Identification of Entomopathogenic Fungi as Naturally Occurring Enemies of the Invasive Oak Lace Bug, Corythucha arcuata (Say) (Hemiptera: Tingidae)




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Fungi Isolation
2.2. Morphological Characterization
2.3. DNA Isolation, Polymerase Chain Reaction (PCR) Amplification and Sequencing
2.4. Phylogenetic Analysis
3. Results
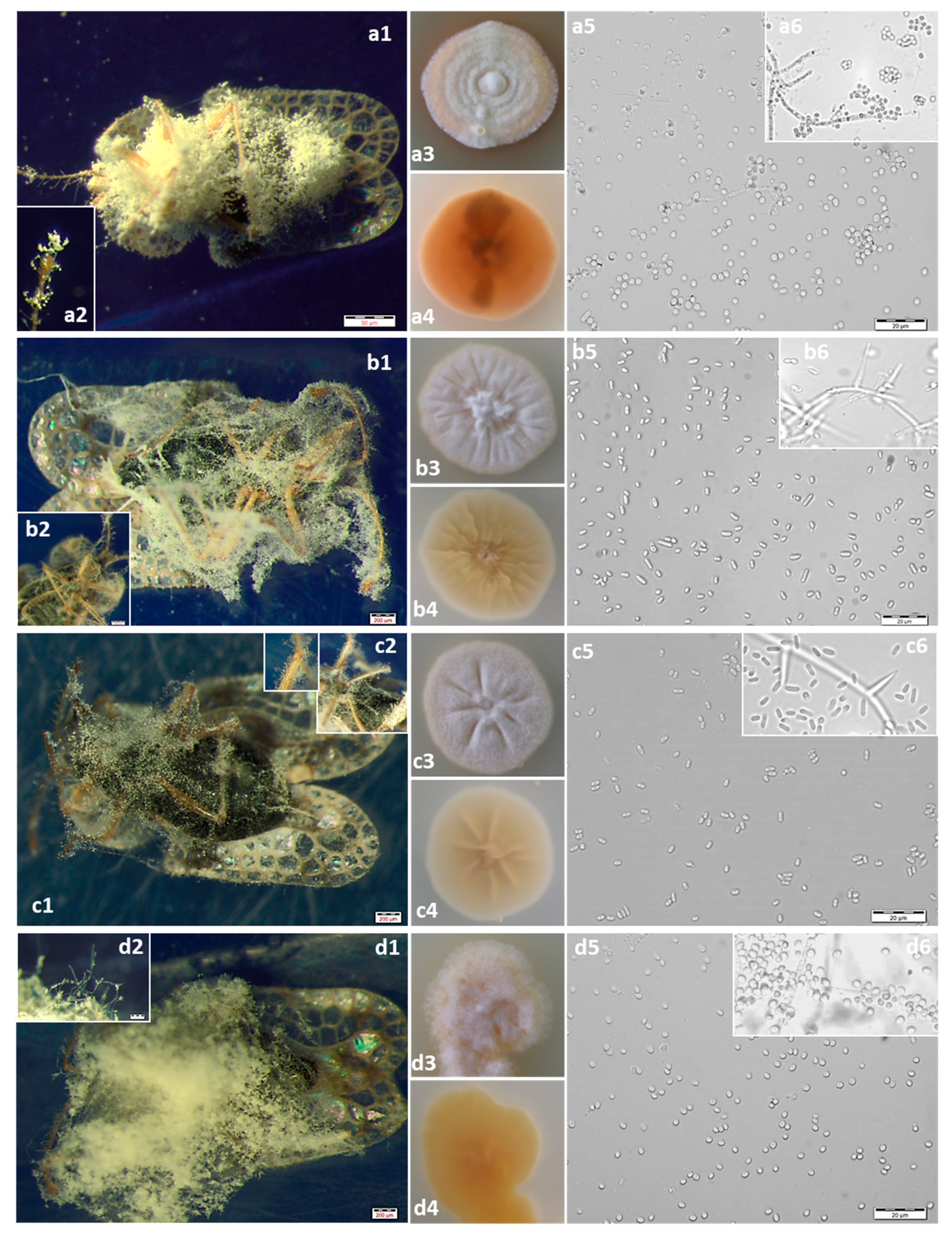
3.1. Isolation and Morphological Identification of Fungi
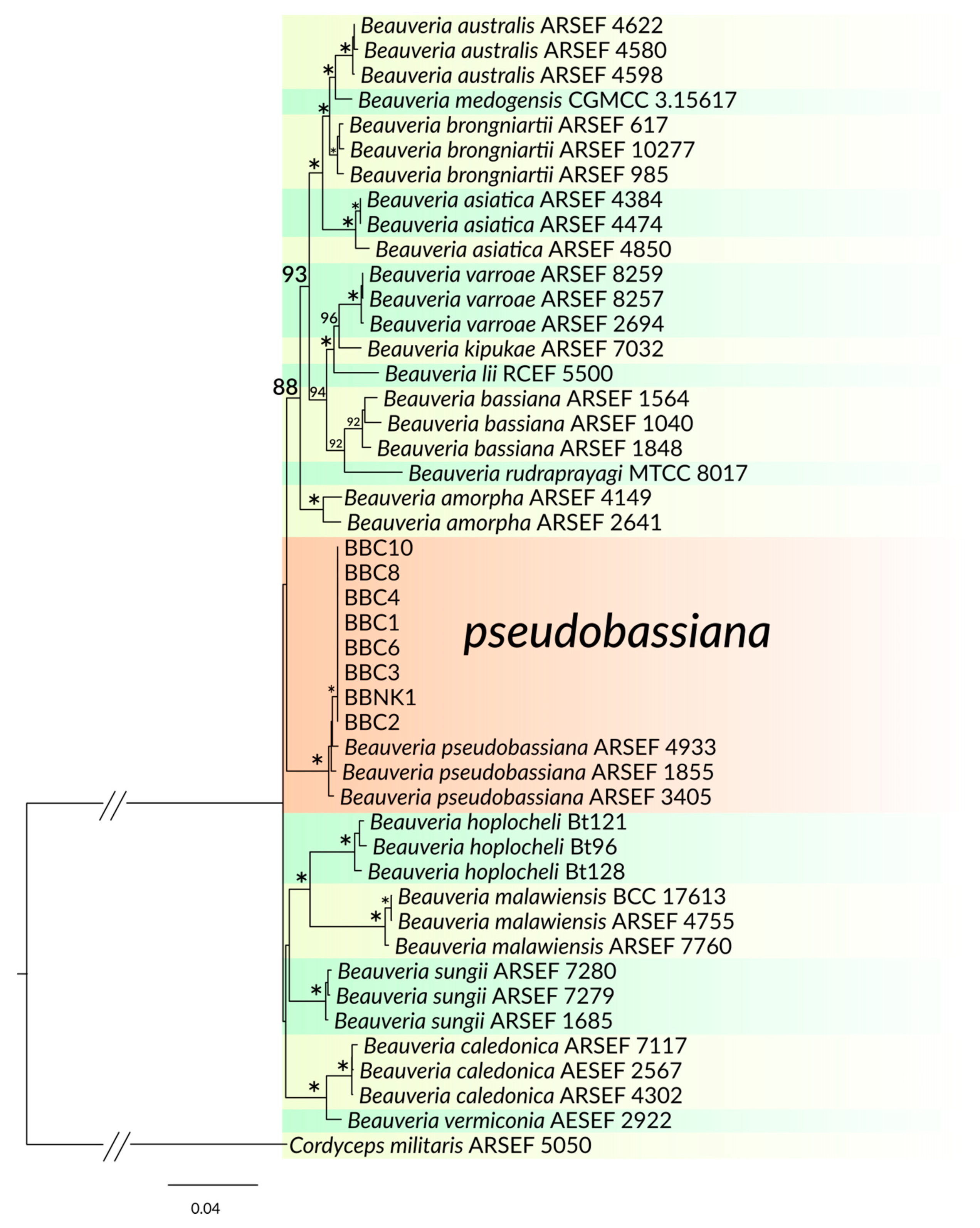
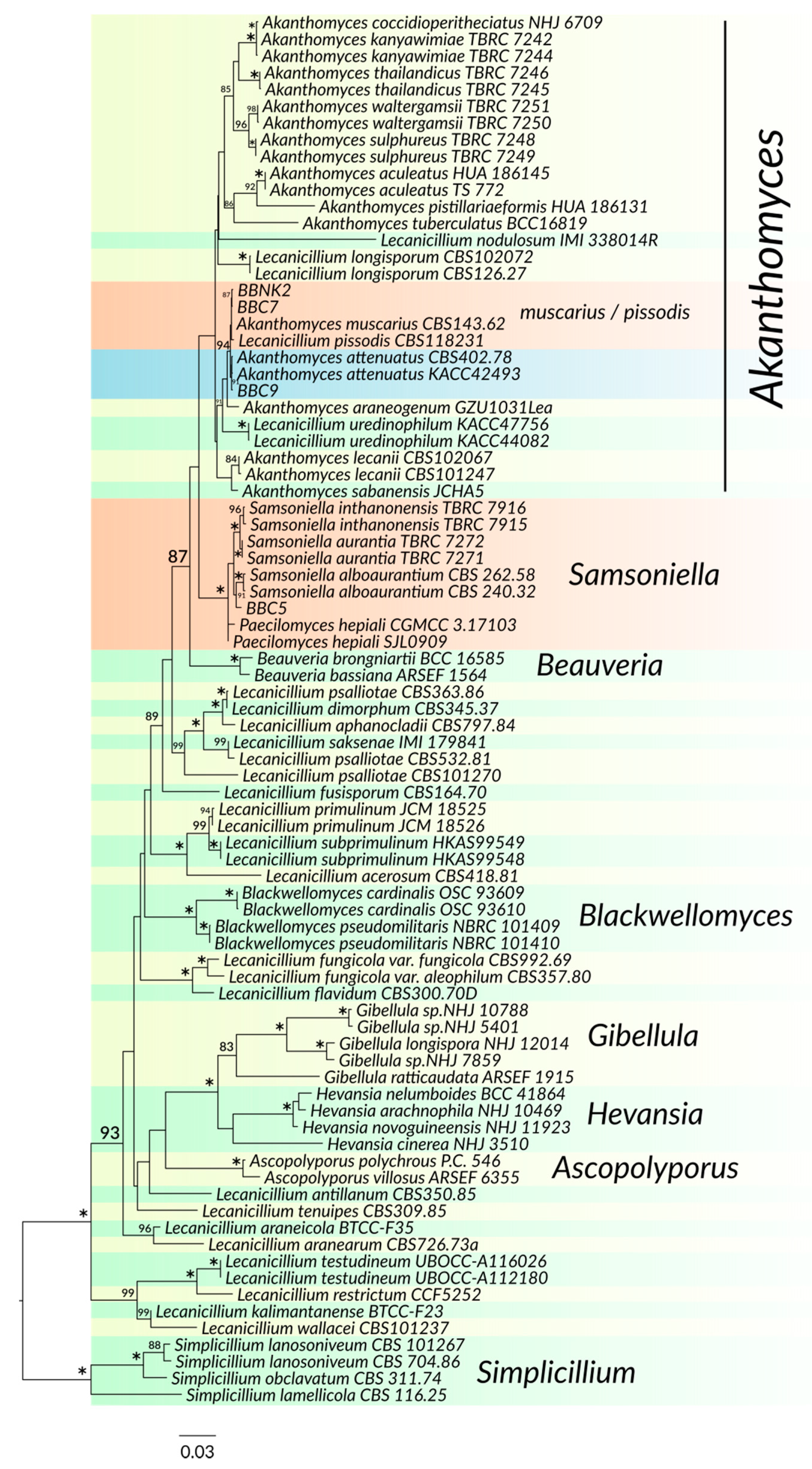
