
Molecular Characterizations and Functional Analyses of LmR2D2 in the Locusta migratoria siRNA Pathway


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Isolation and Sequencing of cDNAs Encoding LmR2D2
2.3. Expression Patterns of the LmR2D2 Gene
2.4. RNAi of RNAi Experiment
2.5. Prokaryotic Expression and Purification of LmR2D2 Protein
2.6. Western Blot Analysis of LmR2D2
2.7. Binding Assay of LmR2D2 Protein and dsRNA
2.8. Statistical Analysis
3. Results
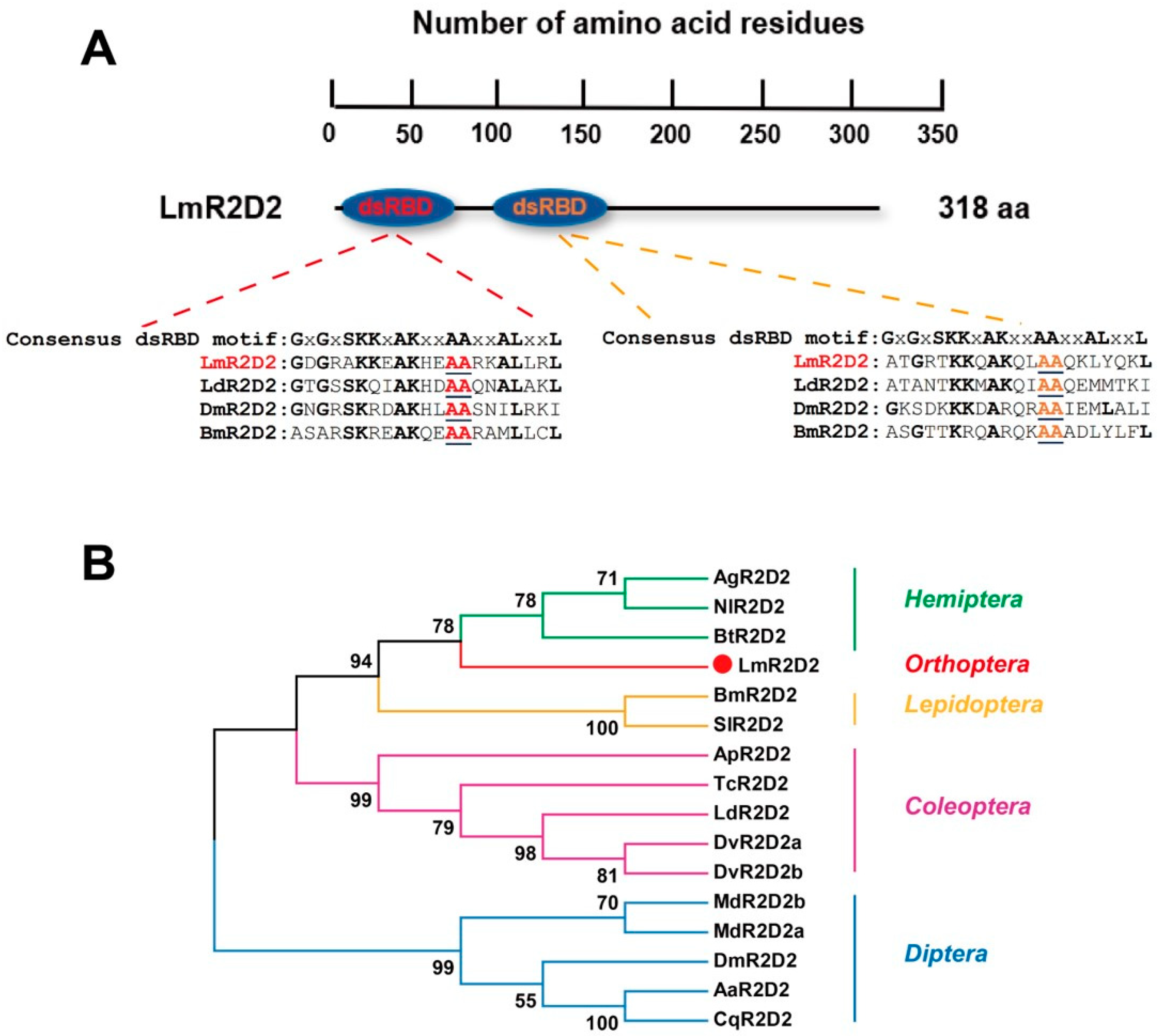
3.1. Domain Structure and Phylogenetic Analysis of Deduced LmR2D2 Protein
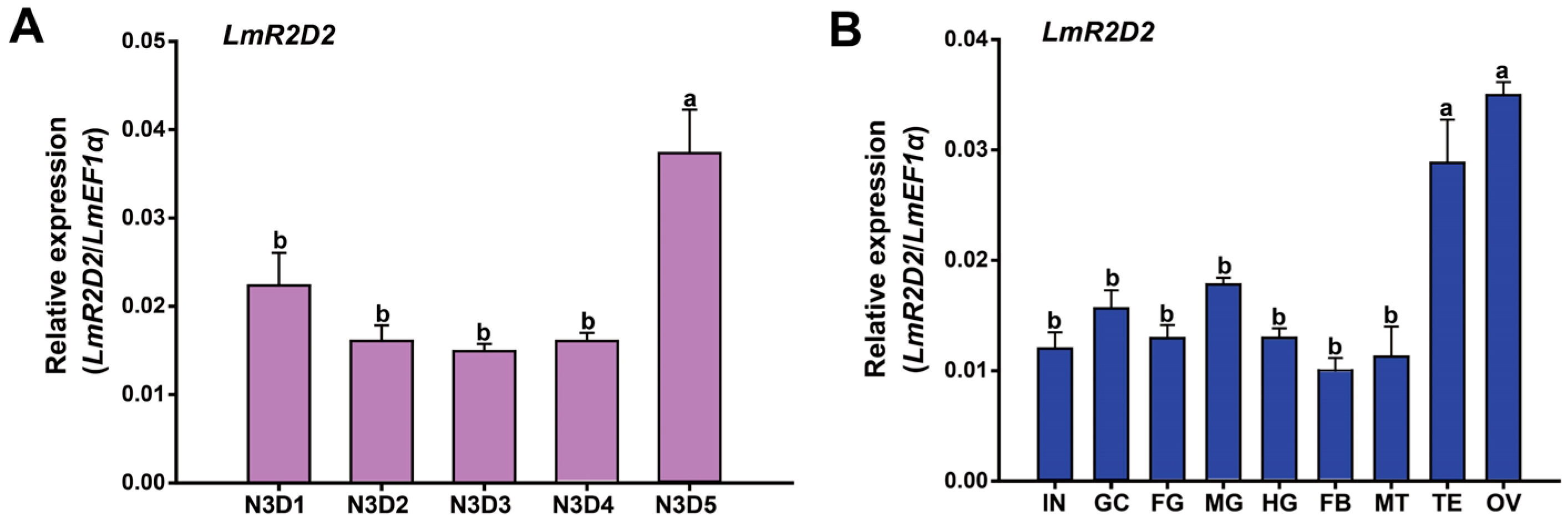
3.2. Developmental and Tissue-Specific Expression Profiles of LmR2D2
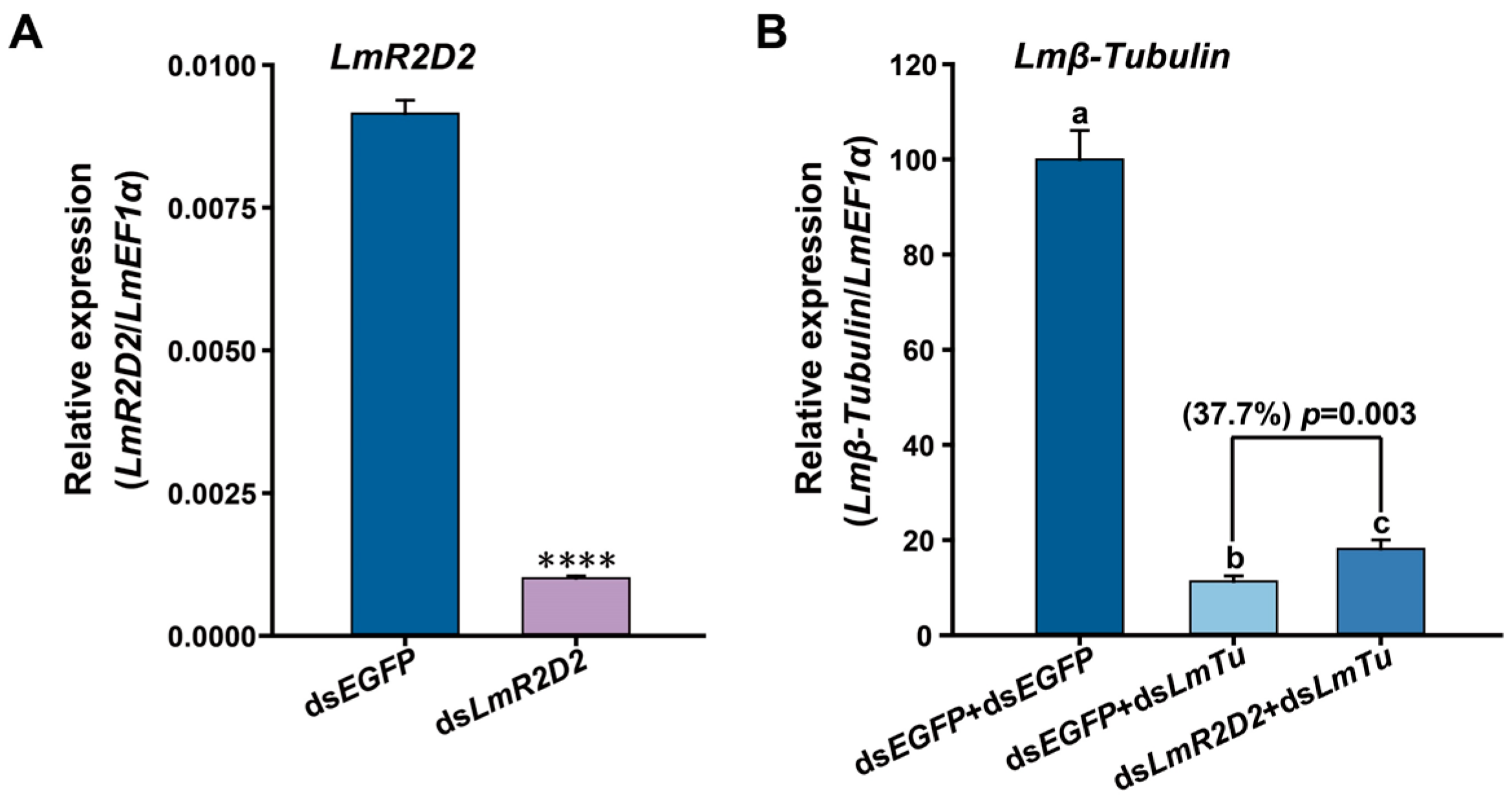
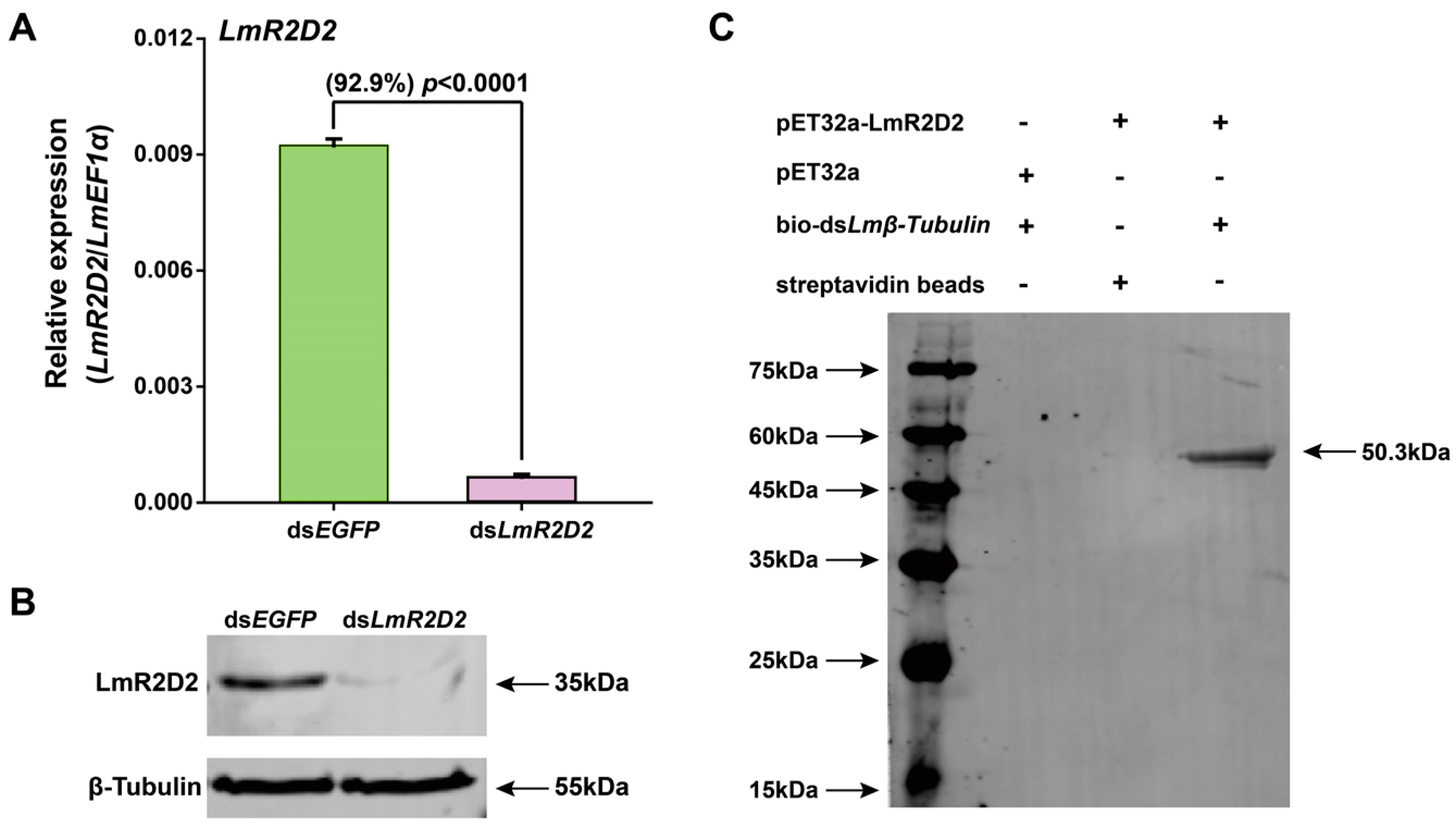
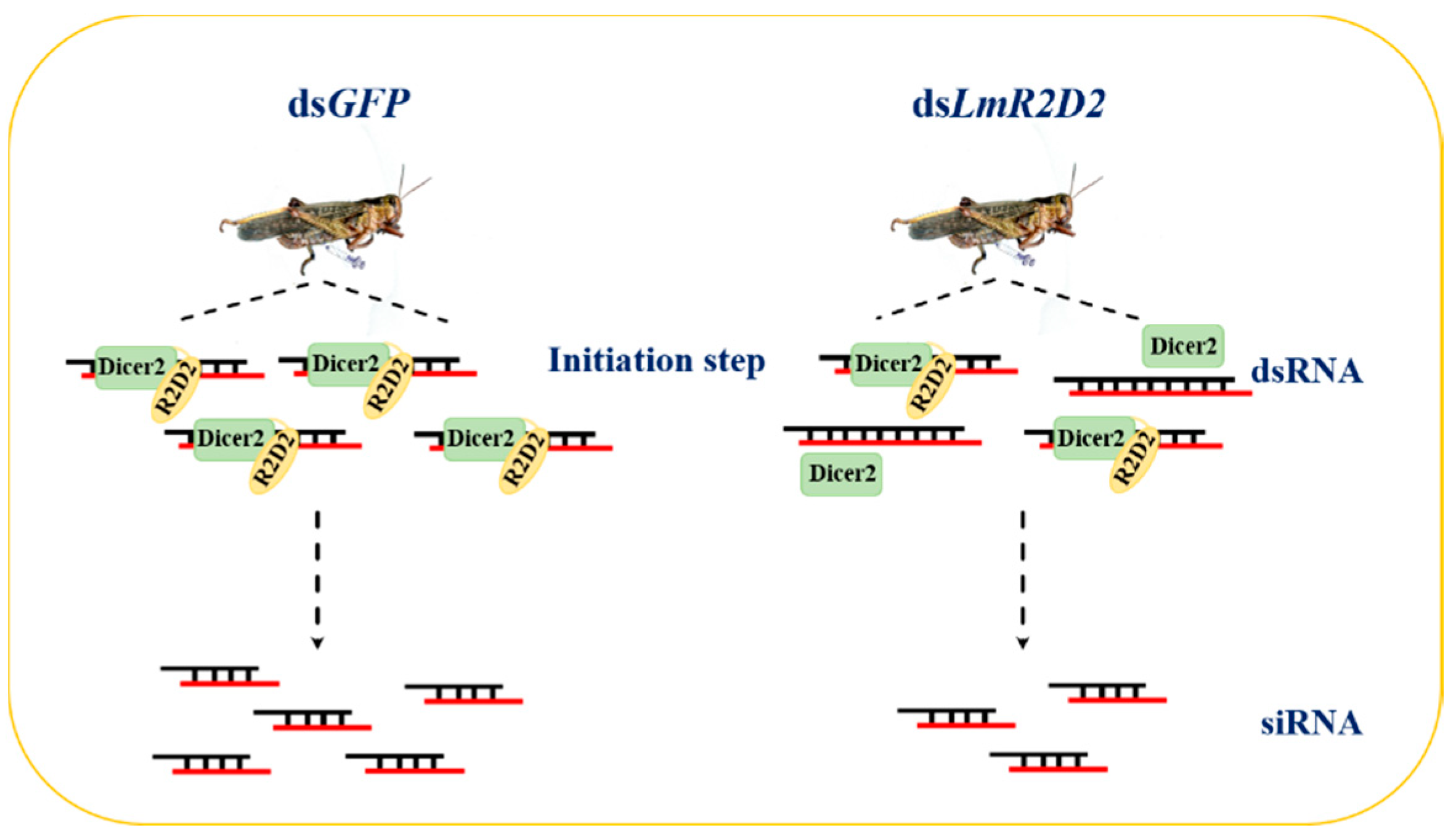
3.3. Effect of RNAi against LmR2D2 on RNAi Efficiency
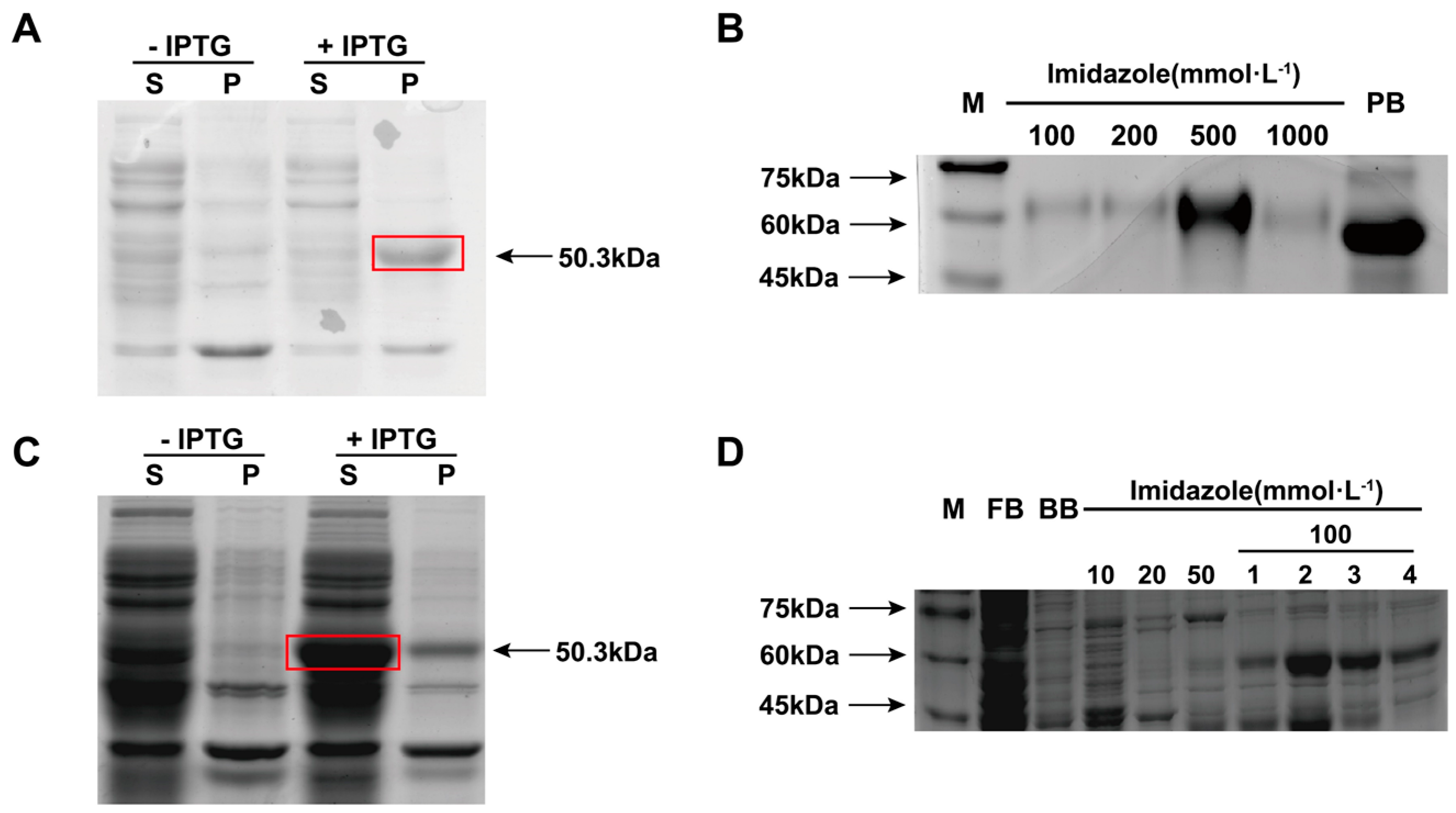
3.4. Expression and Purification of Recombinant LmR2D2 Protein
3.5. Effect of RNAi Targeting LmR2D2 on Its mRNA and Protein Levels
3.6. Binding of LmR2D2 Protein to dsRNA In Vitro
4. Discussion
4.1. Molecular Characterization of R2D2
