
The Importance of Time and Place: Nutrient Composition and Utilization of Seasonal Pollens by European Honey Bees (Apis mellifera L.)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. General Overview
2.2. Pollen Identification
2.3. Pollen Protein and Amino Acid Analyses
2.4. Pollen Lipid and Fatty Acid Analyses
2.5. Cage Setup, Pollen Feeding and Sampling
2.6. Estimating Pollen Protein Digestion
2.7. Hemolymph Protein Concentration
2.8. Measuring Hypopharyngeal Glands
2.9. Expression of vg, hex 70, and hex 110
2.10. Statistical Analysis
3. Results
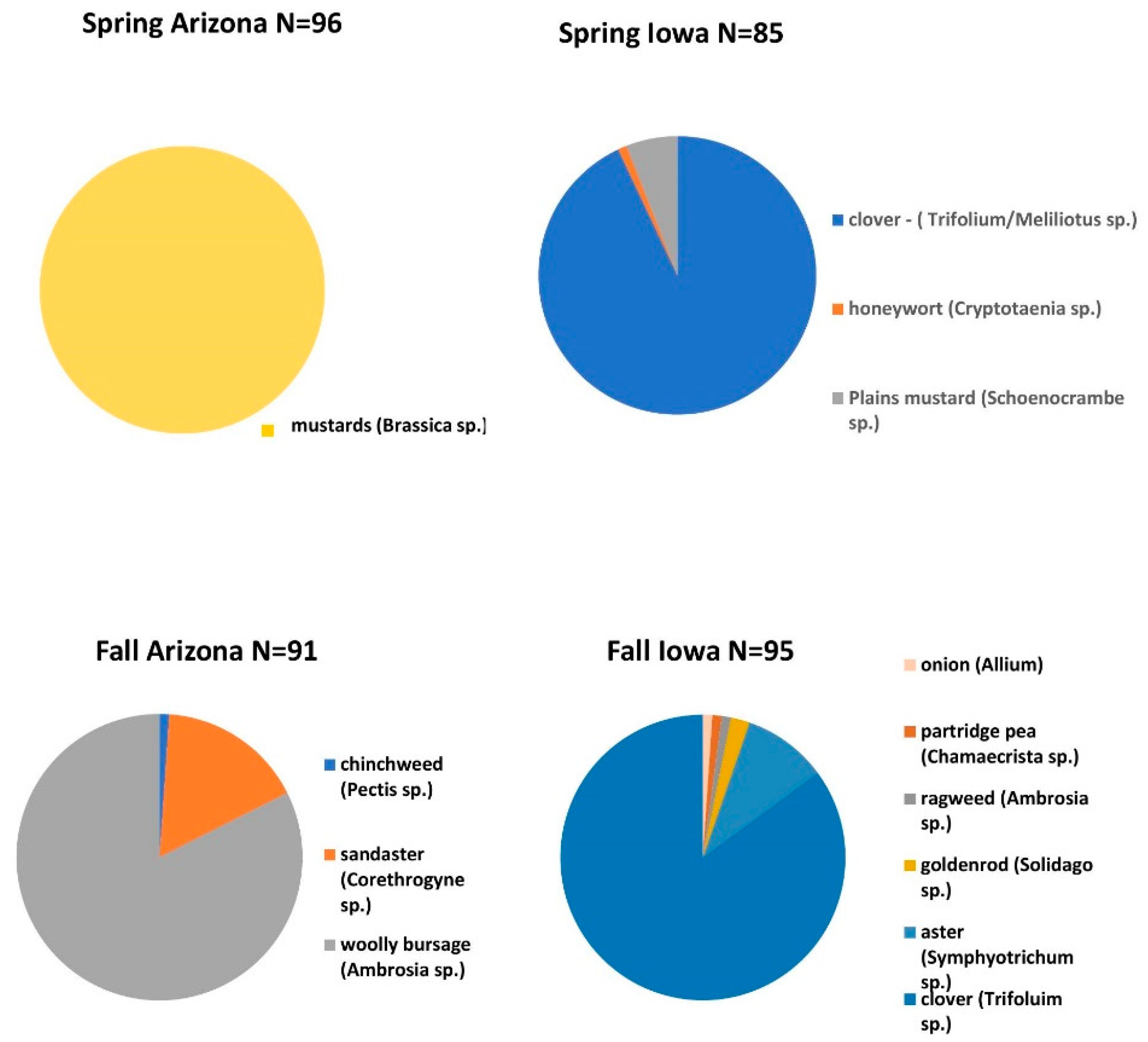
3.1. Floral Composition of Pollens
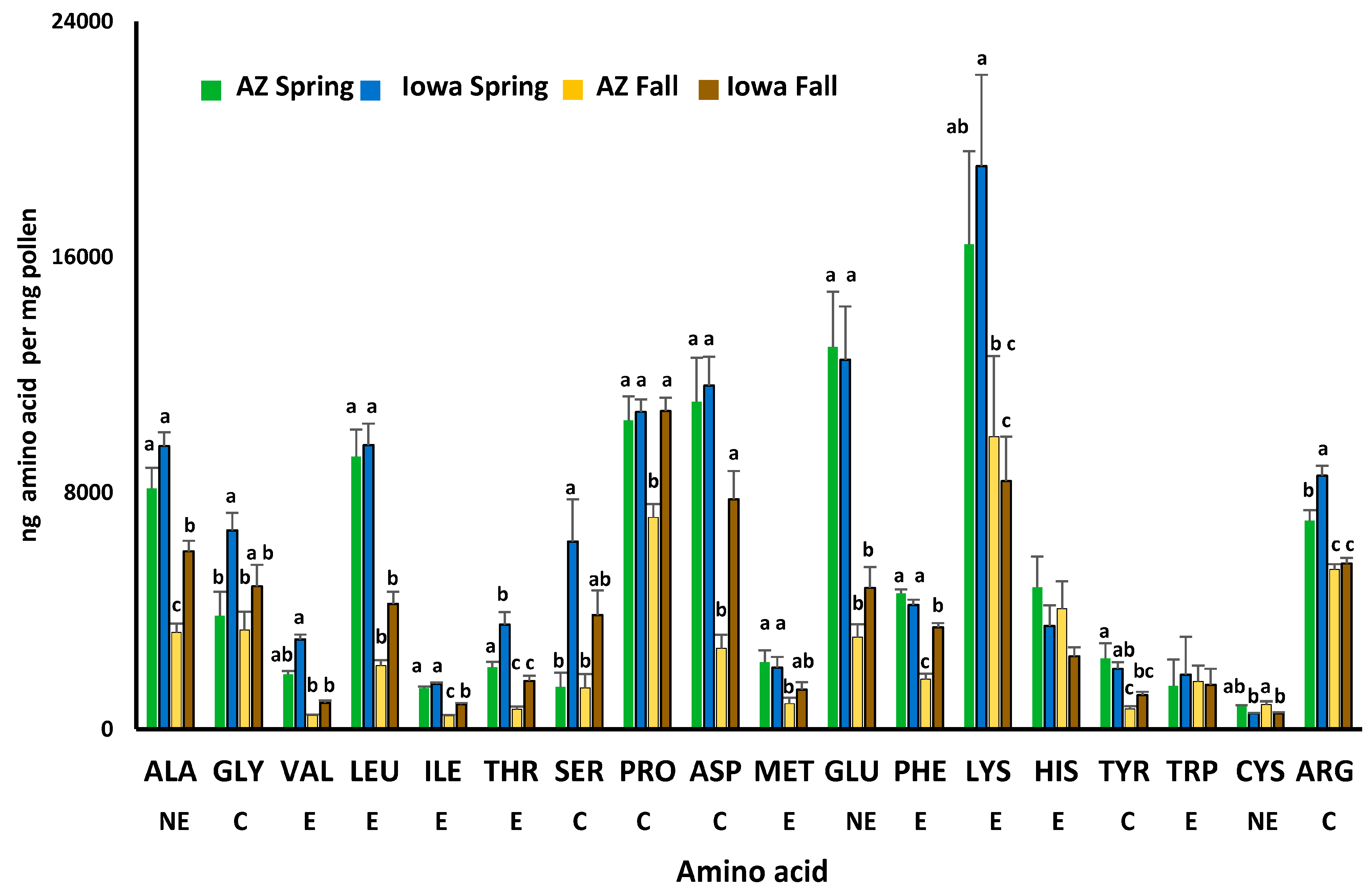
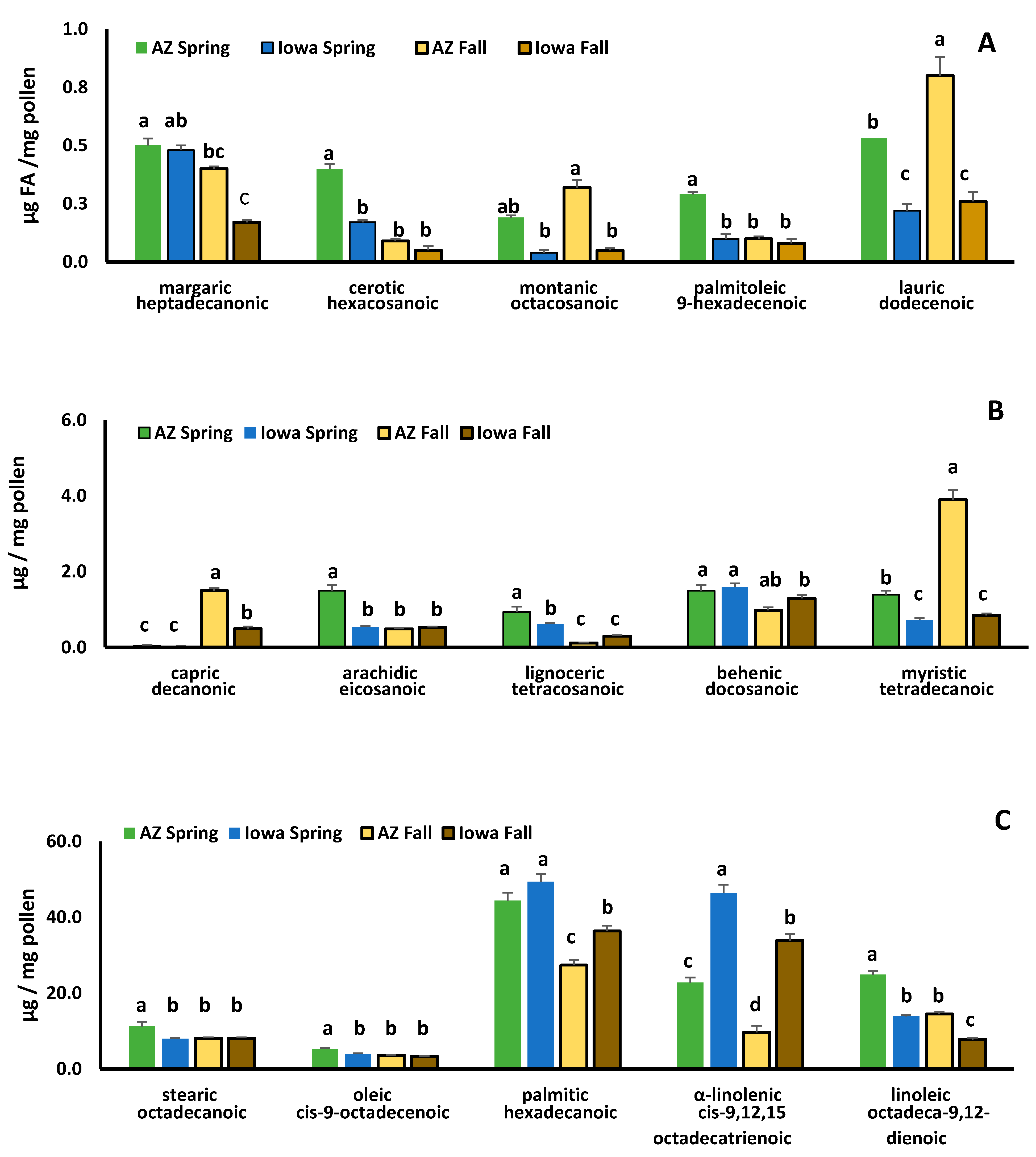
