Evidence for Divergent Selection on Immune Genes between the African Malaria Vectors, Anopheles coluzzii and A. gambiae

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Species Identification
2.3. Immune-Related Genes in A. gambiae s.l.
2.4. SNP Discovery
2.5. SNP Genotyping
2.6. Genotype Data Analysis
2.7. Whole Genome Sequencing and Data Analysis
2.8. Data Accessibility
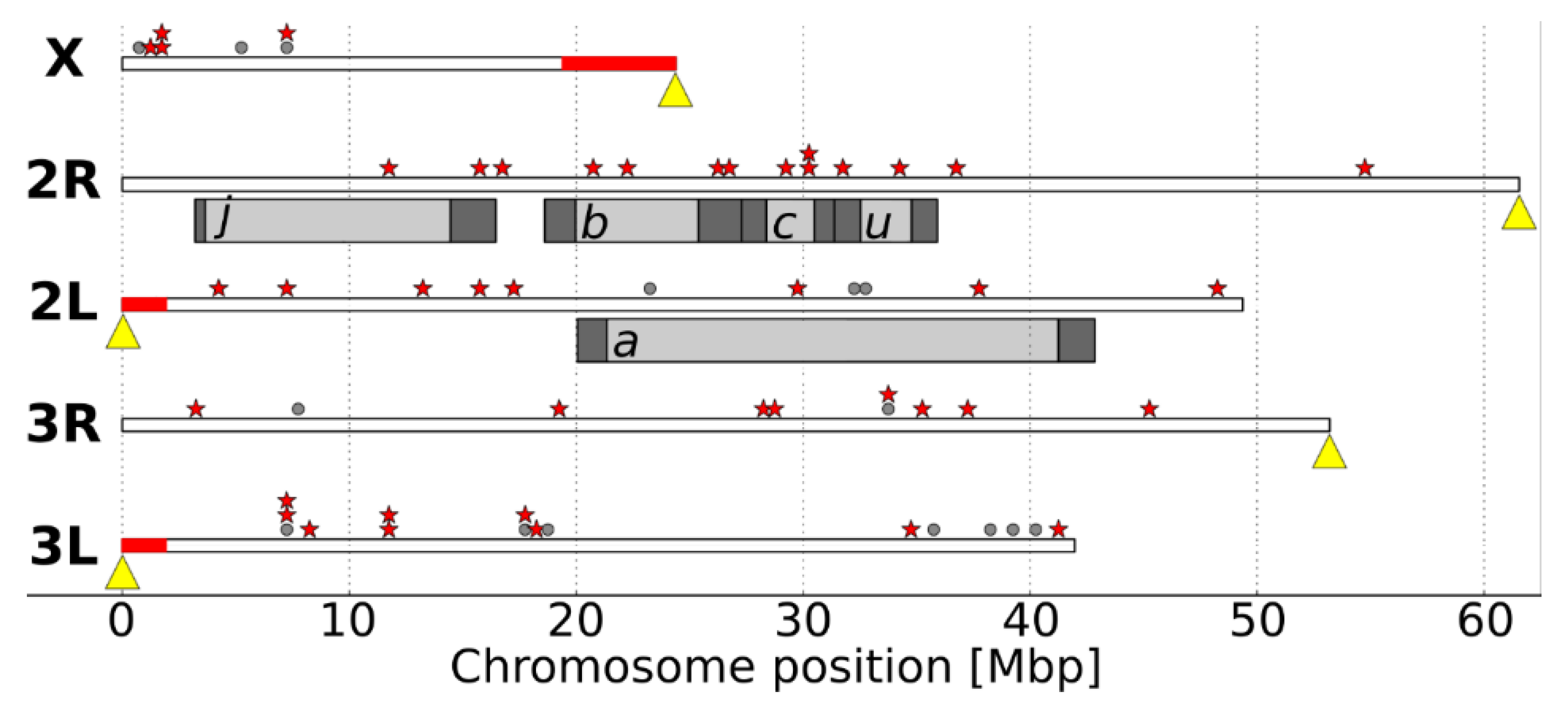
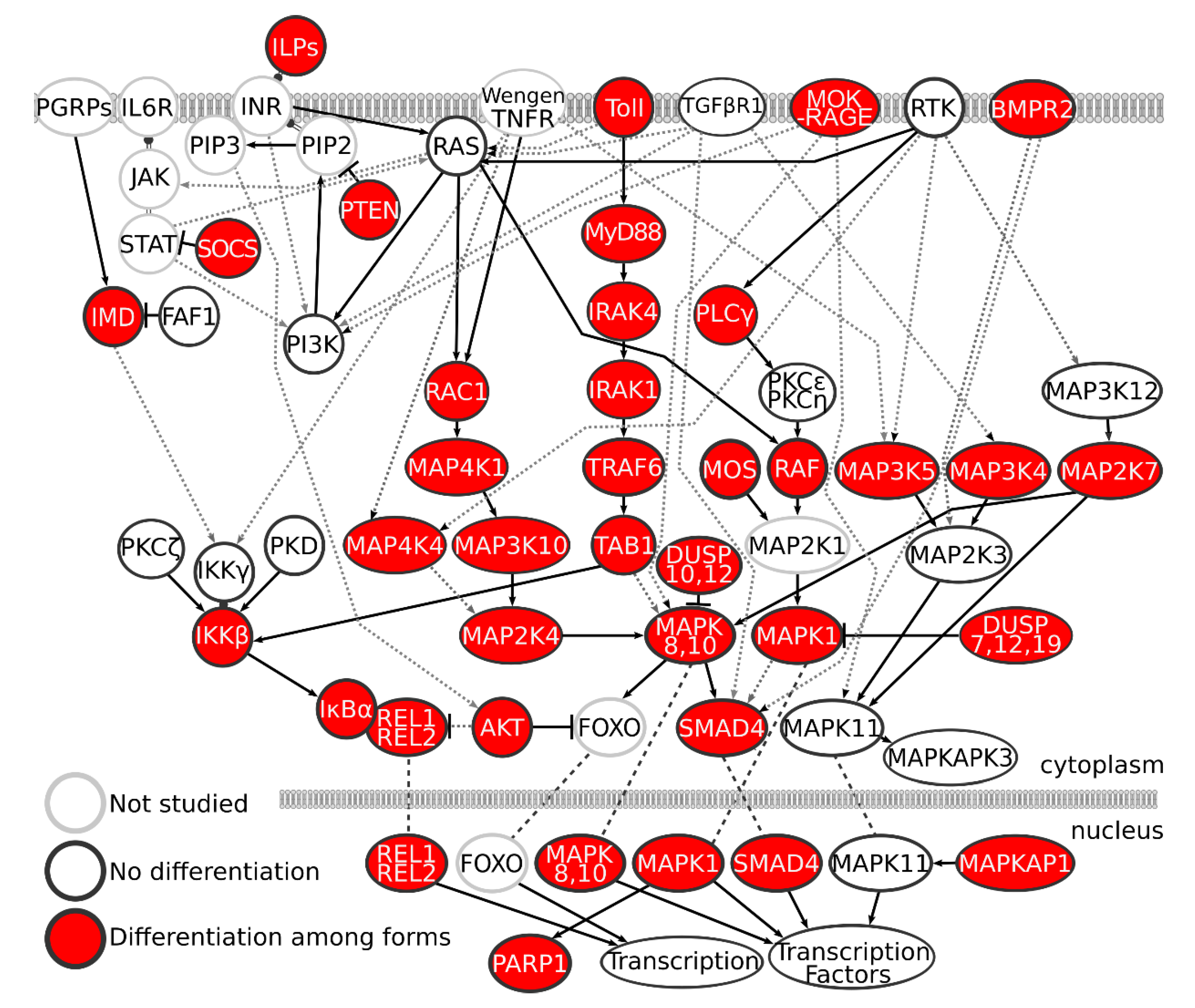
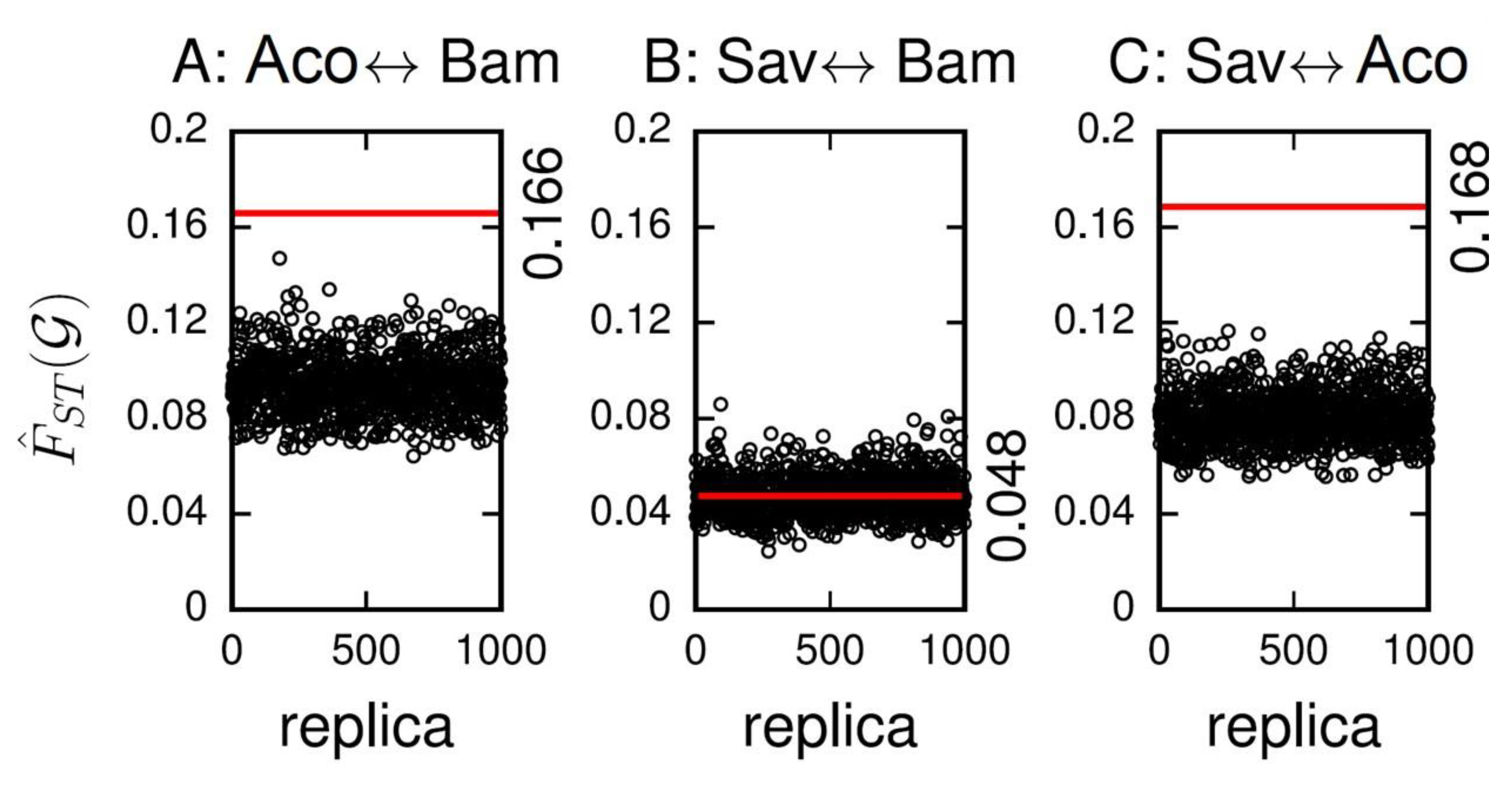
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wegner, K.M.; Kalbe, M.; Milinski, M.; Reusch, T.B. Mortality selection during the 2003 European heat wave in three-spined sticklebacks: Effects of parasites and MHC genotype. BMC Evol. Biol. 2008, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzaro, B.P.; Little, T.J. Immunity in a variable world. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2009, 364, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, R.; Bustamante, C.; Clark, A.G.; Glanowski, S.; Sackton, T.B.; Hubisz, M.J.; Fledel-Alon, A.; Tanenbaum, D.M.; Civello, D.; White, T.J.; et al. A scan for positively selected genes in the genomes of humans and chimpanzees. PLoS Biol. 2005, 3, e170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlenke, T.A.; Begun, D.J. Natural selection drives Drosophila immune system evolution. Genetics 2003, 164, 1471–1480. [Google Scholar] [PubMed]
- Obbard, D.J.; Welch, J.J.; Kim, K.W.; Jiggins, F.M. Quantifying adaptive evolution in the Drosophila immune system. PLoS Genet. 2009, 5, e1000698. [Google Scholar] [CrossRef] [Green Version]
- Crawford, J.E.; Bischoff, E.; Garnier, T.; Gneme, A.; Eiglmeier, K.; Holm, I.; Riehle, M.M.; Guelbeogo, W.M.; Sagnon, N.; Lazzaro, B.P.; et al. Evidence for population-specific positive selection on immune genes of Anopheles gambiae. G3 2012, 2, 1505–1519. [Google Scholar] [CrossRef]
- Sackton, T.B.; Lazzaro, B.P.; Schlenke, T.A.; Evans, J.D.; Hultmark, D.; Clark, A.G. Dynamic evolution of the innate immune system in Drosophila. Nat. Genet. 2007, 39, 1461–1468. [Google Scholar] [CrossRef]
- Riehle, M.M.; Markianos, K.; Niare, O.; Xu, J.; Li, J.; Toure, A.M.; Podiougou, B.; Oduol, F.; Diawara, S.; Diallo, M.; et al. Natural malaria infection in Anopheles gambiae is regulated by a single genomic control region. Science 2006, 312, 577–579. [Google Scholar] [CrossRef] [Green Version]
- Fryxell, R.T.; Nieman, C.C.; Fofana, A.; Lee, Y.; Traore, S.F.; Cornel, A.J.; Luckhart, S.; Lanzaro, G.C. Differential Plasmodium falciparum infection of Anopheles gambiae s.s. molecular and chromosomal forms in Mali. Malar. J. 2012, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Ndiath, M.O.; Cohuet, A.; Gaye, A.; Konate, L.; Mazenot, C.; Faye, O.; Boudin, C.; Sokhna, C.; Trape, J.F. Comparative susceptibility to