4.2. Involvement of LmR2D2 in siRNA Pathway
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Burand, J.P.; Hunter, W.B. RNAi: Future in insect management. J. Invertebr. Pathol. 2013, 112, S68–S74. [Google Scholar] [CrossRef]
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef]
- Andrade, E.C.; Hunter, W.B.; Abdurakhmonov, I.Y. (Eds.) RNA Interference—Natural Gene-Based Technology for Highly Specific Pest Control (HiSPeC) in RNA Interference; InTech: Rijeka, Croatia, 2016; pp. 391–409. [Google Scholar]
- Belles, X. Beyond Drosophila: RNAi in vivo and functional genomics in insects. Annu. Rev. Entomol. 2010, 55, 111–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.-L.; Barthel, A.; et al. RNA interference in Lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, A.M.; Silver, K.; Zhang, J.; Park, Y.; Zhu, K.Y. Molecular mechanisms influencing efficiency of RNA interference in insects. Pest Manag. Sci. 2019, 75, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, D.; Pauli, T.; Donath, A.; Meusemann, K.; Podsiadlowski, L.; Petersen, M.; Peters, R.S.; Mayer, C.; Liu, S.; Zhou, X.; et al. Phylogenetic origin and diversification of RNAi pathway genes in insects. Genome Biol. Evol. 2016, 8, 3784–3793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obbard, D.J.; Gordon, K.H.J.; Buck, A.H.; Jiggins, F.M. The evolution of RNAi as a defence against viruses and transposable elements. Philos. Trans. R. Soc. B 2008, 364, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.Y.; Palli, S.R. Mechanisms, applications, and challenges of insect RNA interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [Green Version]
- Elbashir, S.M.; Lendeckel, W.; Tuschl, T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev. 2001, 15, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Siomi, H.; Siomi, M.C. On the road to reading the RNA-interference code. Nature 2009, 457, 396–404. [Google Scholar] [CrossRef]
- Zamore, P.D.; Tuschl, T.; Sharp, P.A.; Bartel, D.P. RNAi: Double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals. Cell 2000, 101, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Jiang, F.; Kalidas, S.; Smith, D.; Liu, Q. Dicer-2 and R2D2 coordinately bind siRNA to promote assembly of the siRISC complexes. RNA 2006, 12, 1514–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomari, Y.; Du, T.; Zamore, P.D. Sorting