
Physical Mapping of the Anopheles (Nyssorhynchus) darlingi Genomic Scaffolds

 , , , , ,
, , , , ,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Mosquitoes
2.2. Chromosome Preparation
2.3. Probes Preparation and Fluorescence in Situ Hybridization (FISH)
2.4. Physical Mapping and Probe Location Analysis
2.5. DNA Sequencing and Bioinformatics Analyses
3. Results
3.1. Probes and Corresponding Sequences in the Genome Assembly
3.2. Chromosome X
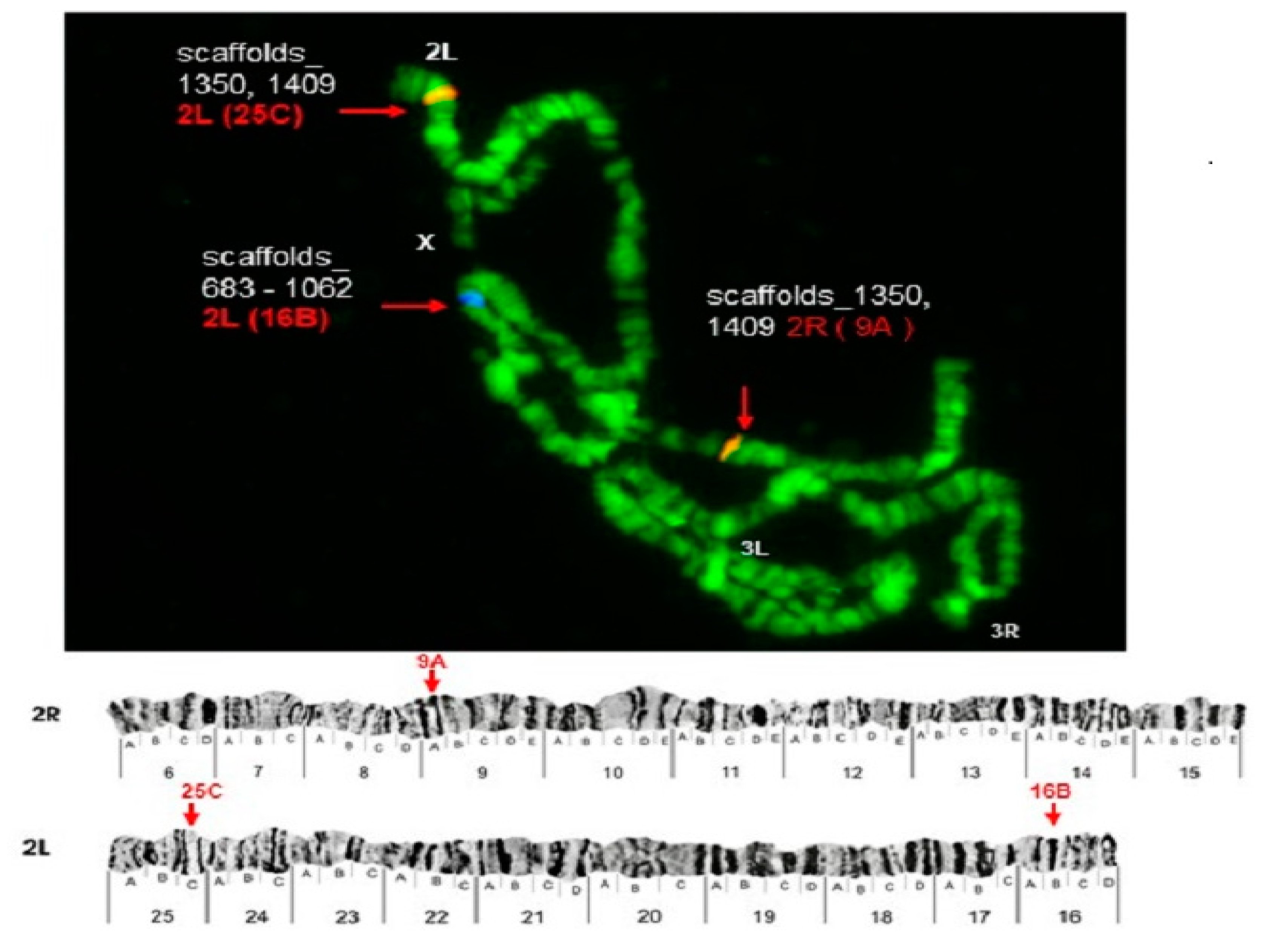
3.3. Chromosomes 2 and 3
3.4. Comparative Mapping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rachou, R.G. Anofelinos do Brasil. Comportamento das espécies vetoras de malaria. Rev. Bras. Malariol. Doenças Trop. 1958, 10, 145–181. [Google Scholar]
- Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. Principais Mosquitos de Importância Sanitária no Brasil, 1st ed.; Editora FIOCRUZ: Rio de Janeiro, Brasil, 1994; pp. 60–92. [Google Scholar]
- World Health Organization (WHO). World Malaria Report 2019, 3rd ed.; Global Malaria Programme-WHO: Geneva, Switzerland, 2019; p. 232. [Google Scholar]
- Harbach, R.E.; Kitching, I.J. The phylogeny of Anophelinae revisited: Inferences about the origin and classification of Anopheles (Diptera: Culicidae). Zool. Scr. 2016, 45, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Deane, L.M. Malaria vectors in Brazil. Mem. Inst. Oswaldo Cruz 1986, 81, 5–14. [Google Scholar] [CrossRef] [Green Version]
- De Arruda, M.; Carvalho, M.B.; Nussenzweig, R.S.; Maracic, M.; Ferreira, A.W.; Cochrane, A.H. Potential vectors of malaria and their different susceptibility to Plasmodium falciparum and Plasmodium vivax in Northern Brazil identified by immunoassay. Am. J. Trop. Med. Hyg. 1986, 5, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Tadei, W.P.; Thatcher, B.D.; Santos, J.M.; Scarpassa, V.M.; Rodrigues, I.B.; Rafael, M.S. Ecologic observations on anopheline