3.2. Comparisons of Protein, Lipids, Amino and Fatty Acids in Spring and Fall Pollens
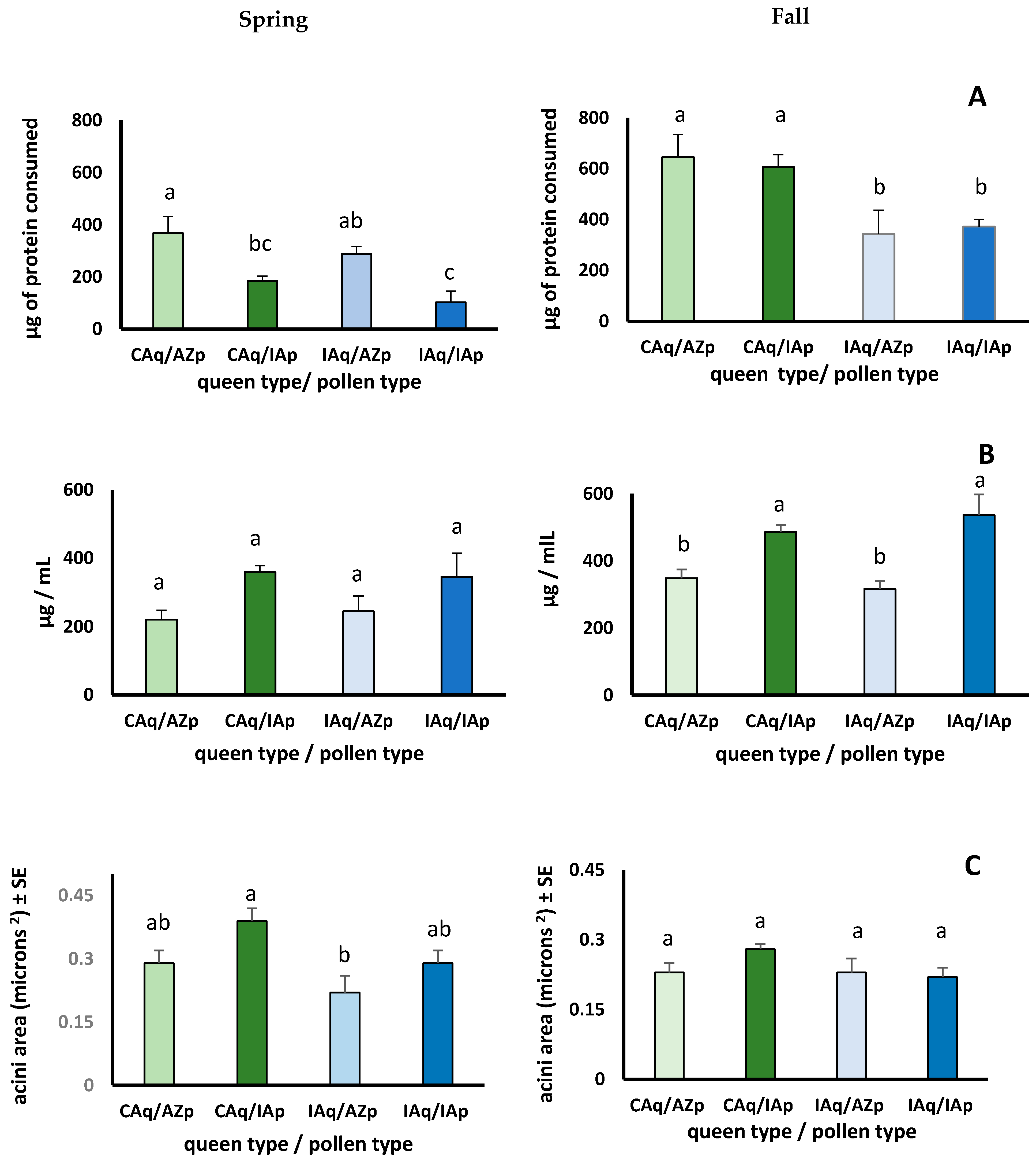
3.3. Seasonal Pollen Consumption, Protein Digestion, Protein Consumption, Hemolymph Protein Concentrations, and Hypopharyngeal Gland Size
