First Account of Phylogeographic Variation, Larval Characters, and Laboratory Rearing of the Endangered Cobblestone Tiger Beetle Cicindelidia marginipennis, Dejean, 1831 with Observations of Their Natural History

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection
2.2. DNA Extraction and Sequencing
2.3. Sequence Alignment and Network Reconstruction
2.4. Phylogenetic Analysis of COI
2.5. Laboratory Rearing of Cicindelidia marginipennis
2.6. Morphological Description of Larvae
3. Results
3.1. mtDNA Sequencing and Analyses
3.2. Phylogeographic Diversity
3.3. Laboratory Rearing
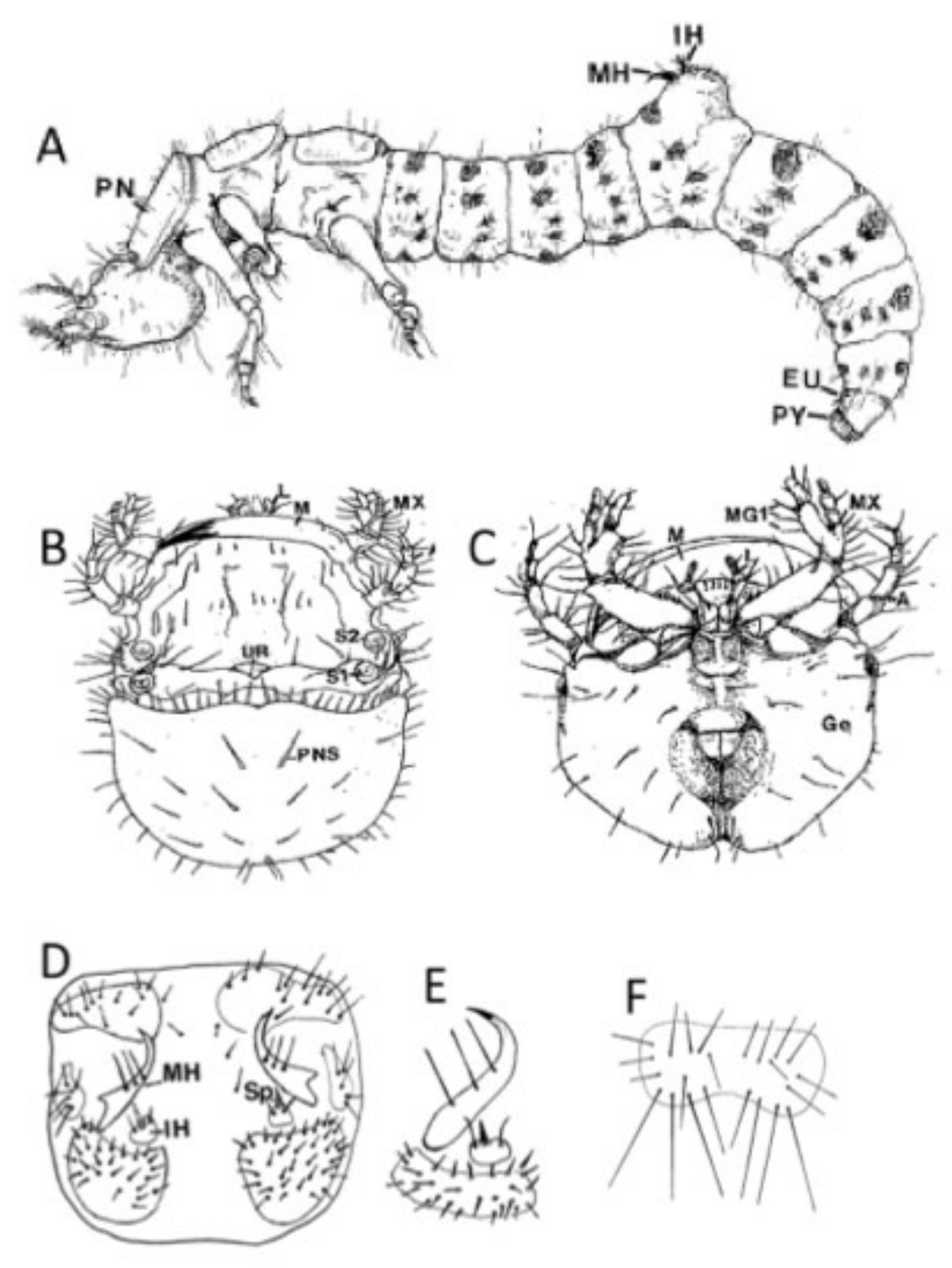
3.4. Larval Descriptions
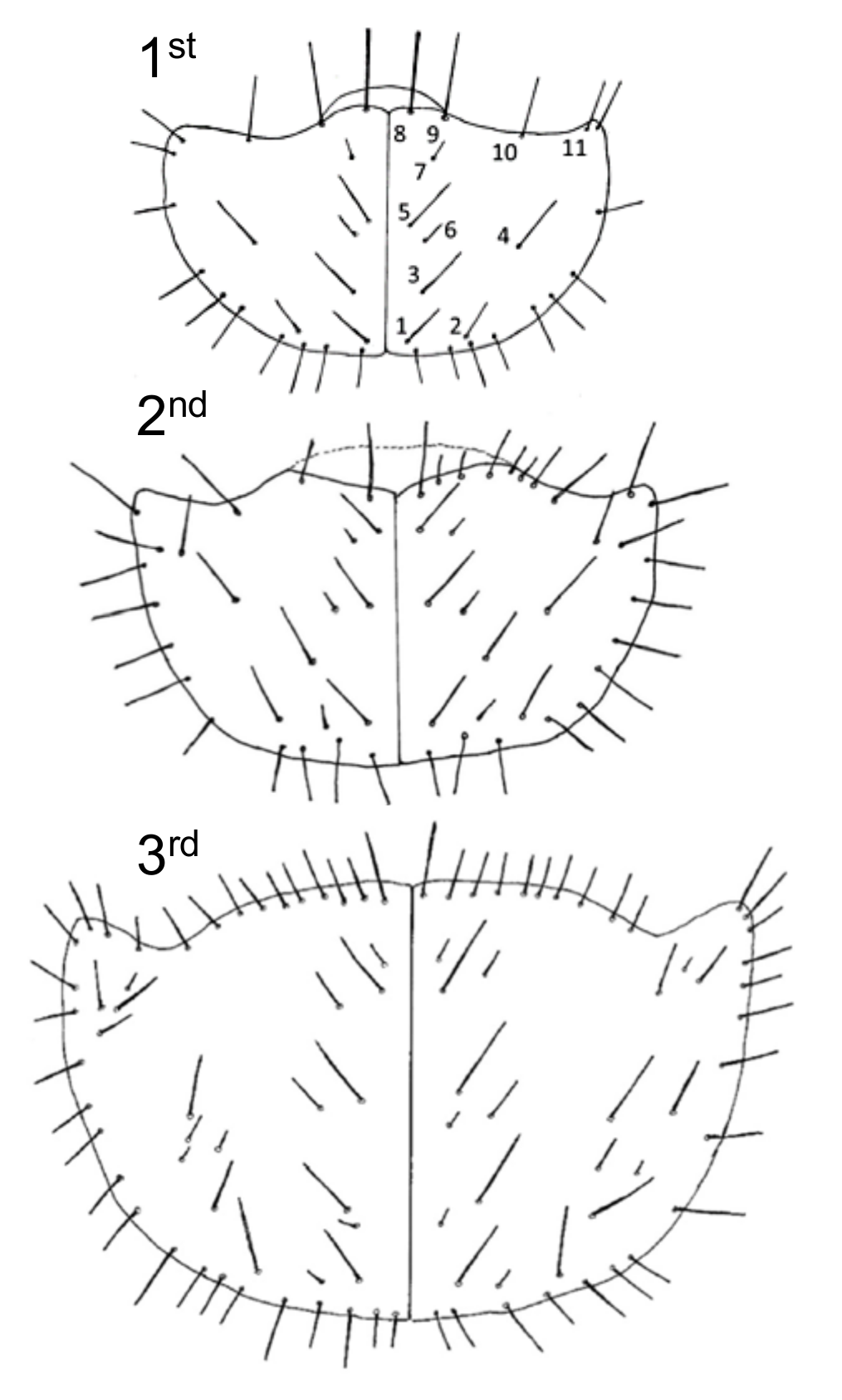
3.4.1. First (1st) Instar Larva
3.4.2. Second (2nd) Instar Larva
3.4.3. Third (3rd) Instar Larva
3.4.4. Key Characters for Distinguishing 3rd Instar Cicindelidia marginipennis
- Head and pronotum black;
- Twenty-six to 38 setae on the pronotal disc (but many small to minute);
- Inner hooks with spine about one-half or greater than total length of the spine;
- Four pairs of setae on 9th eusternum;
- Three setae on each medial hook.