vectors of malaria in the Brazilian Amazon. Am. J. Trop. Med. Hyg. 1998, 59, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Tadei, W.P.; Rodrigues, I.B.; Rafael, M.S.; Sampaio, R.T.M.; Mesquita, H.G.; Pinheiro, V.C.S.; Zequi, J.A.C.; Roque, R.A.; Santos, J.M.M. Adaptative processes, control measures, genetic background, and resilience of malaria vectors and environmental changes in the Amazon region. Hydrobiologia 2017, 789, 179–196. [Google Scholar] [CrossRef]
- Forattini, O.P. Compartamento exófilo de Anopheles darlingi Root, em região meridional do Brasil. Exophilic Behavior of Anopheles darlingi Root in a southern region of Brazil. Rev. Saude Publ. 1987, 21, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Komp, W.H.W. The occurrence of Anopheles darlingi Root in Central America. Am. J. Trop. Med. Hyg. 1941, 21, 659–670. [Google Scholar] [CrossRef]
- Marinotti, O.; Cerqueira, G.C.; de Almeida, L.G.P.; Ferro, M.I.T.; Loreto, E.L.S.; Zaha, A.; Teixeira, S.M. The genome of Anopheles darlingi, the main neotropical malaria vector. Nucleic Acids Res. 2013, 15, 7387–7400. [Google Scholar] [CrossRef]
- Malone, G.; Zimmer, P.D.; Meneghello, G.E.; Binneck, E.; Peske, S.T. Prospecção de genes em bibliotecas de cDNA. R. Bras. Agrociência 2006, 12, 7–13. [Google Scholar]
- Artemov, G.N.; Peery, A.N.; Jiang, X.; Tu, Z.; Stegniy, V.N.; Sharakhova, M.V.; Sharakhov, I.V. The physical genome mapping of Anopheles albimanus corrected scaffold misassemblies and identified interarm rearrangements in genus Anopheles. G3 (Bethesda) 2017, 7, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Rafael, M.S.; Tadei, W.P.; Recco-Pimentel, S.M. Location of ribosomal genes in the chromosomes of Anopheles darlingi and Anopheles nuneztovari (Diptera, Culicidae) from the brazilian Amazon. Mem. Inst. Oswaldo Cruz 2003, 98, 629–635. [Google Scholar] [CrossRef]