Plasmodium falciparum of the molecular forms M and S of Anopheles gambiae and Anopheles arabiensis. Malar. J. 2011, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Beier, J.C.; Killeen, G.F.; Githure, J.I. Short report: Entomologic inoculation rates and Plasmodium falciparum malaria prevalence in Africa. Am. J. Trop. Med. Hyg. 1999, 61, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dia, I.; Ba, H.; Mohamed, S.A.; Diallo, D.; Lo, B.; Diallo, M. Distribution, host preference and infection rates of malaria vectors in Mauritania. Parasites Vectors 2009, 2, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayton, A.M.; Dong, Y.; Dimopoulos, G. The Anopheles innate immune system in the defense against malaria infection. J. Innate Immun. 2014, 6, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Surachetpong, W.; Pakpour, N.; Cheung, K.W.; Luckhart, S. Reactive oxygen species-dependent cell signaling regulates the mosquito immune response to Plasmodium falciparum. Antioxid. Redox Signal. 2011, 14, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Corby-Harris, V.; Drexler, A.; Watkins de Jong, L.; Antonova, Y.; Pakpour, N.; Ziegler, R.; Ramberg, F.; Lewis, E.E.; Brown, J.M.; Luckhart, S.; et al. Activation of Akt signaling reduces the prevalence and intensity of malaria parasite infection and lifespan in Anopheles stephensi mosquitoes. PLoS Pathog 2010, 6, e1001003. [Google Scholar] [CrossRef]
- Souvannaseng, L.; Hun, L.V.; Baker, H.; Klyver, J.M.; Wang, B.; Pakpour, N.; Bridgewater, J.M.; Napoli, E.; Giulivi, C.; Riehle, M.A.; et al. Inhibition of JNK signaling in the Asian malaria vector Anopheles stephensi extends mosquito longevity and improves resistance to Plasmodium falciparum infection. PLoS Pathog 2018, 14, e1007418. [Google Scholar] [CrossRef] [Green Version]
- Weiss, B.; Aksoy, S. Microbiome influences on insect host vector competence. Trends Parasitol. 2011, 27, 514–522. [Google Scholar] [CrossRef] [Green Version]
- Telang, A.; Qayum, A.A.; Parker, A.; Sacchetta, B.R.; Byrnes, G.R. Larval nutritional stress affects vector immune traits in adult yellow fever mosquito Aedes aegypti (Stegomyia aegypti). Med. Vet. Entomol. 2012, 26, 271–281. [Google Scholar] [CrossRef]
- Muturi, E.J.; Blackshear, M., Jr.; Montgomery, A. Temperature and density-dependent effects of larval environment on Aedes aegypti competence for an alphavirus. J. Vector Ecol. 2012, 37, 154–161. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Estevez-Lao, T.Y. Nitric oxide is an essential component of the hemocyte-mediated mosquito immune response against bacteria. Dev. Comp. Immunol. 2010, 34, 141–149. [Google Scholar] [CrossRef]