4. Discussion
4.1. Cryptic Diversity within C. Marginipennis
4.2. Future Recommendations for Rearing C. marginipennis
4.3. Finding Larvae in the Field
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duran, D.P.; Gough, H.M. Validation of tiger beetles as distinct family (Coleoptera: Cicindelidae), review and reclassification of tribal relationships. Syst. Èntomol. 2020, 45, 723–729. [Google Scholar] [CrossRef]
- Mizutani, A.; Toh, Y. Behavioral analysis of two distinct visual responses in the larva of the tiger beetle (Cicindela chinensis). J. Comp. Physiol. A 1998, 182, 277–286. [Google Scholar] [CrossRef]
- Pearson, D.L.; Vogler, A.P. Tiger Beetles, 1st ed.; Cornell Series in Arthropod Biology; Cornell University Press: Ithaca, NY, USA, 2001; 352p. [Google Scholar]
- Toh, Y.; Okamura, J.Y. Behavioural responses of the tiger beetle larva to moving objects: Role of binocular and monocular vision. J. Exp. Biol. 2001, 204, 615–625. [Google Scholar] [PubMed]
- New, T.R. Beetles in Conservation, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; 248p. [Google Scholar]
- Pearson, D.L.; Knisley, C.B.; Duran, D.P.; Kazilek, C.J. A Field Guide to the Tiger Beetles of the United States and Canada: Identification, Natural History and Distribution of the Cicindelidae, 2nd ed.; Oxford University Press: New York, NY, USA, 2015; 251p. [Google Scholar]
- Pearson, D.L.; Shetterly, J.A. How Do Published Field Guides Influence Interactions between Amateurs and Professionals in Entomology? Am. Èntomol. 2006, 52, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Pearson, D.L.; Cassola, F. World-Wide Species Richness Patterns of Tiger Beetles (Coleoptera: Cicindelidae): Indicator Taxon for Biodiversity and Conservation Studies. Conserv. Biol. 1992, 6, 376–391. [Google Scholar] [CrossRef]
- Pearson, D.L.; Carroll, S.S. Global Patterns of Species Richness: Spatial Models for Conservation Planning Using Bioindicator and Precipitation Data. Conserv. Biol. 1998, 12, 809–821. [Google Scholar] [CrossRef]
- Knisley, C.B. Anthropogenic disturbances and rare tiger beetle habitats: Benefits, risks, and implications for conservation. Terr. Arthropod Rev. 2011, 4, 41–61. [Google Scholar] [CrossRef]
- Wuebbles, D.; Easterling, D.; Hayhoe, K.; Knutson, T.; Kopp, R.; Kossin, J.P.; Kunkel, K.; LeGrande, A.; Mears, C.; Sweet, W.; et al. Our Globally Changing Climate. In Climate Science Special Report: Fourth National Climate Assessment; Wuebbles, D.J., Fahey, D.W., Hibbard, K.A., Dokken, D.J., Stewart, B.C., Maycock, T.K., Eds.; USA Global Change Research Program: Washington, DC, USA, 2017; Volume I, pp. 35–72. [Google Scholar] [CrossRef] [Green Version]
- Knisley, C.B.; Kippenhan, M.; Brzoska, D. Conservation status of United States tiger beetles. Terr. Arthropod Rev. 2014, 7, 93–145. [Google Scholar] [CrossRef] [Green Version]