- Rafael, M.S.; Tadei, W.P.; Hunter, F.F. The physical gene Hsp70 map on polytene chromosomes of Anopheles darlingi from the Brazilian Amazon. Genetica 2004, 12, 89–94. [Google Scholar] [CrossRef]
- Bridi, L.C.; Sharakhova, M.V.; Sharakhov, I.V.; Cordeiro, J.; Azevedo Junior, G.M.; Tadei, W.P.; Rafael, M.S. Chromosomal localization of actin genes in the malaria mosquito Anopheles darlingi. Med. Vet. Entomol. 2013, 27, 118–121. [Google Scholar] [CrossRef] [Green Version]
- Granjeiro, W.G.; (National Institute of Amazon Research/INPA, Scientific Initiation Program/PIBIC, Manaus, Brazil). Personal communication, 2013.
- Bridi, L.C.; Rafael, M.S. GNBP domain of Anopheles darlingi: Are polymorphic inversions and gene variation related to adaptive evolution? Genetica 2016, 144, 99–106. [Google Scholar] [CrossRef]
- Rafael, M.S.; Rohde, C.; Bridi, L.C.; Valente, V.G.L.; Tadei, W.P. Salivary polytene chromosome map of Anopheles darlingi, the main vector of neotropical malaria. Am. J. Trop. Med. Hyg. 2010, 83, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Forattini, O.P. Entomologia Médica, 1st ed.; Editora da Universidade de São Paulo: São Paulo, Brazil, 1962; pp. 321–380. [Google Scholar]
- Faran, M.E.; Linthicum, K.J. A handbook of the Amazonian species of Anopheles (Nyssorhynchus) (Diptera: Culicidae). Mosq. Syst. 1981, 13, 1–91. [Google Scholar]
- French, W.L.; Baker, R.H.; Kitzmiller, J.B. Preparation of mosquito chromosomes. Mosq. News 1962, 22, 377–383. [Google Scholar]
- Kumar, V.; Collins, F.H. A technique for nucleic acid in situ hybridization to polytene chromosome of mosquitoes in the Anopheles gambiae complex. Insect Mol. Biol. 1994, 3, 41–47. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.R.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; p. 1546. [Google Scholar]
