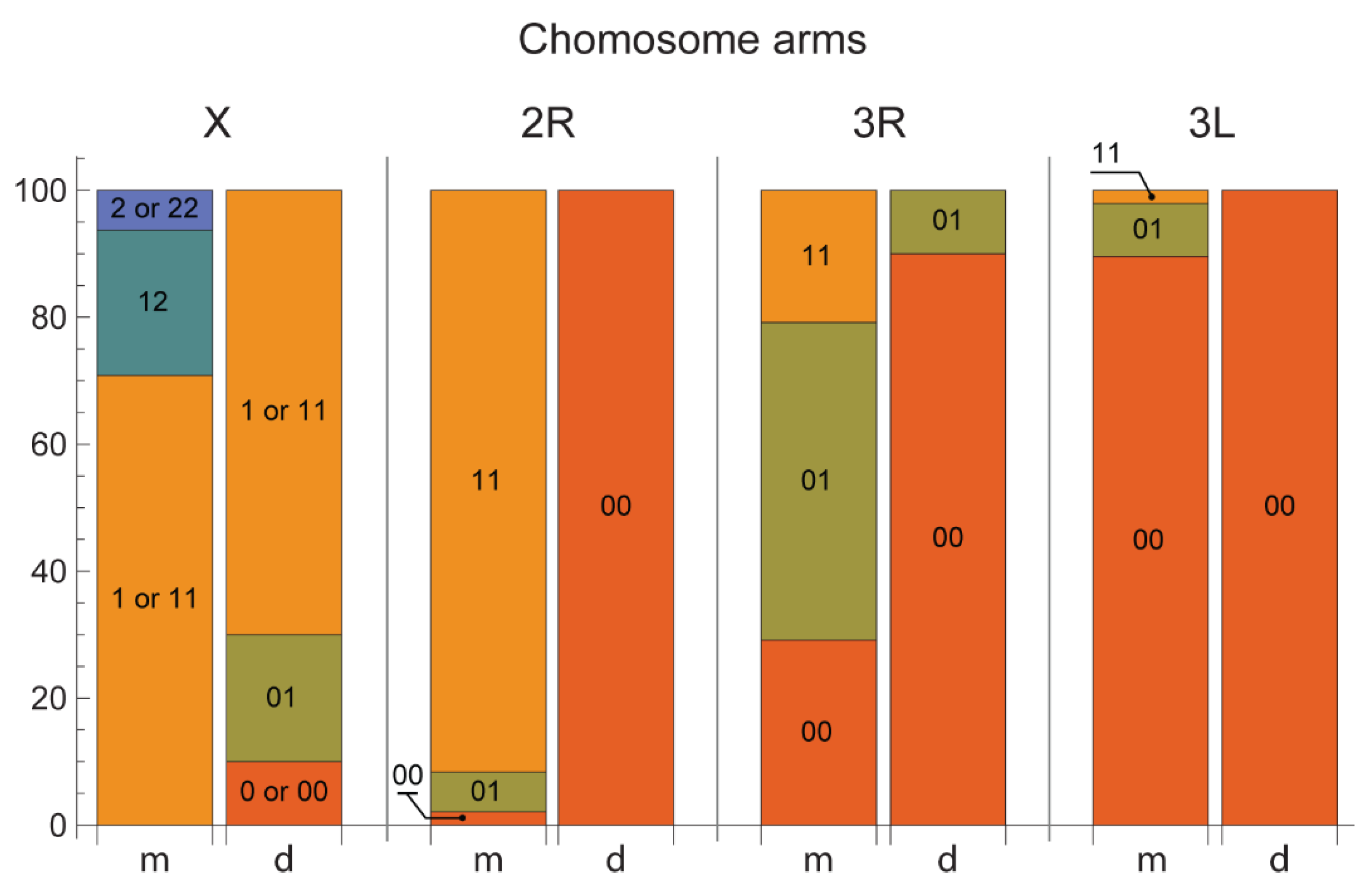
3.2. Chromosome Map Validation by the Analysis of Inversion Polymorphism in a Single Asian Location
3.3. A New ITS2-RFLP Approach for Anopheles messeae and Anopheles daciae Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic precis. Parasit Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gornostaeva, R.M.; Danilov, A.V. On distribution of malaria mosquitoes (Diptera, Culicidae: Anopheles) from Maculipennis Complex in Russian territory. Parasitologia 2002, 26, 33–47. [Google Scholar]
- Zvantsov, A.B.; Gordeev, M.I.; Goriacheva, I.I.; Ezhov, M.N. The distribution of the mosquitoes of the Anopheles maculipennis complex (Diptera, Culicidae, Anophelinae) in Central Asia. Med. Parazitol. (Mosk) 2014, 4, 19–23. [Google Scholar]
- Naumenko, A.N.; Karagodin, D.A.; Yurchenko, A.A.; Moskaev, A.V.; Martin, O.I.; Baricheva, E.M.; Sharakhov, I.V.; Gordeev, M.I.; Sharakhova, M.V. Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae. Genes 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegniy, V.N. Population Genetics and Evolution of Malaria Mosquitoes; Tomsk State University Publisher: Tomsk, Russia, 1991; p. 137. [Google Scholar]
- Kabanova, V.M.; Kartashova, N.N.; Stegnii, V.N. Karyological study of natural populations of malarial mosquitoes in the Middle Ob river. I. Characteristics of the karyotype of Anopheles maculipennis messeae. Tsitologiia 1972, 14, 630–636. [Google Scholar] [PubMed]
- Gordeev, M.I.; Sibataev, A.K. Cytogenetic and phenotypic variation in central and peripheral populations of the malaria mosquito, Anopheles messeae Fall. (Diptera, Culicidae). Genetika 1996, 32, 1199–1205. [Google Scholar]
- Gordeev, M.I.; Bezzhonova, O.V.; Moskaev, A.V. Chromosomal polymorphism in the populations of malaria mosquito Anopheles messeae (Diptera, Culicidae) at the south of Russian Plain. Genetika 2012, 48, 1124–1128. [Google Scholar] [PubMed]
- Stegnii, V.N.; Kabanova, V.M.; Novikov Iu, M. Study of the karyotype of the malaria mosquito. Tsitologiia 1976, 18, 760–766. [Google Scholar]
- Gordeev, M.I.; Stegnii, V.N. Inversion polymorphism of the malaria mosquito Anopheles messeae. VIII. Distribution of larvae in biotopes under conditions of co-habitation with Anopheles beklemishevi. Genetika 1989, 25, 283–291. [Google Scholar] [PubMed]
- Gordeev, M.I.; Stegnii, V.N. Inversion polymorphism in the malaria mosquito Anopheles messeae. VII. Fertility and the population genetics structure of the species. Genetika 1987, 23, 2169–2174. [Google Scholar] [PubMed]