- Okech, B.A.; Gouagna, L.C.; Yan, G.; Githure, J.I.; Beier, J.C. Larval habitats of Anopheles gambiae s.s. (Diptera: Culicidae) influences vector competence to Plasmodium falciparum parasites. Malar. J. 2007, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Rottschaefer, S.M.; Crawford, J.E.; Riehle, M.M.; Guelbeogo, W.M.; Gneme, A.; Sagnon, N.; Vernick, K.D.; Lazzaro, B.P. Population Genetics of Anopheles coluzzii Immune Pathways and Genes. G3 Genes Genom. Genet. 2015, 5, 329–339. [Google Scholar] [CrossRef] [Green Version]
- della Torre, A.; Costantini, C.; Besansky, N.J.; Caccone, A.; Petrarca, V.; Powell, J.R.; Coluzzi, M. Speciation within Anopheles gambiae—The glass is half full. Science 2002, 298, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.L.; Hahn, M.W.; Nuzhdin, S.V. Genomic islands of speciation in Anopheles gambiae. PLoS Biol. 2005, 3, e285. [Google Scholar] [CrossRef] [PubMed]
- Lanzaro, G.C.; Lee, Y. Speciation in Anopheles gambiae—The Distribution of Genetic Polymorphism and Patterns of Reproductive Isolation Among Natural Populations. In Anopheles Mosquitoes—New Insights into Malaria Vector; Manguin, S., Ed.; InTech: Rijeka, Croatia, 2013. [Google Scholar]
- Coetzee, M.; Hunt, R.H.; Wilkerson, R.; Della Torre, A.; Coulibaly, M.B.; Besansky, N.J. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa 2013, 3619, 246–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touré, Y.T.; Petrarca, V.; Traoré, S.F.; Coulibaly, A.; Maiga, H.M.; Sankare, O.; Sow, M.; Di Deco, M.A.; Coluzzi, M. The distribution and inversion polymorphism of chromosomally recognized taxa of the Anopheles gambiae complex in Mali, West Africa. Parassitologia 1998, 40, 477–511. [Google Scholar]
- Lee, Y.; Collier, T.C.; Sanford, M.R.; Marsden, C.D.; Fofana, A.; Cornel, A.J.; Lanzaro, G.C. Chromosome inversions, genomic differentiation and speciation in the African malaria mosquito Anopheles gambiae. PLoS ONE 2013, 8, e57887. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.; Touré, Y.T.; Carnahan, J.; Norris, D.E.; Dolo, G.; Traoré, S.F.; Edillo, F.E.; Lanzaro, G.C. Gene flow among populations of the malaria vector, Anopheles gambiae, in Mali, West Africa. Genetics 2001, 157, 743–750. [Google Scholar]
- Favia, G.; della Torre, A.; Bagayoko, M.; Lanfrancotti, A.; Sagnon, N.; Toure, Y.T.; Coluzzi, M. Molecular identification of sympatric chromosomal forms of Anopheles gambiae and further evidence of their reproductive isolation. Insect Mol. Biol. 1997, 6, 377–383. [Google Scholar] [CrossRef]
- Fanello, C.; Santolamazza, F.; della Torre, A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP. Med. Vet. Entomol. 2002, 16, 461–464. [Google Scholar] [CrossRef]