3.3.1. Do Workers Respond Differently to Pollen Mixes That Differ in Composition?
3.3.2. Do Workers Respond Differently to Pollen Based on Season?
3.3.3. Do Responses to Season and Pollen Sources Differ between Queen Lines?
3.3.4. Expression of hex 70a, hex 110 and vg
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Solon-Biet, S.M.; Walters, K.A.; Simanainen, U.K.; McMahon, A.C.; Ruohonen, K.; Ballard, J.W.O.; Raubenheimer, D.; Handelsman, D.J.; Le Couteur, D.G.; Simpson, S.J. Macronutrient balance, reproductive function, and lifespan in aging mice. Proc. Natl. Acad. Sci. USA 2015, 112, 3481–3486. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Speakman, J.R.; Wu, Q.; Zhang, C.; Hu, Y.; Xia, M.; Hambly, C.; Wang, L.; Wei, W.; Zhang, J.; et al. Exceptionally low daily energy expenditure in the bamboo-eating giant panda. Science 2015, 349, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Sword, G.A.; Lorch, P.D.; Couzin, I.D. Cannibal crickets on a forced march for protein and salt. Proc. Natl. Acad. Sci. USA 2006, 103, 4152–4156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machovsky-Capuska, G.E.; Senior, A.M.; Zantis, S.P.; Barna, K.; Cowieson, A.J.; Pandya, S.; Pavard, C.; Shiels, M.; Raubenheimer, D. Dietary protein selection in a free-ranging urban population of common myna birds. Behav. Ecol. 2016, 27, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Baker, H.G. Non-sugar chemical constituents of nectar. Apidologie 1977, 8, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Crailsheim, K.; Schneider, L.H.W.; Hrassnigg, N.; Bühlmann, G.; Brosch, U.; Gmeinbauer, R.; Schöffmann, B. Pollen consumption and utilization in worker honeybees (Apis mellifera carnica): Dependence on individual age and function. J. Insect Physiol. 1992, 38, 409–419. [Google Scholar] [CrossRef]
- Crailsheim, K. The protein balance of the honey bee worker. Apidologie 1990, 21, 417–429. [Google Scholar] [CrossRef]
- Crailsheim, K.; Hrassnigg, N.; Gmeinbauer, R.; Szolderits, M.J.; Schneider, L.H.W. Pollen utilization in non-breeding honeybees in winter. J. Insect Physiol. 1993, 39, 369–373. [Google Scholar] [CrossRef]
- Fahrenholz, L.; Lamprecht, I.; Schricker, B. Thermal investigations of a honey bee colony: Thermoregulation of the hive during summer and winter and heat production of members of different bee castes. J. Comp. Physiol. B 1989, 159, 551–560. [Google Scholar] [CrossRef]
- Simpson, J.S. Nest climate regulation in honey bee colonies. Science 1961, 133, 1327–1334. [Google Scholar] [CrossRef]
- Stabentheiner, A.; Kovac, H.; Brodschneider, R. Honeybee colony thermoregulation—Regulatory mechanisms and contribution of individuals in dependence on age, location and thermal stress. PLoS ONE 2010, 5, e8967. [Google Scholar] [CrossRef] [PubMed]
- Fluri, P.; Lüscher, M.; Wille, H.; Gerig, L. Changes in weight of the pharyngeal gland and haemolymph titres of juvenile hormone, protein and vitellogenin in worker honey bees. J. Insect Physiol. 1982, 28, 61–68. [Google Scholar] [CrossRef]
- Amdam, G.; Omholt, S.W. The regulatory anatomy of honeybee lifespan. J. Theor. Biol. 2002, 216, 209–228. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.Y.; Robinson, G.E. Seasonal changes in juvenile hormone titers and rates of biosynthesis in honey bees. J. Comp. Physiol. B 1995, 165, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Fluri, P.; Wille, H.; Gerig, L.; Lüscher, M. Juvenile hormone, vitellogenin and haemocyte composition in winter worker honeybees (Apis mellifera). Experientia 1977, 33, 1240–1241. [Google Scholar] [CrossRef]
- Münch, D.; Kreibich, C.D.; Amdam, G.V. Aging and its modulation in a long-lived worker caste of the honey bee. J. Exper. Biol. 2013, 216, 1638–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Zheng-Guo, L.; Peng, Q.; Zhang, W.; Chen, X.; Fan, R.; Yin, L.; Ji, T. Investigating the regulation of hypopharnygeal gland activity in honey bees (Apis mellifera L.) under overwintering conditions via morphologic analysis combined with iTRAQ-based comparative proteomics. Ann. Entomol. Soc. Am. 2018, 111, 127–135. [Google Scholar] [CrossRef]
- De Grandi-Hoffman, G.; Gage, S.L.; Corby-Harris, V.; Carroll, M.; Chambers, M.; Graham, H.; de Jong, E.W.; Hidalgo, G.; Calle, S.; Azzouz-Olden, F.; et al. Connecting the nutrient composition of seasonal pollens with changing nutritional needs of honey bee (Apis mellifera L.) colonies. J. Insect Physiol. 2018, 109, 114–124. [Google Scholar] [CrossRef]
- Reznick, D.N.; Ghalambor, C.K. The population ecology of contemporary adaptations: What empirical studies reveal about the conditions that promote adaptive evolution. In Microevolution Rate, Pattern, Process. Contemporary Issues in Genetics and Evolution; Hendry, A.P., Kinnison, M.T., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 8, pp. 183–198. [Google Scholar] [CrossRef]