of Drosophila small silencing RNAs. Cell 2007, 130, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipowicz, W. RNAi: The nuts and bolts of the RISC machine. Cell 2005, 122, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Mello, C.C.; Conte, D.J. Revealing the world of RNA interference. Nature 2004, 431, 338–342. [Google Scholar] [CrossRef]
- Liu, Q.; Rand, T.A.; Kalidas, S.; Du, F.; Kim, H.E.; Smith, D.P.; Wang, X. R2D2, a bridge between the initiation and effector steps of the Drosophila RNAi pathway. Science 2003, 301, 1921–1925. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, S.K.; Zhu, L.; Fukunaga, R. The C-terminal dsRNA-binding domain of Drosophila Dicer-2 is crucial for efficient and high-fidelity production of siRNA and loading of siRNA to Argonaute2. RNA 2017, 23, 1139–1153. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.S.; Shukla, J.N.; Gong, Z.J.; Mogilicherla, K.; Palli, S.R. RNA interference in the Colorado potato beetle, Leptinotarsa decemlineata: Identification of key contributors. Insect Biochem. Mol. Biol. 2016, 78, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Swevers, L.; Liu, J.; Huvenne, H.; Smagghe, G. Search for limiting factors in the RNAi pathway in silkmoth tissues and the Bm5 cell line: The RNA-binding proteins R2D2 and Translin. PLoS ONE 2011, 6, e20250. [Google Scholar] [CrossRef] [Green Version]
- Kolliopoulou, A.; Swevers, L. Functional analysis of the RNAi response in ovary-derived silkmoth Bm5 cells. Insect Biochem. Mol. Biol. 2013, 43, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, X.; Ma, Z.; Xue, L.; Han, J.; Yu, D.; Kang, L. CSP and takeout genes modulate the switch between attraction and repulsion during behavioral phase change in the migratory locust. PLoS Genet. 2011, 7, e1001291. [Google Scholar] [CrossRef]
- Ma, E.B.; He, Y.P.; Zhu, K.Y. Comparative studies of acetylcholinesterases purified from two field populations of the oriental migratory locust (Locusta migratoria manilensis): Implications of insecticide resistance. Pestic. Biochem. Physiol. 2004, 78, 67–77. [Google Scholar] [CrossRef]