- White, B.J.; Cheng, C.; Simard, F.; Costantini, C.; Besansky, N.J. Genetic association of physically unlinked islands of genomic divergence in incipient species of Anopheles gambiae. Mol. Ecol. 2010, 19, 925–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimonneau, G.; Pombi, M.; Choisy, M.; Morand, S.; Dabire, R.K.; Simard, F. Larval habitat segregation between the molecular forms of the mosquito Anopheles gambiae in a rice field area of Burkina Faso, West Africa. Med. Vet. Entomol. 2012, 26, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Sanford, M.R.; Ramsay, S.; Cornel, A.J.; Marsden, C.D.; Norris, L.C.; Patchoke, S.; Fondjo, E.; Lanzaro, G.C.; Lee, Y. A preliminary investigation of the relationship between water quality and Anopheles gambiae larval habitats in Western Cameroon. Malar. J. 2013, 12, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoukis, N.C.; Powell, J.R.; Toure, M.B.; Sacko, A.; Edillo, F.E.; Coulibaly, M.B.; Traore, S.F.; Taylor, C.E.; Besansky, N.J. A test of the chromosomal theory of ecotypic speciation in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2008, 105, 2940–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riehle, M.M.; Guelbeogo, W.M.; Gneme, A.; Eiglmeier, K.; Holm, I.; Bischoff, E.; Garnier, T.; Snyder, G.M.; Li, X.; Markianos, K.; et al. A cryptic subgroup of Anopheles gambiae is highly susceptible to human malaria parasites. Science 2011, 331, 596–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coluzzi, M.; Petrarca, V.; Di Deco, M. Chromosomal inversion intergradation and incipient speciation in Anopheles gambiae. Boll. Zool. 1985, 52, 45–63. [Google Scholar] [CrossRef]
- Kimchi-Sarfaty, C.; Oh, J.M.; Kim, I.W.; Sauna, Z.E.; Calcagno, A.M.; Ambudkar, S.V.; Gottesman, M.M. A “silent“ polymorphism in the MDR1 gene changes substrate specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Purvis, I.J.; Bettany, A.J.; Santiago, T.C.; Coggins, J.R.; Duncan, K.; Eason, R.; Brown, A.J. The efficiency of folding of some proteins is increased by controlled rates of translation in vivo. A hypothesis. J. Mol. Biol. 1987, 193, 413–417. [Google Scholar] [CrossRef]
- Brenton, A.A.; Souvannaseng, L.; Cheung, K.; Anishchenko, M.; Brault, A.C.; Luckhart, S. Engineered single nucleotide polymorphisms in the mosquito MEK docking site alter Plasmodium berghei development in Anopheles gambiae. Parasites Vectors 2014, 7, 287. [Google Scholar] [CrossRef] [Green Version]
- Hunt, R.H. A cytological technique for the study of Anopheles gambiae complex. Parassitologia 1973, 15, 137–139. [Google Scholar]
- Scott, J.A.; Brogdon, W.G.; Collins, F.H. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am. J. Trop. Med. Hyg. 1993, 49, 520–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favia, G.; Lanfrancotti, A.; Spanos, L.; Siden-Kiamos, I.; Louis, C. Molecular characterization of ribosomal DNA polymorphisms discriminating among chromosomal forms of Anopheles gambiae s.s. Insect Mol. Biol. 2001, 10, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Coluzzi, M.; Sabatini, A.; della Torre, A.; Di Deco, M.A.; Petrarca, V. A polytene chromosome analysis of the Anopheles gambiae species complex. Science 2002, 298. [Google Scholar] [CrossRef] [PubMed]