- Büchler, R.; Costa, C.; Hatjina, F.; Andonov, S.; Marina, D.; LeConte, Y.; Uzunov, A.; Berg, S.; Bienkowska, M.; Bouga, M.; et al. The influence of genetic origin and its interaction with environmental effects on the survival of Apis mellifera L. colonies in Europe. J. Apic. Res. 2014, 53, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Hatjina, F.; Costa, C.; Büchler, R.; Uzunov, A.; Drazic, M.; Filipi, J.; Charistos, L.; Ruottinen, L.; Andonov, S.; Meixner, M.D.; et al. Population dynamics of European honey bee genotypes under different environmental conditions. J. Apic. Res. 2014, 53, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Döke, M.A.; McGrady, C.M.; Otieno, M.; Grozinger, C.M.; Frazier, M. Colony size, rather than geographic origin of stocks predicts overwintering success in honey bees (Hymenoptera: Apidae) in the Northeastern United States. J. Econ. Entomol. 2019, 112, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.; Melathopoulos, A.P.; White, R.; Pernal, S.F.; Guarna, M.M.; Foster, L.J. Ecological adaptation of diverse honey bee (Apis mellifera) populations. PLoS ONE 2010, 5, e11096. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.R.; Nunes, F.M.; Cristino, A.S.; Simões, Z.L.; Bitondi, M.M. The four hexamerin genes in the honey bee: Structure, molecular evolution and function deduced from expression patterns in queens, workers and drones. BMC Mol. Biol. 2010, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.R.; Nunes, F.M.F.; Simões, Z.L.P.; Bitondi, M.M.G. A honeybee storage protein gene, hex 70a, expressed in developing gonads and nutritionally regulated in adult fat body. J. Insect Physiol. 2008, 54, 867–877. [Google Scholar] [CrossRef] [PubMed]
- DeGrandi-Hoffman, G.; Watkins, J.C.; Collins, A.M.; Loper, G.M.; Martin, J.H.; Arias, M.C.; Sheppard, W.S. Queen developmental time as a factor in the Africanization of European honey bee (Hymenoptera: Apidae) populations. Ann. Entomol. Soc. Am. 1998, 91, 52–58. [Google Scholar] [CrossRef]
- Little, D.P.; Schwarzbach, A.E.; Adams, R.P.; Hsieh, C.F. The circumscription and phylogenetic relationships of Callitropsis and the newly described genus Xanthocyparis (Cupressaceae). Am. J. Bot. 2004, 91, 1872–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y.; de Jong, E.W.; Chambers, M.L.; Hidalgo, G. Effects of oral exposure to fungicides on honey bee nutrition and virus levels. J. Econ. Entomol. 2015, 108, 2518–2528. [Google Scholar] [CrossRef] [Green Version]
- Hušek, P.; Šimek, P.; Hartvich, P.; Zahradníčková, H. Fluoroalkyl chloroformates in treating amino acids for gas chromatographic analysis. J. Chromatogr. A 2008, 1186, 391–400. [Google Scholar] [CrossRef]
- Yust, M.M.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F.; Alaiz, M. Determination of tryptophan by high-performance liquid chromatography of alkaline hydrolysates with spectrophotometric detection. Food Chem. 2004, 85, 317–320. [Google Scholar] [CrossRef]
- Manneberg, M.; Lahm, H.; Fountoulakis, M. Quantification of Cysteine Residues Following Oxidation to Cysteic Acid in the Presence of Sodium Azide. Anal. Biochem. 1995, 231, 349–353. [Google Scholar] [CrossRef]
- Amenta, J.S. A rapid extraction and quantification of total lipids and lipid fractionsin blood and feces. Clin. Chem. 1970, 16, 339–346. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y.; Huang, E.; Huang, M.H. The effect of diet on protein concentration, hypopharyngeal gland development and virus load in worker honey bees (Apis mellifera L.). J. Insect Physiol. 2010, 56, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- DeGrandi-Hoffman, G.; Chen, Y.; Rivera, R.; Carroll, M.; Chambers, M.; Hidalgo, G.; de Jong, E.W. Honey bee colonies provided with natural forage have lower pathogen loads and higher overwinter survival than those fed protein supplements. Apidologie 2016, 47, 186–196. [Google Scholar] [CrossRef] [Green Version]
- Corby-Harris, V.; Meador, C.A.D.; Snyder, L.A.; Schwan, M.R.; Maes, P.; Jones, B.M.; Walton, A.; Anderson, K.E. Transcriptional, translational, and physiological signatures of undernourished honey bees (Apis mellifera) suggest a role for hormonal factors in hypopharyngeal gland degradation. J. Insect Physiol. 2016, 85, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 ΔΔCt method. Methods 2001, 408, 402–408. [Google Scholar] [CrossRef]
- Calflora. Cenchrus Longispinus (Hack.) Fernald. Available online: https://www.calflora.org/cgi-bin/species_query.cgi?where-calrecnum=1145 (accessed on 4 August 2020).