- Novikov, I.M.; Kabanova, V.M. Adaptive association of inversions in a natural population of the malaria mosquito Anopheles messeae Fall. Genetika 1979, 15, 1033–1045. [Google Scholar] [PubMed]
- Novikov, I.M.; Shevchenko, A.I. Inversion polymorphism and the divergence of two cryptic forms of Anopheles messeae (Diptera, Culicidae) at the level of genomic DNA repeats. Genetika 2001, 37, 915–925. [Google Scholar] [PubMed]
- Vaulin, O.V.; Novikov, Y.M. Geographic variability of ITS2 rDNA and COI mtDNA and cryptic species of malaria mosquito Anopheles messeae Fall. (Diptera: Culicidae). Vestn. VOGIS 2010, 14, 546–555. [Google Scholar]
- Vaulin, O.V.; Karagodin, D.A.; Zakharov, I.K.; Baricheva, E.M. The dynamics of the species composition of malaria mosquitoes in Siberian populations, detected using restriction analysis. Genetika 2018, 54, 832–842. [Google Scholar]
- Novikov, Y.M.; Shevchenko, A.I. Cryptic species of Anopheles messeae sensu lato (Diptera: Culicidae), their identification, features and nomenclature. Tomsk. State Univ. J. Biol. 2020, 51, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, G.; Linton, Y.M.; Vladimirescu, A.; Howard, T.M.; Harbach, R.E. Mosquitoes of the Anopheles maculipennis group (Diptera: Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence. Bull. Entomol. Res. 2004, 94, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Danabalan, R.; Monaghan, M.T.; Ponsonby, D.J.; Linton, Y.M. Occurrence and host preferences of Anopheles maculipennis group mosquitoes in England and Wales. Med. Vet. Entomol. 2014, 28, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Kitzmiller, J.B.; Frizzi, G.; Baker, R. Evolution and speciation within the Maculipennis complex of the genus Anopheles. In Genetics of Insect Vectors of Disease; Wright, J.W., Ed.; Elsevier Publishing Company: Amsterdam, The Netherlands; London, UK; New York, NY, USA, 1967; pp. 151–210. [Google Scholar]
- Rai, K.S. A comparative study of mosquito karyotypes. Ann. Entomol. Soc. Am. 1963, 56, 160–170. [Google Scholar] [CrossRef]
- Kabanova, V.M.; Kartashova, N.N.; Stegii, V.N. Kariological study of malaria mosquito in the Middle Ob region. Report 1. Characteristics of the Anopheles maculipennis messeae karyotype. Tsitologiya 1972, 14, 630–636. [Google Scholar]
- Coluzzi, M.; Sabatini, A. Cytotaxonomic observations of species A and B of Anopheles gambiae complex. Parassitologia 1967, 9, 73–88. [Google Scholar]
- Subbarao, S. Genetics of malaria vectors. Proc. Nat. Acad. Sci. USA 1996, 66, 51–76. [Google Scholar]
- Sharma, G.P.R.; Parshad, R.; Narang, S.L.; Kitzmiller, J.B. Tha salivary gland chromosomes of Anopheles stephensi stephensi. J. Med. Entomol. 1969, 6, 68–71. [Google Scholar] [CrossRef]