- Yang, M.L.; Zhang, J.Z.; Zhu, K.Y.; Xuan, T.; Liu, X.J.; Guo, Y.P.; Ma, E.B. Mechanisms of organophosphate resistance in a field population of oriental migratory locust, Locusta migratoria manilensis (Meyen). Arch. Insect Biochem. Physiol. 2009, 71, 3–15. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [Green Version]
- Tabara, H.; Yigit, E.; Siomi, H.; Mello, C.C. The dsRNA binding protein RDE-4 interacts with RDE-1, DCR-1, and a DExH-box helicase to direct RNAi in C. elegans. Cell 2002, 109, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Li, Z.; Cao, J.; Zhang, S.; Zhang, H.; Wu, X.; Zhang, Q.; Liu, X. Selection and assessment of reference genes for quantitative PCR normalization in migratory locust Locusta migratoria (Orthoptera: Acrididae). PLoS ONE 2014, 9, e98164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Kalidas, S.; Sanders, C.; Ye, X.; Strauss, T.; Kuhn, M.; Liu, Q.; Smith, D.P. Drosophila R2D2 mediates follicle formation in somatic tissues through interactions with Dicer-1. Mech. Dev. 2008, 125, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application of Primers | Primer Names | Primer Sequence (5′–3′) | Product (bp) |
|---|---|---|---|
| Full-length verification | LmR2D2-F | ACATGAATCAGAAGACACCAGTGTCAGT | 954 bp |
| LmR2D2-R | TTCATCCTGGACGCCTTCCTGATGAGAG | ||