- Calflora. Ambrosia Psilostachya DC. Available online: https://www.calflora.org/cgi-bin/species_query.cgi?where-calrecnum=298 (accessed on 4 August 2020).
- Bilikova, K.; Hanes, J.; Saenger, W.; Klaudiny, J.; Šimúth, J. Apisimin, a new serine-valine-rich peptide from honeybee (Apis mellifera L.) royal jelly: Purification and molecular characterization. FEBS Lett. 2002, 528, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Briand, L.; Swasdipan, N.; Nespoulous, C.; Bézirard, V.; Blon, F.; Huet, J.C.; Ebert, P.; Pernollet, J.-C. Characterization of a chemosensory protein (ASP3c) from honeybee (Apis mellifera L.) as a brood pheromone carrier. Eur. J. Biochem. 2002, 269, 4586–4596. [Google Scholar] [CrossRef]
- Ma, L.; Wang, Y.; Zhang, W.; Wang, H.; Liu, Z.; Xu, B. Alterations in protein and amino acid metabolism in honeybees (Apis mellifera) fed different L -leucine diets during the larval stage. J. Asia Pac. Entomol. 2016, 19, 769–774. [Google Scholar] [CrossRef]
- Münch, D.; Amdam, G.V. The curious case of aging plasticity in honey bees. FEBS Lett. 2010, 584, 2496–2503. [Google Scholar] [CrossRef] [Green Version]
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in honey bees: Digital gene expression analysis of pollen’s nutritive effects on healthy and varroa-parasitized bees. BMC Genom. 2011, 12, 496. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Sun, Z.; Zhang, J.; Kang, K.; Chen, J.; Zhang, W. Activation of the TOR signalling pathway by glutamine regulates insect fecundity. Sci. Rep. 2015, 5, 10694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teulier, L.; Weber, J.; Crevier, J.; Darveau, C. Proline as a fuel for insect flight: Enhancing carbohydrate oxidation in hymenopterans. Proc. Biol. Sci. 2016, 283, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Standifer, L.N. A comparison of the protein quality of pollens for growth-stimulation of the hypopharyngeal glands and longevity of honey bees, Apis mellifera L. (Hymenoptera: Apidae). Insectes Sociaux 1967, 14, 415–425. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arien, Y.; Dag, A.; Zarchin, S.; Masci, T.; Shafir, S. Omega-3 deficiency impairs honey bee learning. Proc. Natl. Acad. Sci. USA 2015, 112, 201517375. [Google Scholar] [CrossRef] [Green Version]
- Arien, Y.; Dag, A.; Yona, S.; Tietel, Z.; Lapidot, T.; Sha, S. Effect of diet lipids and omega-6: 3 ratio on honey bee brood development, adult survival and body composition. J. Insect Physiol. 2020, 124, 104074. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Allier, F.; Decourtye, A.; Odoux, J.-F.; Tamic, T.; Chabirand, M.; Delestra, E.; Decugis, F.; Le Conte, Y.; Henry, M. A ‘Landscape physiology’ approach for assessing bee health highlights the benefits of floral landscape enrichment and semi-natural habitats. Sci. Rep. 2017, 7, 40568. [Google Scholar] [CrossRef] [PubMed]
- Avni, D.; Hendriksma, H.P.; Dag, A.; Uni, Z.; Shafir, S. Nutritional aspects of honey bee-collected pollen and constraints on colony development in the eastern Mediterranean. J. Insect Physiol. 2014, 69, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, M.T.O.; Marquina, A.D.; Serrano, R.B.; Blazquez Abellán, G. The importance of bee-collected pollen in the diet: A study of its composition. Int. J. Food Sci. Nutr. 2002, 53, 217–224. [Google Scholar] [CrossRef]
- Manning, R. Fatty acids in pollen: A review of their importance for honey bees. Bee World 2001, 82, 60–75. [Google Scholar] [CrossRef]
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.K.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malcicka, M.; Visser, B.; Ellers, J. An evolutionary perspective on linoleic acid synthesis in animals. Evol. Biol. 2018, 45, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Snyder, L.; Meador, C.; Ayotte, T. Honey bee (Apis mellifera) nurses do not consume pollens based on their nutritional quality. PLoS ONE 2018, 13, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.M.; Awmack, C.S.; Murray, D.A.; Williams, I.H. Are honey bees’ foraging preferences affected by pollen amino acid composition? Ecol. Entomol. 2003, 28, 622–627. [Google Scholar] [CrossRef]