- Christophides, G.K.; Zdobnov, E.; Barillas-Mury, C.; Birney, E.; Blandin, S.; Blass, C.; Brey, P.T.; Collins, F.H.; Danielli, A.; Dimopoulos, G.; et al. Immunity-related genes and gene families in Anopheles gambiae. Science 2002, 298, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Garver, L.S.; Bahia, A.C.; Das, S.; Souza-Neto, J.A.; Shiao, J.; Dong, Y.; Dimopoulos, G. Anopheles Imd pathway factors and effectors in infection intensity-dependent anti-Plasmodium action. PLoS Pathog 2012, 8, e1002737. [Google Scholar] [CrossRef]
- Barillas-Mury, C.; Han, Y.S.; Seeley, D.; Kafatos, F.C. Anopheles gambiae Ag-STAT, a new insect member of the STAT family, is activated in response to bacterial infection. EMBO J. 1999, 18, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Pakpour, N.; Napoli, E.; Drexler, A.; Glennon, E.K.; Surachetpong, W.; Cheung, K.; Aguirre, A.; Klyver, J.M.; Lewis, E.E.; et al. Anopheles stephensi p38 MAPK signaling regulates innate immunity and bioenergetics during Plasmodium falciparum infection. Parasites Vectors 2015, 8, 424. [Google Scholar] [CrossRef] [Green Version]
- Surachetpong, W.; Singh, N.; Cheung, K.W.; Luckhart, S. MAPK ERK signaling regulates the TGF-beta1-dependent mosquito response to Plasmodium falciparum. PLoS Pathog 2009, 5, e1000366. [Google Scholar] [CrossRef] [Green Version]
- Pietri, J.E.; Cheung, K.W.; Luckhart, S. Knockdown of mitogen-activated protein kinase (MAPK) signalling in the midgut of Anopheles stephensi mosquitoes using antisense morpholinos. Insect Mol. Biol. 2014, 23, 558–565. [Google Scholar] [CrossRef] [Green Version]
- Ferrandon, D.; Imler, J.L.; Hetru, C.; Hoffmann, J.A. The Drosophila systemic immune response: Sensing and signalling during bacterial and fungal infections. Nat. Rev. Immunol. 2007, 7, 862–874. [Google Scholar] [CrossRef]
- Napetschnig, J.; Wu, H. Molecular basis of NF-kappaB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Gowda, D.C.; Krishnegowda, G.; Luckhart, S. Induction of nitric oxide synthase in Anopheles stephensi by Plasmodium falciparum: Mechanism of signaling and the role of parasite glycosylphosphatidylinositols. Infect Immun. 2005, 73, 2778–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luckhart, S.; Giulivi, C.; Drexler, A.L.; Antonova-Koch, Y.; Sakaguchi, D.; Napoli, E.; Wong, S.; Price, M.S.; Eigenheer, R.; Phinney, B.S.; et al. Sustained activation of Akt elicits mitochondrial dysfunction to block Plasmodium falciparum infection in the mosquito host. PLoS Pathog 2013, 9, e1003180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakpour, N.; Corby-Harris, V.; Green, G.P.; Smithers, H.M.; Cheung, K.W.; Riehle, M.A.; Luckhart, S. Ingested human insulin inhibits the mosquito NF-kappaB-dependent immune response to Plasmodium falciparum. Infect Immun. 2012, 80, 2141–2149. [Google Scholar] [CrossRef] [Green Version]