- Cornel, A.J.; Brisco, K.K.; Tadei, W.P.; Secundino, N.F.; Rafael, M.S.; Galardo, A.K.; Medeiros, J.F.; Pessoa, F.A.; Ríos-Velásquez, C.M.; Lee, Y.; et al. Anopheles darlingi polytene chromosomes: Revised maps including newly described inversions and evidence for population structure in Manaus. Mem. Inst. Oswaldo Cruz 2016, 111, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo-Calderon, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; the VectorBase Consortium; Madey, G.; et al. VectorBase: An updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef] [PubMed]
- Bridi, L.C. Mapeamento Físico de Genes Expressos de Resposta Imune e Sensorial de Anopheles (Nyssorhynchus) Darlingi, Vetor da Malária Neotropical. Ph.D. Thesis, National Institute of Amazon Research/INPA, Manaus, Brazil, 2016. [Google Scholar]
- Charlwood, J.D. Biological variation in Anopheles darlingi Root. Mem. Inst. Oswaldo Cruz 1996, 91, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Vittor, A.Y.; Gilman, R.H.; Tielsch, J.; Glass, G.; Shields, T.; Lozano, W.S.; Pinedo-Cancino, V.; Patz, J.A. The effect of deforestation on the human-biting rate of Anopheles darlingi, the primary vector of falciparum malaria in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2006, 74, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiwat, H.; Bretas, G. Ecology of Anopheles darlingi Root with respect to vector importance: A review. Parasit. Vectors 2011, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrelli, M.T.; Honório, N.A.; Flores-Mendoza, C.; Lourenço-de-Oliveira, R.; Marinotti, O.; Kloetzel, J.K. Comparative susceptibility of two members of the Anopheles oswaldoi complex, An. oswaldoi and An. konderi, to infection by Plasmodium vivax. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 381–384. [Google Scholar] [CrossRef]
- Okuda, K.; Caroci, A.; Ribolla, P.; Marinotti, O.; de Bianchi, A.G.; Bijovsky, A.T. Morphological and enzymatic analysis of the midgut of Anopheles darlingi during blood digestion. J. Insect Physiol. 2005, 51, 769–776. [Google Scholar] [CrossRef]