- Artemov, G.N.; Sharakhova, M.V.; Naumenko, A.N.; Karagodin, D.A.; Baricheva, E.M.; Stegniy, V.N.; Sharakhov, I.V. A standard photomap of ovarian nurse cell chromosomes in the European malaria vector Anopheles atroparvus. Med. Vet. Entomol 2015, 29, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Artemov, G.N.; Velichevskaya, A.I.; Bondarenko, S.M.; Karagyan, G.H.; Aghayan, S.A.; Arakelyan, M.S.; Stegniy, V.N.; Sharakhov, I.V.; Sharakhova, M.V. A standard photomap of the ovarian nurse cell chromosomes for the dominant malaria vector in Europe and Middle East Anopheles sacharovi. Malar. J. 2018, 17, 276. [Google Scholar] [CrossRef] [Green Version]
- Artemov, G.N.; Gordeev, M.I.; Kokhanenko, A.A.; Moskaev, A.V.; Velichevskaya, A.I.; Stegniy, V.N.; Sharakhov, I.V.; Sharakhova, M.V. A standard photomap of ovarian nurse cell chromosomes and inversion polymorphism in Anopheles beklemishevi. Parasit Vectors 2018, 11, 211. [Google Scholar] [CrossRef] [PubMed]
- Lilja, T.; Eklof, D.; Jaenson, T.G.T.; Lindstrom, A.; Terenius, O. Single nucleotide polymorphism analysis of the ITS2 region of two sympatric malaria mosquito species in Sweden: Anopheles daciae and Anopheles messeae. Med. Vet. Entomol. 2020, 34, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezzhonova, O.V.; Goryacheva, I.I. Intragenomic heterogeneity of rDNA internal transcribed spacer 2 in Anopheles messeae (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Kronefeld, M.; Dittmann, M.; Zielke, D.; Werner, D.; Kampen, H. Molecular confirmation of the occurrence in Germany of Anopheles daciae (Diptera, Culicidae). Parasit Vectors 2012, 5, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaulin, O.V.; Novikov Iu, M. Polymorphism and interspecific variability of cytochrome oxidase subunit I (COI) gene nucleotide sequence in sibling species of A and B Anopheles messeae and An. beklemishevi (Diptera: Culicidae). Russ. J. Genet. Appl. Res. 2012, 2, 421–429. [Google Scholar] [CrossRef]
- Artemov, G.N.; Stegniy, V.N.; Sharakhova, M.V.; Sharakhov, I.V. The Development of Cytogenetic Maps for Malaria Mosquitoes. Insects 2018, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Beebe, N.W.; Saul, A. Discrimination of all members of the Anopheles punctulatus complex by polymerase chain reaction—restriction fragment length polymorphism analysis. Am. J. Trop. Med. Hyg. 1995, 53, 478–481. [Google Scholar] [CrossRef]
- Tesler, G. GRIMM: Genome rearrangements web server. Bioinformatics 2002, 18, 492–493. [Google Scholar] [CrossRef] [PubMed]
- Microsoft, C. Available online: https://office.microsoft.com/excel (accessed on 20 March 2021).