- Duran, D.P.; Herrmann, D.P.; Roman, S.J.; Gwiazdowski, R.A.; Drummond, J.; Hood, G.R.; Egan, S.P. Cryptic diversity in the North American Dromochous tiger beetles (Coleoptera: Carabidae: Cicindelinae): A congruence-based method for species discovery. Zool. J. Linn. Soc. 2019, 186, 250–285. [Google Scholar] [CrossRef]
- Duran, D.P.; Laroche, R.A.; Gough, H.M.; Gwiazdowski, R.A.; Knisley, C.B.; Herrmann, D.P.; Roman, S.J.; Egan, S.P. Geographic Life History Differences Predict Genomic Divergence Better than Mitochondrial Barcodes or Phenotype. Genes 2020, 11, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erwin, T.L.; Pearson, D.L. A Treatise on the Western Hemisphere Caraboidea (Coleoptera). Their Classification, Distributions, and Ways of Life; Carabidae-Nebriiformes 2-Cicindelitae; Pensoft Series Faunistica 84; Pensoft Publishers: Sofia, Bulgaria, 2008; Volume II, 400p. [Google Scholar]
- Rivalier, É. Démembrement du Genre Cicindela Linné, II. Faune Am. Rev. Fr. Entomol. 1954, 21, 248–268. [Google Scholar]
- Rivalier, É. Remarques sur la tribu des Cicindelini (Col. Cicindelidae) et sa subdivision en soustribus. Nouv. Rev. Entomol. 1971, 1, 135–143. [Google Scholar]
- Wiesner, J. Verzeichis der Sandlaufkäfer der Welt; Verlag, Erna Bauer: Keltern-Weiler, Germany, 1992; 366p. [Google Scholar]
- Gough, H.M.; Duran, D.P.; Kawahara, A.Y.; Toussaint, E.F.A. A comprehensive molecular phylogeny of tiger beetles (Coleoptera, Carabidae, Cicindelinae). Syst. Èntomol. 2018, 44, 305–321. [Google Scholar] [CrossRef]
- Dejean, P.F.M.A.; Aubé, C. Species Général des Coléoptères de la Collection de M. le Comte Dejean; Tome Cinquième, Méquignon-Marvis: Paris, France, 1831; Volume VIII, 883p. [Google Scholar]
- Normandeau Associates, Inc. ILP Study 26 Cobblestone and Puritan Tiger Beetle Survey Study Report: In Support of Federal Energy Regulatory Commission Relicensing of: Wilder Hydroelectric Project (FERC Project No. 1892-026) Bellows Fall, Hydroelectric Project (FERC Project No. 1855-045), Vernon Hydroelectric Project (FERC Project No. 1904-073); Prepared for TransCanada Hydro Northeast Inc.; TransCanada Hydro Northeast Inc.: Concord, NH, USA, 2016; 65p. [Google Scholar]
- Leonard, J.G. Bell RT Northeastern Tiger Beetles. A Field Guide to Tiger Beetles of New England and Eastern Canada; CRC Press: Boca Raton, FL, USA, 1998; 192p. [Google Scholar]
- Hudgins, R.M. Habitat Selection, Dispersal and Detectability of Cobblestone Tiger Beetles (Cicindela marginipennis Dejean) along the Genesee River, New York. Master’s Thesis, State University of New York College at Brockport, Brockport, NY, USA, 2010. [Google Scholar]
- USFWS, USA Fish and Wildlife Service. Species Status Assessment (SSA) Report for the Cobblestone Tiger Beetle (Cicindela marginipennis) Version 1.1; USA Fish and Wildlife Service Northeast Region: Hadley, MA, USA, 2018. Available online: https://ecos.fws.gov/ServCat/DownloadFile/164797?Reference=111532 (accessed on 1 July 2020).