- Scarpassa, V.M.; Conn, J.E. Population genetic structure of the major malaria vector Anopheles darlingi (Diptera: Culicidae) from the Brazilian Amazon, using microsatellite markers. Mem. Inst. Oswaldo Cruz 2007, 102, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Vineis, J.H.; Yanoviak, S.P.; Scarpassa, V.M.; Póvoa, M.M.; Padilla, N.; Achee, N.L.; Conn, J.E. Microsatellite data suggest significant population structure and differentiation within the malaria vector Anopheles darlingi in Central and South America. BMC Ecol. 2008, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Perea, E.Z.; León, R.B.; Salcedo, M.P.; Brogdon, W.G.; Devine, G.J. Adaptation and evaluation of the bottle assay for monitoring insecticide resistance in disease vector mosquitoes in the Peruvian Amazon. Malar. J. 2009, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kobylinski, K.C.; Escobedo-Vargas, K.S.; López-Sifuentes, V.M.; Durand, S.; Smith, E.S.; Baldeviano, G.C.; Gerbasi, R.V.; Ballard, S.B.; Stoops, C.A.; Vásquez, G.M. Ivermectin susceptibility, sporontocidal effect, and inhibition of time to re-feed in the Amazonian malaria vector Anopheles darlingi. Malar. J. 2017, 16, 474. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, A.F.; Machado, L.C.; de Paula, M.B.; Vieira, C.J.D.S.P.; de Morais Bronzoni, R.V.; de Melo Santos, M.A.V.; Wallau, G.L. Culicidae evolutionary history focusing on the Culicinae subfamily based on mitochondrial phylogenomics. Sci. Rep. 2020, 10, 18823. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.M.; Wides, R.; et al. The genome sequence of the malaria mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Marinotti, O.; Krzywinski, J.; Tadei, W.P.; James, A.A.; Achee, L.N.; Conn, E.J. Complete mtDNA genomes of Anopheles darlingi and an approach to anopheline divergence time. Malaria J. 2010, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Guedes, A.S.; Amorim, E.M.; Schreiber, G. Análise dos cromossomos salivares em anofelinos brasileiros. Ver. Bras. Malar. D. Trop. 1957, 9, 247–250. [Google Scholar]