- Paradis, E. Pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weir, B.S.; Hill, W.G. Estimating F-statistics. Annu. Rev. Genet. 2002, 36, 721–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradburg, G. BEDASSLE: Quantifies Effects of Geo/Eco Distance on Genetic Differentiation. Available online: https://CRAN.R-project.org/package=BEDASSLE (accessed on 20 March 2021).
- Zhimulev, I.F. Polytene chromosomes, heterochromatin, and position effect variegation. Adv. Genet. 1998, 37, 1–566. [Google Scholar]
- Gordeev, M.; Moskaev, A.; Brusentsov, I.; Karagodin, D.; Burlak, V.; Artemov, G.; Sharakhov, I.; Sharakhova, M. Chromosomal Polymorphism of Anopheles daciae and An. messeae malaria mosquitoes. In Proceedings of the VΙΙ Internatiional Scientific and Practical Conference, “Actual Problems Biological and Chemical Ecology”, Moscow, Russia, 21–22 April 2021. [Google Scholar]
- Stegnii, V.N.; Sharakhova, M.V. Systemic reorganization of the architechtonics of polytene chromosomes in onto- and phylogenesis of malaria mosquitoes. Structural features regional of chromosomal adhesion to the nuclear membrane. Genetika 1991, 27, 828–835. [Google Scholar] [PubMed]
- George, P.; Sharakhova, M.V.; Sharakhov, I.V. High-resolution cytogenetic map for the African malaria vector Anopheles gambiae. Insect Mol. Biol. 2010, 19, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Stegnii, V.N. Inversion polymorphism of the malarial mosquito Anopheles messeae. IV. The stability of the frequency distribution of the inversions by species area. Genetika 1983, 19, 466–473. [Google Scholar] [PubMed]
- Zhimulev, I.F. Morphology and structure of polytene chromosomes. Adv. Genet. 1996, 39, 1–497. [Google Scholar]
- Sharakhova, M.V.; Sharakhov, I.V.; Stegnii, V.N. Chromocenter organization in salivary glands of the malaria mosquito Anopheles messeae Fall. Genetika 1997, 33, 196–201. [Google Scholar] [PubMed]
- Artemov, G.N.; Bondarenko, S.M.; Naumenko, A.N.; Stegniy, V.N.; Sharakhova, M.V.; Sharakhov, I.V. Partial-arm translocations in evolution of malaria mosquitoes revealed by high-coverage physical mapping of the Anopheles atroparvus genome. BMC Genom. 2018, 19, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharakhova, M.V.; Xia, A.; McAlister, S.I.; Sharakhov, I.V. A standard cytogenetic photomap for the mosquito Anopheles stephensi (Diptera: Culicidae): Application for physical mapping. J. Med. Entomol. 2006, 43, 861–866. [Google Scholar] [CrossRef] [Green Version]
- Perevozkin, V.P.; Gordeev, M.I.; Bondarchuk, S.S. Chromosome polymorphism and regularities of the subpopulation organization of malaria mosquitoes Anopheles (Diptera, Culicidae) in biotopes of the Tomsk oblast. Genetika 2009, 45, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Stegniy, V.N.; Pishchelko, A.O.; Sibataev, A.K.; Abylkassymova, G. Spatial and temporal variations of the chromosomal inversion frequencies across the range of malaria mosquito Anopheles messeae Fall. (Culicidae) during the 40-year monitoring period. Genetika 2016, 52, 664–671. [Google Scholar] [CrossRef]
- Perevozkin, V.P.; Gordeev, M.I.; Nikolaeva, N.V.; Becker, N. The population-and-species-specific structure of Anopheles (Diptera, Culicidae) mosquitoes in the Upper Rhine Valley, Germany. Med. Parazitol. (Mosk) 2010, 2, 27–32. [Google Scholar]