- Schlesinger, M.D.; Novak, P.G. Status and conservation of an imperiled tiger beetle fauna in New York State, USA. J. Insect Conserv. 2011, 15, 839–852. [Google Scholar] [CrossRef]
- Federal Register. Review of Invertebrate Wildlife for Listing as Endangered or Threatened Species. In Endangered and Threatened Wildlife and Plants; USA Fish and Wildlife Service: Washington, DC, USA, 1984; Volume 49, pp. 21664–21675. [Google Scholar]
- Federal Register. Twelve Species Not Warranted for Listing as Endangered or Threatened Species. In Endangered and Threatened Wildlife and Plants; U.S Fish and Wildlife Service: Washington, DC, USA, 2019; Volume 84, pp. 53336–53343. [Google Scholar]
- Gwiazdowski, R. Report on the First Observation of Mitochondrial Diversity, and Morphological Descriptions of Larvae for the Cobblestone Tiger Beetle; Cicindela marginipennis Dejean; Report Prepared for the New England Field office of the USA Fish and Wildlife Service; USA Fish and Wildlife Service: Concord, NH, USA, 2020; 20p. [Google Scholar]
- Ward, M.A.; Mays, J.D. New Records of The White Mountain Tiger Beetle, Cicindela ancocisconensis Harris (1852), and First Record of the Cobblestone Tiger Beetle, Cicindela marginipennis Dejean (1831) In Maine. Cicindela 2010, 42, 11–21. [Google Scholar]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, Weighting, and Phylogenetic Utility of Mitochondrial Gene Sequences and a Compilation of Conserved Polymerase Chain Reaction Primers. Ann. Èntomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Crozier, R.H.; Crozier, Y.C. The cytochrome b and ATPase genes of honeybee mitochondrial DNA. Mol. Biol. Evol. 1992, 9, 474–482. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A Cladistic Analysis of Phenotypic Associations with Haplotypes Inferred from Restriction Endonuclease Mapping and DNA Sequence Data. III. Cladogram Estimation. Genetics 1992, 132, 619–633. [Google Scholar] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MRBAYES 3.2: Efficient Bayesian phylogenetic inference and model selection across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shelford, V.E. Life-histories and larva habitats of the tiger beetles (Cicindelidae). J. Linn. Soc. (London) Zoo 1908, 30, 157–184. [Google Scholar] [CrossRef]
- Gwiazdowski, R.A.; Gillespie, S.; Weddle, R.; Elkinton, J.S. Laboratory Rearing of Common and Endangered Species of North American Tiger Beetles (Coleoptera: Carabidae: Cicindelinae). Ann. Entomol. Soc. Am. 2011, 104, 534–542. [Google Scholar] [CrossRef] [Green Version]
- Gwiazdowski, R.A.; Rodger Gwiazdowski, Advanced BioConsulting, LLC., 139 Spring St., Shrewsbury, MA 01545, USA. Personal communication. 2016–2019.
- Knisley, C.B.; Pearson, D.L. Biosystematics of larval tiger beetles of the Sulphur Springs Valley, Arizona. Trans. Am. Ent. Soc. 1984, 110, 465–551. [Google Scholar]
- Valenti, M.A. Synopsis of reported larval descriptions of tiger beetles (Coleoptera: Cicindelidae) from North America north of Mexico. Cicindela 1996, 28, 45–52. [Google Scholar]
- Hamilton, C.C. Studies on the morphology, taxonomy, an ecology of the larvae of Holarctic tiger-beetles (Family Cicindelidae). Proc. USNM 1925, 65, 1–87. [Google Scholar]
- Leffler, S.R. Tiger Beetles of the Pacific Northwest (Coleoptera: Cicindelidae). Ph.D. Thesis, University of Washington, Seattle, WA, USA, 1979. [Google Scholar]
- Willis, H.L. Bionomics and zoogeography of tiger beetles of saline habitats in the central United States. Univ. Kans. Sci. Bull. 1967, 47, 145–313. [Google Scholar]
- Gaumer, G.C. The Variation and Taxonomy of Cicindela formosa Say (Coleoptera: Cicindelidae). Ph.D. Thesis, Texas A & M University, College Station, TX, USA, 1977. [Google Scholar]
- Horn, G.H. Descriptions of the Larvae of North American Genera of Cicindelidae, also of Dicaelus with a Note on Rhynchophorus. Trans. Am. Ent. Soc. 1878, 7, 28–40. [Google Scholar] [CrossRef]
- Beatty, D.R.; Knisley, C.B. A description of the larval stages of Cicindela rufiventris Dejean (Coleoptera:Cicindelidae). Cicindela 1982, 13, 1–17. [Google Scholar]
- Kaulbers, M.M.; Freitag, R. A description of the third instar larva of Cicindela denikei. Cicindela 1993, 25, 45–48. [Google Scholar]
- Spanton, T.G. The Cicindela sylvatica group: Geographical variation and classification of the neartic taxa, and constructed phylogeny and geographic history of the species. Quaest. Entomol. 1988, 24, 51–161. [Google Scholar]
- Willis, H.L. Description of the larva of Cicindela patruela. Cicindela 1980, 12, 49–56. [Google Scholar]
- Knisley, C.B.; Schultz, T.D. The Biology of Tiger Beetles and a Guide to the Species of the South Atlantic States; Virginia Museum of Natural History: Martinsville, VA, USA, 1997. [Google Scholar]
- Gwiazdowski, R.A.; Department of Environmental Conservation, University of Massachusetts, Amherst, MA 01002, USA. Personal communication. 2019–2020.