- Schreiber, G.; Guedes, A.S. Estudo comparativo do cromosoma X em algumas espécies de Anopheles do sub-gen. Nyssorhynchus (Dipt. Culic.). Ciên. Cult. 1959, 11, 128–129. [Google Scholar]
- Schreiber, G.; Guedes, A.S. Perspectivas citológicas na sistemática dos anofelinos (S.G Nyssorhynchus). Rev. Bras. Mal. D. Trop. 1960, 12, 355–358. [Google Scholar]
- Kreutzer, R.D.; Kitzmiller, J.B.; Ferreira, E. Inversion polymorphism in the salivary gland chromosomes of Anopheles darlingi Root. Mosq. News 1972, 32, 555–565. [Google Scholar]
- Tadei, W.P.; Santos, J.M.; Rabbani, M.G. Biologia de anofelinos amazônicos. V. Polimorfismo cromossômico de Anopheles darlingi Root (Diptera, Culicidae). Acta Amaz. 1982, 12, 353–369. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Armistead, J.S.; Provost-Javier, K.N.; Sakamoto, J.M.; Rasgon, J.L. Duplication, concerted evolution and purifying selection drive the evolution of mosquito vitellogenin genes. BMC Evol. Biol. 2010, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Coluzzi, M.; Sabatini, A.; Petrarca, V.; Dideco, M.A. Chromosomal differentiation and adaptation to human environments in the Anopheles gambiae complex. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 483–497. [Google Scholar] [CrossRef]
- Dobzhansky, T. Chromosomal Races in Drosophila Pseudoobscura and Drosophila Persimilis; Carnegie Institution of Washington: Washington, DC, USA, 1944; Volume 554, pp. 47–144. [Google Scholar]
- Hoffmann, A.A.; Sgrò, C.M.; Weeks, A.R. Chromosomal inversion polymorphisms and adaptation. Trends Ecol. Evol. 2004, 19, 482–488. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe. | BLAST Hits | In Situ Chromosome Mapping Anopheles darlingi | In Silico chromosome Mapping Anopheles albimanus | In Silico Chromosome Mapping Anopheles gambiae |