- Ruedenauer, F.A.; Spaethe, J.; Leonhardt, S.D. Hungry for quality—individual bumblebees forage flexibly to collect high-quality pollen. Behav. Ecol. Sociobiol. 2016, 70, 1209–1217. [Google Scholar] [CrossRef]
- Sheppard, W.S. Honey Bee Diversity—Races, Ecotypes, Strains. In The Hive and the Honey Bee; Graham, G.M., Ed.; Dadant & Sons: Hamilton, IL, USA, 2015; pp. 53–67. [Google Scholar]
- Ruttner, F. Biogeography and Taxonomy of Honey Bees; Springer: Berlin, Germany, 1988; pp. 1–284. [Google Scholar]
- Knoll, S.; Pinna, W.; Varcasia, A.; Scala, A.; Grazia, M. The honey bee (Apis mellifera L., 1758) and the seasonal adaptation of productions. Highlights on summer to winter transition and back to summer metabolic activity. A review. Livest. Sci. 2020, 235, 104011. [Google Scholar] [CrossRef]
- Traynor, K.S.; Wang, Y.; Brent, C.S.; Amdam, G.V.; Page, R.E. Young and old honeybee (Apis mellifera) larvae differentially prime the developmental maturation of their caregivers. Anim. Behav. 2017, 124, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Mattila, H.R.; Harris, J.L.; Otis, G.W. Timing of production of winter bees in honey bee (Apis mellifera) colonies. Insectes Soc. 2001, 48, 88–93. [Google Scholar] [CrossRef]
- Döke, M.A.; Frazier, M.; Grozinger, C.M. Overwintering honey bees: Biology and management. Curr. Opin. Insect Sci. 2015, 10, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Walton, A.; Dolezal, A.G.; Bakken, M.A.; Toth, A.L. Hungry for the queen: Honeybee nutritional environment affects worker pheromone response in a life stage—Dependent manner. Func. Ecol. 2018, 32, 2699–2706. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kaftanoglu, O.; Brent, C.S.; Page, R.E.; Amdam, G.V. Starvation stress during larval development facilitates an adaptive response in adult worker honey bees (Apis mellifera L.). J. Exper. Biol. 2016, 219, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations | Arizona Spring | Iowa Spring | Arizona Fall | Iowa Fall | F * | p |
|---|---|---|---|---|---|---|
| total soluble protein (µg/mg) | 353.7 ± 7.5 a | 275.8 ± 14.2 b | 368.8 ± 29.1 a | 379.4 ± 29.9 a | 5.28 | 0.015 |
| total amino acids (ng/mg) | 102.4 ± 9.0 a | 116.0 ± 0.1 a | 49.7 ± 6.0 b | 70.0 ± 4.8 b | 17.1 | <0.0001 |
| total lipids (µg/mg) | 127.2 ± 8.0 ab | 157.7 ± 21.8 a | 87.9 ± 12.8 b | 148.3 ± 12.5 a | 4.45 | 0.025 |
| total fatty acids (µg/mg) | 33.5 ± 1.7 a | 36.6 ± 1.4 a | 20.9 ± 1.0 c | 27.1 ± 1.1 b | 27.4 | <0.0001 |
| Protein to lipid ratio | 1:0.36 b | 1:0.57 a | 1:0.24 b | 1:0.39 ab | 8.36 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeGrandi-Hoffman, G.; Corby-Harris, V.; Carroll, M.; Toth, A.L.; Gage, S.; Watkins deJong, E.; Graham, H.; Chambers, M.; Meador, C.; Obernesser, B. The Importance of Time and Place: Nutrient Composition and Utilization of Seasonal Pollens by European Honey Bees (Apis mellifera L.). Insects 2021, 12, 235. https://doi.org/10.3390/insects12030235
DeGrandi-Hoffman G, Corby-Harris V, Carroll M, Toth AL, Gage S, Watkins deJong E, Graham H, Chambers M, Meador C, Obernesser B. The Importance of Time and Place: Nutrient Composition and Utilization of Seasonal Pollens by European Honey Bees (Apis mellifera L.). Insects. 2021; 12(3):235. https://doi.org/10.3390/insects12030235
Chicago/Turabian StyleDeGrandi-Hoffman, Gloria, Vanessa Corby-Harris, Mark Carroll, Amy L. Toth, Stephanie Gage, Emily Watkins deJong, Henry Graham, Mona Chambers, Charlotte Meador, and Bethany Obernesser. 2021. "The Importance of Time and Place: Nutrient Composition and Utilization of Seasonal Pollens by European Honey Bees (Apis mellifera L.)" Insects 12, no. 3: 235. https://doi.org/10.3390/insects12030235