- Crampton, A.; Luckhart, S. The role of As60A, a TGF-beta homolog, in Anopheles stephensi innate immunity and defense against Plasmodium infection. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2001, 1, 131–141. [Google Scholar]
- Price, I.; Ermentrout, B.; Zamora, R.; Wang, B.; Azhar, N.; Mi, Q.; Constantine, G.; Faeder, J.R.; Luckhart, S.; Vodovotz, Y. In vivo, in vitro, and in silico studies suggest a conserved immune module that regulates malaria parasite transmission from mammals to mosquitoes. J. Theor. Biol. 2013, 334, 173–186. [Google Scholar] [CrossRef] [Green Version]
- Vodovotz, Y.; Zamora, R.; Lieber, M.J.; Luckhart, S. Cross-talk between nitric oxide and transforming growth factor-beta1 in malaria. Curr. Mol. Med. 2004, 4, 787–797. [Google Scholar] [CrossRef]
- Collins, F.H.; Sakai, R.K.; Vernick, K.D.; Paskewitz, S.; Seeley, D.C.; Miller, L.H.; Collins, W.E.; Campbell, C.C.; Gwadz, R.W. Genetic selection of a Plasmodium-refractory strain of the malaria vector Anopheles gambiae. Science 1986, 234, 607–610. [Google Scholar] [CrossRef]
- Rios-Velasquez, C.M.; Martins-Campos, K.M.; Simoes, R.C.; Izzo, T.; Dos Santos, E.V.; Pessoa, F.A.; Lima, J.B.; Monteiro, W.M.; Secundino, N.F.; Lacerda, M.V.; et al. Experimental Plasmodium vivax infection of key Anopheles species from the Brazilian Amazon. Malar. J. 2013, 12, 460. [Google Scholar] [CrossRef] [Green Version]
- Komar, A.A.; Lesnik, T.; Reiss, C. Synonymous codon substitutions affect ribosome traffic and protein folding during in vitro translation. FEBS Lett. 1999, 462, 387–391. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 6 March 2020).
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Hofer, T.; Foll, M. Detecting loci under selection in a hierarchically structured population. Heredity 2009, 103, 285–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Shete, S.; Swartz, M.; Amos, C.I. Examining the effect of linkage disequilibrium on multipoint linkage analysis. BMC Genet. 2005, 6 (Suppl. 1), S83. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Norris, L.C.; Main, B.J.; Lee, Y.; Collier, T.C.; Fofana, A.; Cornel, A.J.; Lanzaro, G.C. Adaptive introgression in an African malaria mosquito coincident with the increased usage of insecticide-treated bed nets. Proc. Natl. Acad. Sci. USA 2015, 112, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Giraldo-Calderon, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; VectorBase, C.; Madey, G.; et al. VectorBase: An updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997v1302. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Davison, A.C.; Hinkley, D.V. Bootstrap Methods and Their Application; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1997; 582p. [Google Scholar]