- Pineda, P.M.; Kondratieff, B.C. The Larvae of Cicindela theatina (Coleoptera: Cicindelidae); A Regional North American Sand Dune Endemic. Entomol. News 2002, 113, 163–172. [Google Scholar]
- Barraclough, T.G.; Vogler, A.P. Recent Diversification Rates in North American Tiger Beetles Estimated from a Dated mtDNA Phylogenetic Tree. Mol. Biol. Evol. 2002, 19, 1706–1716. [Google Scholar] [CrossRef] [Green Version]
- Duran, D.P. Speciation and Diversification in The North American Tiger Beetles Of The Cicindela sylvatica Group: Morphological Variation and an Ecophylogeographic Approach. Ph.D. Thesis, Vanderbilt University, Nashville, TN, USA, 2010. [Google Scholar]
- Diogo, A.; Vogler, A.; Gimenez, A.; Gallego, D.; Galián, J. Conservation Genetics of Cicindela Deserticoloides, an Endangered Tiger Beetle Endemic to Southeastern Spain. J. Insect Conserv. 1999, 3, 117–123. [Google Scholar] [CrossRef]
- Doorenweerd, C.; Jose, M.S.; Barr, N.; Leblanc, L.; Rubinoff, D. Highly Variable COI Haplotype Diversity between Three Species of Invasive Pest Fruit Fly Reflects Remarkably Incongruent Demographic Histories. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gwiazdowski, R.A.; Elkinton, J.; Sremac, M.; Dewaard, J.R. Phylogeographic Diversity of the Winter Moths Operophtera brumata and O. bruceata (Lepidoptera: Geometridae) in Europe and North America. Ann. Èntomol. Soc. Am. 2013, 106, 143–151. [Google Scholar] [CrossRef]
- Luo, Y.; Agnarsson, I. Global MtDNA Genetic Structure and Hypothesized Invasion History of a Major Pest of Citrus, Diaphorina citri (Hemiptera: Liviidae). Ecol. Evol. 2018, 8, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knisley, C.B.; Gowan, C.; Wirth, C. Effects of soil moisture, vegetation and food on adult activity, oviposition and larval development in the tiger beetle, Cicindela albissima Rumpp. J. Insect Conserv. 2018, 22, 443–449. [Google Scholar] [CrossRef]
- Gwiazdowski, R.A. Interim Report on observed natural oviposition for the Puritan tiger beetle, Ellipsoptera (Cicindela) puritana, in southern Connecticut. Unpublished report to the USA Fish and Wildlife Service. New England Field Office: Concord, NH, USA, 2020; 9p. [Google Scholar]
- Cunningham, A.A.; Daszak, P. Extinction of a Species of Land Snail Due to Infection with a Microsporidian Parasite. Conserv. Biol. 1998, 12, 1139–1141. [Google Scholar] [CrossRef]
- Goldstein, P.Z.; National Museum of Natural History, Washington, DC, USA. Personal Communication, 2018.