|---|---|---|---|---|
| Pb2r | scaffold_732 | 2R (8A, 10A, 10E) | 2R (10A) | 2R (18B) |
| scaffold_1942 | ||||
| scaffold_1482 | ||||
| Pb5r | scaffold_112 | 2R (15 B), 3R (31A) | 2R (12C) | 2R (12C) |
| Pb7b | scaffold_281 | 2R (6A) | 2R (10B) | 2R (13B) |
| Pb17r | scaffold_1409 | 2L (25C), 2R(9A) | 2L (24A) | 3R (32D) |
| scaffold_1350 | ||||
| Pb18b | scaffold_683 | 2L (16B) | 2L (17A) | 3R (29A) |
| scaffold_1062 | ||||
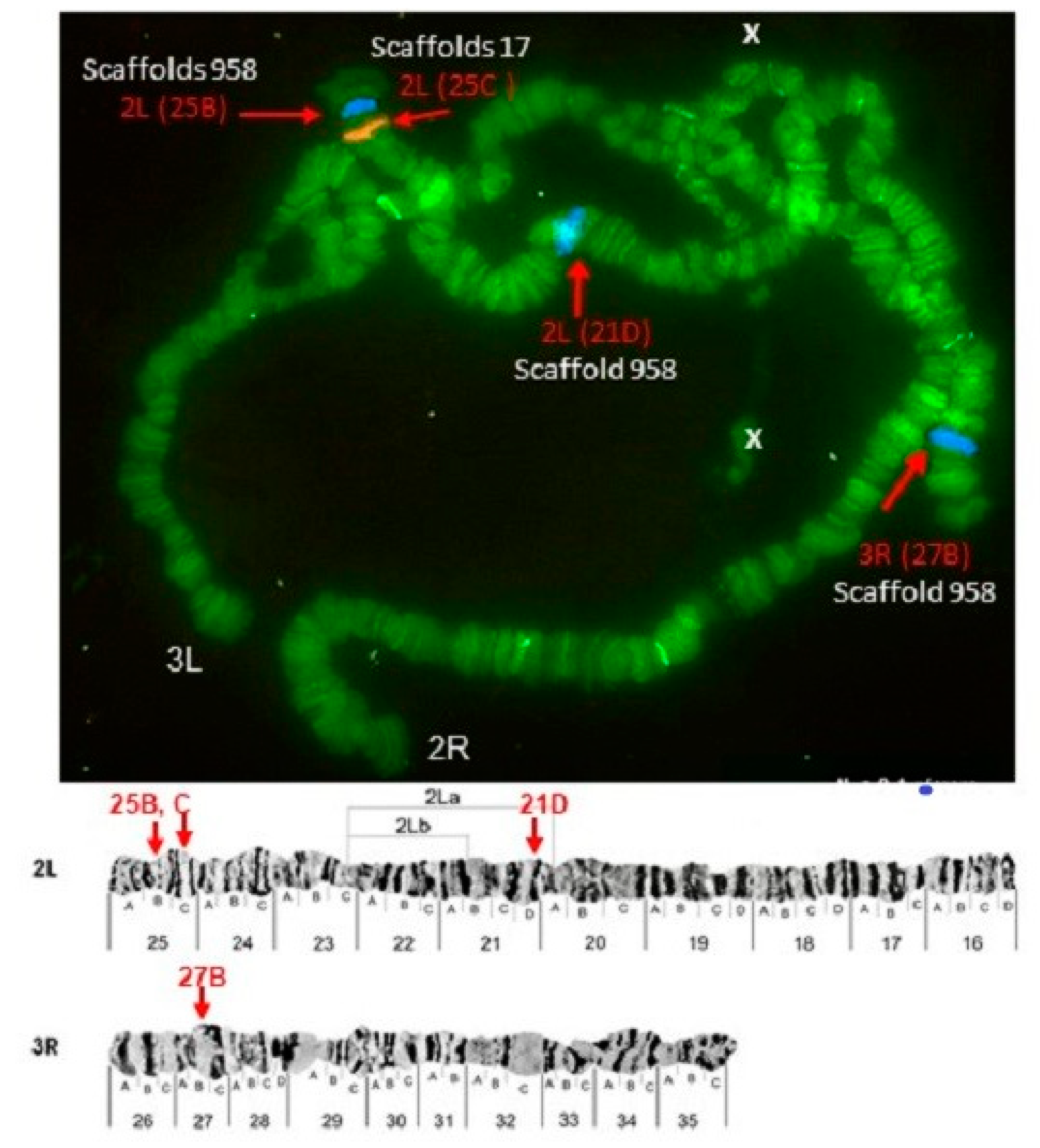
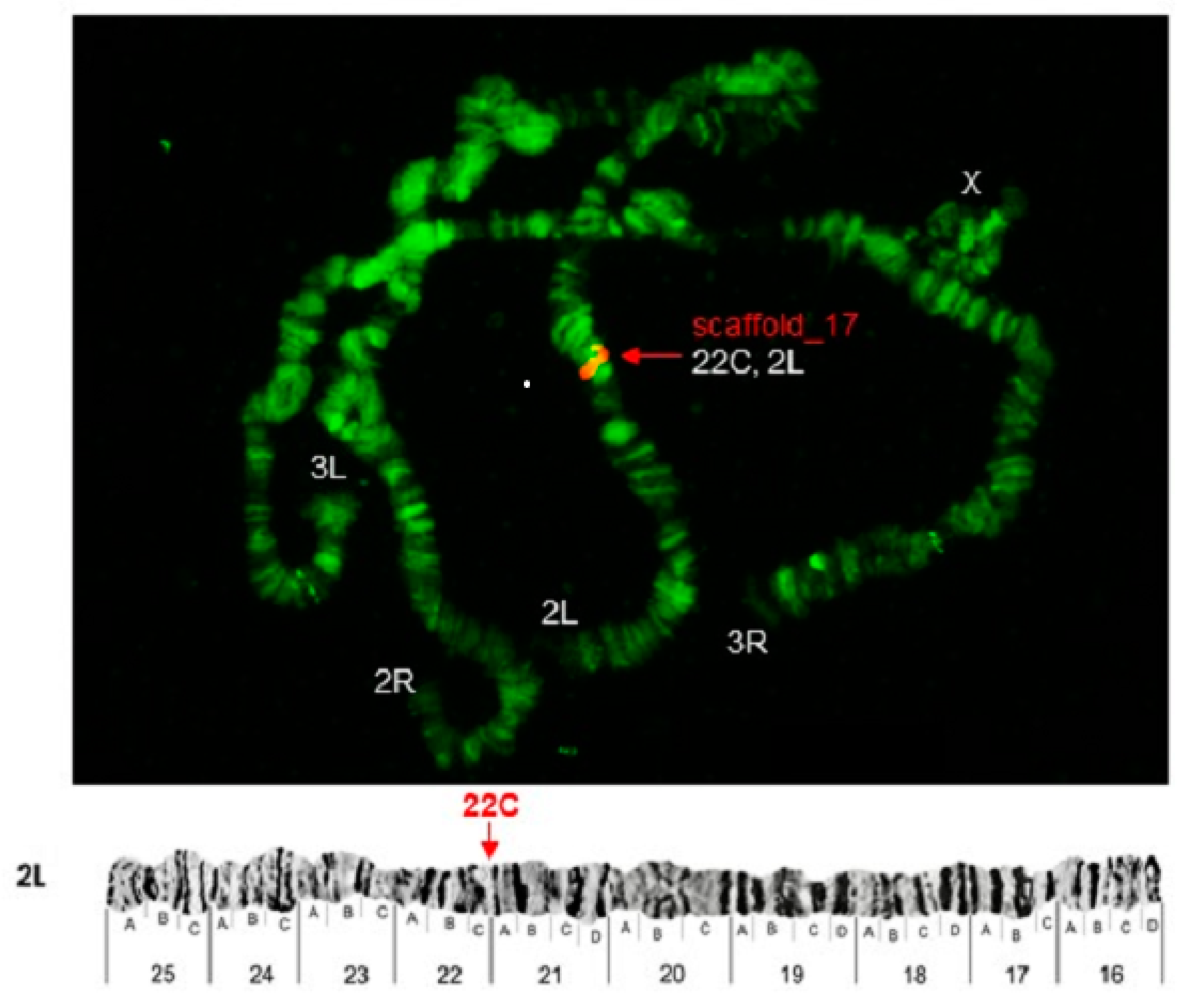
| Pb19r | scaffold_17 | 2L (25C) | 3L (45A) | 2L (21D) |
| Pb20b | scaffold_958 | 2L (21D,25B), 3R (27B) | X (1A) | X (5B) |
| Pb22b | scaffold_17 | 3L (43 C) | 3L (45A) | 2L (28C) |
| Pb23r | scaffold_17 | 2L (22C) | 3L (45A) | 2L (23D) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafael, M.S.; Bridi, L.C.; Sharakhov, I.V.; Marinotti, O.; Sharakhova, M.V.; Timoshevskiy, V.; Guimarães-Marques, G.M.; Santos, V.S.; da Silva, C.G.N.; Astolfi-Filho, S.; et al. Physical Mapping of the Anopheles (Nyssorhynchus) darlingi Genomic Scaffolds. Insects 2021, 12, 164. https://doi.org/10.3390/insects12020164
Rafael MS, Bridi LC, Sharakhov IV, Marinotti O, Sharakhova MV, Timoshevskiy V, Guimarães-Marques GM, Santos VS, da Silva CGN, Astolfi-Filho S, et al. Physical Mapping of the Anopheles (Nyssorhynchus) darlingi Genomic Scaffolds. Insects. 2021; 12(2):164. https://doi.org/10.3390/insects12020164
Chicago/Turabian StyleRafael, Míriam Silva, Leticia Cegatti Bridi, Igor V. Sharakhov, Osvaldo Marinotti, Maria V. Sharakhova, Vladimir Timoshevskiy, Giselle Moura Guimarães-Marques, Valéria Silva Santos, Carlos Gustavo Nunes da Silva, Spartaco Astolfi-Filho, and et al. 2021. "Physical Mapping of the Anopheles (Nyssorhynchus) darlingi Genomic Scaffolds" Insects 12, no. 2: 164. https://doi.org/10.3390/insects12020164