- North, B.V.; Curtis, D.; Sham, P.C. A note on the calculation of empirical P values from Monte Carlo procedures. Am. J. Hum. Genet. 2002, 71, 439–441. [Google Scholar] [CrossRef] [Green Version]
- Betran, E.; Thornton, K.; Long, M. Retroposed new genes out of the X in Drosophila. Genome Res. 2002, 12, 1854–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, J.J.; Kaessmann, H.; Betran, E.; Long, M. Extensive gene traffic on the mammalian X chromosome. Science 2004, 303, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, P. Variation in immune defence as a question of evolutionary ecology. Proc. Biol. Sci. R. Soc. 2003, 270, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawniczak, M.K.; Emrich, S.J.; Holloway, A.K.; Regier, A.P.; Olson, M.; White, B.; Redmond, S.; Fulton, L.; Appelbaum, E.; Godfrey, J.; et al. Widespread divergence between incipient Anopheles gambiae species revealed by whole genome sequences. Science 2010, 330, 512–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, K.J.; Dudley, J.T.; Kukurba, K.R.; Chen, R.; Butte, A.J.; Montgomery, S.B.; Snyder, M. Systematic functional regulatory assessment of disease-associated variants. Proc. Natl. Acad. Sci. USA 2013, 110, 9607–9612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diabate, A.; Dabire, R.K.; Kim, E.H.; Dalton, R.; Millogo, N.; Baldet, T.; Simard, F.; Gimnig, J.E.; Hawley, W.A.; Lehmann, T. Larval development of the molecular forms of Anopheles gambiae (Diptera: Culicidae) in different habitats: A transplantation experiment. J. Med. Entomol. 2005, 42, 548–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- League, G.P.; Estévez-Lao, T.Y.; Yan, Y.; Garcia-Lopez, V.A.; Hillyer, J.F. Anopheles gambiae larvae mount stronger immune responses against bacterial infection than adults: Evidence of adaptive decoupling in mosquitoes. Parasites Vectors 2017, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Coluzzi, M.; Sabatini, A.; Petrarca, V.; Di Deco, M.A. Chromosomal differentiation and adaptation to human environments in the Anopheles gambiae complex. Trans. R Soc. Trop. Med. Hyg. 1979, 73, 483–497. [Google Scholar] [CrossRef]
- Persiani, A.; Di Deco, M.A.; Petrangeli, G. Laboratory observation of inversion polymorphisms originating from the crossing of various populations of Anopheles gambiae s.s. Ann. Ist. Super Sanita 1986, 22, 221–223. [Google Scholar]
- Diabate, A.; Dabire, R.K.; Millogo, N.; Lehmann, T. Evaluating the effect of postmating isolation between molecular forms of Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 60–64. [Google Scholar] [CrossRef]