| RT-qPCR Analysis | LmR2D2 RT-F | TCCGTTGGTTTGCTGATTGA | 103 bp |
| LmR2D2 RT-R | TGGTGAACTGCTTGGCGTGT | ||
| EF1α RT-F | AGCCCAGGAGATGGGTAAAG | 155 bp | |
| EF1α RT-R | CTCTGTGGCCTGGAGCATC | ||
| Lmβ-Tubulin RT-F | GAAATGGAGTTCACGGAAGC | 109 bp | |
| Lmβ-Tubulin RT-R | CTTGCTCCTCATCAAACTCG | ||
| dsRNA synthesis | EGFP T7-F | taatacgactcactatagggGACGTAAACGGCCACAAGTT | 496 bp |
| EGFP T7-R | taatacgactcactatagggTGTTCTGCTGGTAGTGGTCG | ||
| LmR2D2 T7-F | taatacgactcactatagggATCAAAAGATTGCCACAGGC | 493 bp | |
| LmR2D2 T7-R | taatacgactcactatagggGCATTCTGCATAGCCTCCTC | ||
| Lmβ-Tubulin T7-F | taatacgactcactatagggAGGCCACTACACAGAGGGTG | 401 bp | |
| Lmβ-Tubulin T7-R | taatacgactcactatagggTGACGCCAGACATGGTAAGA |
| Species | Gene Name | GenBank Accession No. |
|---|---|---|
| Aphis glycines | AgR2D2 | JX870426.1 |
| Nilaparvatalugens | NlR2D2 | KC316044.1 |
| Bemisiatabaci | BtR2D2 | KF192312.1 |
| Locusta migratoria | LmR2D2 | MZ313537 |
| Bombyx mori | BmR2D2 | AB566385.1 |
| Spodoptera litura | SlR2D2 | KF717086.1 |
| Agrilusplanipennis | ApR2D2 | KP036494.1 |
| Triboliumcastaneum | TcR2D2 | NM_001134953.1 |
| Drosophila melanogaster | DmR2D2 | NM_135308.2 |