- Gordeev, M.I.; Moskaev, A.V. Chromosomal polymorphism in the populations of malaria mosquito Anopheles messeae (Diptera, Culicidae) in the Volga region. Genetika 2016, 52, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Stegniy, V.N.; Pischelko, A.O.; Sibataev, A.K.; Abylkasymova, G. Spatial-temporal dynamics of inversion frequencies along the distribution of malaria mosquito Anopheles messeae Fall. (Culicidae). Genetica 2016, 52, 664–671. [Google Scholar]
- Collins, F.H.; Paskewitz, S.M. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol. Biol. 1996, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Paskewitz, S.M.; Wesson, D.M.; Collins, F.H. The internal transcribed spacers of ribosomal DNA in five members of the Anopheles gambiae species complex. Insect Mol. Biol. 1993, 2, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Hackett, B.J.; Gimnig, J.; Guelbeogo, W.; Costantini, C.; Koekemoer, L.L.; Coetzee, M.; Collins, F.H.; Besansky, N.J. Ribosomal DNA internal transcribed spacer (ITS2) sequences differentiate Anopheles funestus and An. rivulorum, and uncover a cryptic taxon. Insect Mol. Biol. 2000, 9, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Beebe, N.W.; Maung, J.; van den Hurk, A.F.; Ellis, J.T.; Cooper, R.D. Ribosomal DNA spacer genotypes of the Anopheles bancroftii group (Diptera: Culicidae) from Australia and Papua New Guinea. Insect Mol. Biol. 2001, 10, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, R.C.; Reinert, J.F.; Li, C. Ribosomal DNA ITS2 sequences differentiate six species in the Anopheles crucians complex (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.H.; Porter, C.H.; Cope, S.E. Comparison of rDNA and mtDNA in the sibling species Anopheles freeborni and A. hermsi. Am. J. Trop. Med. Hyg. 1990, 42, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Cornel, A.J.; Porter, C.H.; Collins, F.H. Polymerase chain reaction species diagnostic assay for Anopheles quadrimaculatus cryptic species (Diptera: Culicidae) based on ribosomal DNA ITS2 sequences. J. Med. Entomol. 1996, 33, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Gordeev, M.I.; Zvantsov, A.B.; Goriacheva, I.I.; Shaikevich, E.V.; Ezhov, M.N. Description of the new species Anopheles artemievi sp.n. (Diptera, Culicidae). Med. Parazitol. (Mosk) 2005, 2, 4–5. [Google Scholar]
- Sedaghat, M.M.; Linton, Y.M.; Oshaghi, M.A.; Vatandoost, H.; Harbach, R.E. The Anopheles maculipennis complex (Diptera: Culicidae) in Iran: Molecular characterization and recognition of a new species. Bull. Entomol. Res. 2003, 93, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Kampen, H. The ITS2 ribosomal DNA of Anopheles beklemishevi and further remarks on the phylogenetic relationships within the Anopheles maculipennis group of species (Diptera: Culicidae). Parasitol. Res. 2005, 97, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Kampen, H. Integration of Anopheles beklemishevi (Diptera: Culicidae) in a PCR assay diagnostic for palaearctic Anopheles maculipennis sibling species. Parasitol. Res. 2005, 97, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Proft, J.; Maier, W.A.; Kampen, H. Identification of six sibling species of the Anopheles maculipennis complex (Diptera: Culicidae) by a polymerase chain reaction assay. Parasitol. Res. 1999, 85, 837–843. [Google Scholar] [CrossRef]