- Cornelisse, T.M.; Vasey, M.C.; Holl, K.D.; Letourneau, D.K. Artificial bare patches increase habitat for the endangered Ohlone tiger beetle (Cicindela ohlone). J. Insect Conserv. 2012, 17, 17–22. [Google Scholar] [CrossRef]
- Gwiazdowski, R.A. Puritan Tiger Beetle (PTB) Recovery Project 2018 Annual Report; Unpublished report to the USA Fish and Wildlife Service, Conte National Wildlife Refuge; USA Fish and Wildlife Service: Sunderland, MA, USA, 2019; 32p. [Google Scholar]
- Dunn, G.A.; Wilson, D. Cicindela marginipennis in New Hampshire. Cicindela 1979, 11, 49–56. [Google Scholar]
- Nothnagle, P. Survey of the White River for the Cobblestone Tiger Beetle, Cicindela marginipennis; Report Prepared for the Nongame and Natural Heritage Program, Vermont Fish and Wildlife Department; Vermont Fish and Wildlife Department: Waterbury, VT, USA, 1995; 17p. [Google Scholar]
- Kritsky, G.; Cortright, B.; Duennes, M.; Smith, J.; Pierce, S. The Status of Cicindela marginipennis (Coleoptera: Carabidae) In Southeastern Indiana. Proc. Indiana Acad. Sci. 2009, 118, 139–142. [Google Scholar]
- Hudgins, R.; Norment, C.; Schlesinger, M.D.; Novak, P.G. Habitat Selection and Dispersal of the Cobblestone Tiger Beetle (Cicindela marginipennis Dejean) along the Genesee River, New York. Am. Midl. Nat. 2011, 165, 304–318. [Google Scholar] [CrossRef]
- Hudgins, R.M.; Norment, C.; Schlesinger, M.D. Assessing detectability for monitoring of rare species: A case study of the cobblestone tiger beetle (Cicindela marginipennis Dejean). J. Insect Conserv. 2011, 16, 447–455. [Google Scholar] [CrossRef]
- Mawdsley, J.R. Use of simple remote sensing tools to expedite surveys for rare tiger beetles (Insecta: Coleoptera: Cicindelidae). J. Insect Conserv. 2007, 12, 689–693. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Rearing Event |
|---|---|
| 21 July 2019 | Adults field collected and brought into the lab |
| 23 July 2019 | Adults moved into oviposition bins with native soil |
| 4 August 2019 | First oviposition |
| 26 August 2019 | First hatch (1st instar) |
| 26 September 2019 | First 2nd instar |
| 6 January 2020 | Moved to larger aquaria |
| 22 January 2020 | First 3rd instar |
| 30 March 2020 | Most remaining larvae in 3rd instar |
| Max. 1st Instar | Sex | Days Alive | Collected | Deceased | Accession # |
|---|---|---|---|---|---|
| 34 | Female | 54 | 21 July | 13 September | VT1 |
| 32 | Female | 75 | 21 July | 4 October | NH1 |
| 11 | Female | 36 | 21 July | 26 August | VT3 |
| 10 | Female | 33 | 21 July | 23 August | NH3 |
| 9 | Female | 31 | 21 July | 21 August | NH2 |
| 8 | Female | 44 | 21 July | 3 September | VT2 |
| 20 | Female | 19 | 20 September | 9 October | NH5.1 |
| 5 | Female | 14 | 20 September | 4 October | NH4 |
| 3 | Female | 8 | 20 September | 28 September | NH5 |
| N/A | Male | 70 | 21 July | 29 September | NH3 |
| N/A | Male | 69 | 21 July | 28 September | NH1 |
| N/A | Male | 54 | 21 July | 13 September | VT1 |
| N/A | Male | 38 | 21 July | 28 August | VT3 |
| N/A | Male | 36 | 21 July | 26 August | VT2 |
| N/A | Male | 8 | 20 September | 28 September | NH4 |
| Region | Alabama | ME/NH | Northeast | WV |
|---|---|---|---|---|
| Alabama | 99.31 | 3.47 | 3.47 | 3.20 |
| ME/NH | 1.72 | 100.00 | 0.58 | 0.66 |
| Northeast | 1.50 | 0.33 | 99.94 | 0.87 |
| WV | 1.45 | 0.65 | 0.49 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gwiazdowski, R.A.; Andersen, J.C.; Knisley, C.B.; Griffin, B.P.; Elkinton, J.S. First Account of Phylogeographic Variation, Larval Characters, and Laboratory Rearing of the Endangered Cobblestone Tiger Beetle Cicindelidia marginipennis, Dejean, 1831 with Observations of Their Natural History. Insects 2020, 11, 708. https://doi.org/10.3390/insects11100708
Gwiazdowski RA, Andersen JC, Knisley CB, Griffin BP, Elkinton JS. First Account of Phylogeographic Variation, Larval Characters, and Laboratory Rearing of the Endangered Cobblestone Tiger Beetle Cicindelidia marginipennis, Dejean, 1831 with Observations of Their Natural History. Insects. 2020; 11(10):708. https://doi.org/10.3390/insects11100708
Chicago/Turabian StyleGwiazdowski, Rodger A., Jeremy C. Andersen, C. Barry Knisley, Brian P. Griffin, and Joseph S. Elkinton. 2020. "First Account of Phylogeographic Variation, Larval Characters, and Laboratory Rearing of the Endangered Cobblestone Tiger Beetle Cicindelidia marginipennis, Dejean, 1831 with Observations of Their Natural History" Insects 11, no. 10: 708. https://doi.org/10.3390/insects11100708