| Aedes aegypti | AaR2D2 | KJ598053.1 |
| Culex quinquefasciatus | CqR2D2 | XM_001861911.1 |
| Mayetiola destructor | R2D2a | AFX89026.1 |
| Mayetiola destructor | R2D2b | AFX89027.1 |
| Diabrotica virgifera virgifera | R2D2a | XP_028140225.1 |
| Diabrotica virgifera virgifera | R2D2b | XP_028148994.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, L.; Wang, Y.; Abbas, M.; Zhang, T.; Ma, E.; Xing, S.; Zhu, K.Y.; Zhang, J. Molecular Characterizations and Functional Analyses of LmR2D2 in the Locusta migratoria siRNA Pathway. Insects 2021, 12, 812. https://doi.org/10.3390/insects12090812
Gao L, Wang Y, Abbas M, Zhang T, Ma E, Xing S, Zhu KY, Zhang J. Molecular Characterizations and Functional Analyses of LmR2D2 in the Locusta migratoria siRNA Pathway. Insects. 2021; 12(9):812. https://doi.org/10.3390/insects12090812
Chicago/Turabian StyleGao, Lu, Yanli Wang, Mureed Abbas, Tingting Zhang, Enbo Ma, Shuping Xing, Kun Yan Zhu, and Jianzhen Zhang. 2021. "Molecular Characterizations and Functional Analyses of LmR2D2 in the Locusta migratoria siRNA Pathway" Insects 12, no. 9: 812. https://doi.org/10.3390/insects12090812