- Pennetier, C.; Warren, B.; Dabire, K.R.; Russell, I.J.; Gibson, G. “Singing on the wing” as a mechanism for species recognition in the malarial mosquito Anopheles gambiae. Curr. Biol. 2010, 20, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzaro, B.P.; Clark, A.G. Molecular population genetics of inducible antibacterial peptide genes in Drosophila melanogaster. Mol. Biol. Evol. 2003, 20, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Hill-Burns, E.M.; Clark, A.G. Functional regulatory divergence of the innate immune system in interspecific Drosophila hybrids. Mol. Biol. Evol. 2010, 27, 2596–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Marsden, C.D.; Norris, L.C.; Collier, T.C.; Main, B.J.; Fofana, A.; Cornel, A.J.; Lanzaro, G.C. Spatiotemporal dynamics of gene flow and hybrid fitness between the M and S forms of the malaria mosquito, Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2013, 110, 19854–19859. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Chromosome | Diverged Genes | Total Genes Genotyped | % Divergence |
|---|---|---|---|
| X | 4 | 7 | 57.1% |
| 2L | 8 | 11 | 72.7% |
| 2R | 14 | 14 | 100% |
| 3L | 9 | 16 | 56.3% |
| 3R | 8 | 10 | 80.0% |
| Total | 43 | 58 | 72.9% |
| Chromosome | SNP | Genotype | Ac | Bam | Sav |
|---|---|---|---|---|---|
| 2R | MAP2K4-164 | C/C | 164 | 13 | 58 |
| C/T | 1 | 74 | 27 | ||
| T/T | 0 | 87 | 1 | ||
| 2L | Raf-784 | A/A | 22 | 128 | 82 |
| A/G | 75 | 3 | 0 | ||
| G/G | 55 | 0 | 0 | ||
| 2L | Raf-851 | A/A | 12 | 2 | 69 |
| A/C | 78 | 20 | 2 | ||
| C/C | 60 | 106 | 3 |
| Forms | r2 > 0.5 | r2 > 0.8 | r2 > 0.95 |
|---|---|---|---|
| LD between genes | |||
| A. gambiae (Bamako) | 32.1% | 28.6% | 25.2% |
| A. coluzzii | 30.6% | 27.4% | 23.8% |
| A. gambiae (Savanna) | 32.7% | 29.7% | 26.0% |
| LD within a gene | |||
| A. gambiae (Bamako) | 35.4% | 30.9% | 27.1% |
| A. coluzzii | 34.3% | 28.4% | 25.9% |
| A. gambiae (Savanna) | 34.8% | 31.3% | 26.7% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Souvannaseng, L.; Collier, T.C.; Main, B.J.; Norris, L.C.; Fofana, A.; Traoré, S.F.; Cornel, A.J.; Luckhart, S.; Lanzaro, G.C. Evidence for Divergent Selection on Immune Genes between the African Malaria Vectors, Anopheles coluzzii and A. gambiae. Insects 2020, 11, 893. https://doi.org/10.3390/insects11120893
Lee Y, Souvannaseng L, Collier TC, Main BJ, Norris LC, Fofana A, Traoré SF, Cornel AJ, Luckhart S, Lanzaro GC. Evidence for Divergent Selection on Immune Genes between the African Malaria Vectors, Anopheles coluzzii and A. gambiae. Insects. 2020; 11(12):893. https://doi.org/10.3390/insects11120893
Chicago/Turabian StyleLee, Yoosook, Lattha Souvannaseng, Travis C. Collier, Bradley J. Main, Laura C. Norris, Abdarahamane Fofana, Sekou F. Traoré, Anthony J. Cornel, Shirley Luckhart, and Gregory C. Lanzaro. 2020. "Evidence for Divergent Selection on Immune Genes between the African Malaria Vectors, Anopheles coluzzii and A. gambiae" Insects 11, no. 12: 893. https://doi.org/10.3390/insects11120893