- Hodge, J.M.; Yurchenko, A.A.; Karagodin, D.A.; Masri, R.A.; Smith, R.C.; Gordeev, M.I.; Sharakhova, M.V. The new Internal Transcribed Spacer 2 diagnostic tool clarifies the taxonomic position and geographic distribution of the North American malaria vector Anopheles Punctipennis. Malar. J. 2021, 20, 141. [Google Scholar] [CrossRef]
- Calzolari, M.; Desiato, R.; Albieri, A.; Bellavia, V.; Bertola, M.; Bonilauri, P.; Callegari, E.; Canziani, S.; Lelli, D.; Mosca, A.; et al. Mosquitoes of the Maculipennis complex in Northern Italy. Sci. Rep. 2021, 11, 6421. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosomes | X | 2 | 3 |
|---|---|---|---|
| Average length (μm) | 58.5 ± 6.1 | 297.5 ± 40.1 | 312.9 ± 36.9 |
| Relative length (%) | 8.6 | 44.5 | 46.9 |
| Relative short arm length (%) | NA | 46.9 | 38.5 |
| Chromosome or Chromosome Arms | Telomere | Additional Landmarks |
|---|---|---|
| X | Sharp end with the thin dark band in 1A | Puffed region in 2B division starting with a dark band, and the light area with diffused bands in divisions 4B–5A |
| 2R | Often round shape end with wide pale bands followed by a dark band in the 6B division | Two sets of dark bands in the middle of the arm in the 10A,B division |
| 2L | Long, light end limited by a thin band in the 21A division | No additional landmarks identified |
| 3R | Short flared end with two thin bands in division 22A | A neck-like region in 24C followed by 3 sets of several dark bands in the 25A, 25B, and 25C divisions |
| 3L | A set of five bands in 39B–E | A “bird’s eye” landmark in the 35C region—dot-like band coupled with long and dark often curved bands |
| Chromosomal Variant | An. messeae | An. daciae |
|---|---|---|
| X1 | 81.5% | 77.8% |
| X2 | 18.5% | 0% |
| 2R1 | 94.8% | 0% |
| 3R1 | 45.8% | 5% |
| 3L1 | 6.3% | 0% |
| Chromosomes | HWE, p-Value An. messeae | HWE, p-Value An. daciae | Fst |
|---|---|---|---|
| X | - | - | 0.091 |
| 2R | 0.104 | 1.000 | 0.915 |
| 3R | 1.000 | 1.000 | 0.261 |
| 3L | 0.153 | 1.000 | 0.000 |
| Total | - | - | 0.629 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artemov, G.N.; Fedorova, V.S.; Karagodin, D.A.; Brusentsov, I.I.; Baricheva, E.M.; Sharakhov, I.V.; Gordeev, M.I.; Sharakhova, M.V. New Cytogenetic Photomap and Molecular Diagnostics for the Cryptic Species of the Malaria Mosquitoes Anopheles messeae and Anopheles daciae from Eurasia. Insects 2021, 12, 835. https://doi.org/10.3390/insects12090835
Artemov GN, Fedorova VS, Karagodin DA, Brusentsov II, Baricheva EM, Sharakhov IV, Gordeev MI, Sharakhova MV. New Cytogenetic Photomap and Molecular Diagnostics for the Cryptic Species of the Malaria Mosquitoes Anopheles messeae and Anopheles daciae from Eurasia. Insects. 2021; 12(9):835. https://doi.org/10.3390/insects12090835
Chicago/Turabian StyleArtemov, Gleb N., Valentina S. Fedorova, Dmitriy A. Karagodin, Ilya I. Brusentsov, Elina M. Baricheva, Igor V. Sharakhov, Mikhail I. Gordeev, and Maria V. Sharakhova. 2021. "New Cytogenetic Photomap and Molecular Diagnostics for the Cryptic Species of the Malaria Mosquitoes Anopheles messeae and Anopheles daciae from Eurasia" Insects 12, no. 9: 835. https://doi.org/10.3390/insects12090835