3.2. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Drake, C.J.; Ruhoff, F.A. Lacebugs of the world, a Catalog (Hemiptera: Tingidae). Bull. US Natl. Mus. 1965, 243, 1–634. [Google Scholar] [CrossRef]
- Rabitsch, W. Alien true bugs of Europe (Insecta: Hemiptera: Heteroptera). Zootaxa 2008, 1827, 1–44. [Google Scholar] [CrossRef] [Green Version]
- Bernardinelli, I.; Zandigiacomo, P. Prima segnalazione di Corythucha arcuata (Say) (Heteroptera, Tingidae) in Europa. Inf. Fitopatol. 2000, 50, 47–49. [Google Scholar]
- Mutun, S. First report of the oak lace bug, Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae) from Bolu, Turkey. Isr. J. Zool. 2003, 33, 263–268. [Google Scholar]
- Forster, B.; Giacalone, I.; Moretti, M.; Dioli, P.; Wermelinger, B. Die amerikanische Eichennetzwanze Corythucha arcuata (Say) (Heteroptera, Tingidae) hat die Süsschweiz erreicht. Mitt. Schweiz. Entomol. Ges. 2005, 78, 317–323. [Google Scholar]
- Samin, N.; Linnavuori, R.E. A contribution to the Tingidae (Heteroptera) from north and northwestern Iran. Entomofauna 2011, 32, 373–380. [Google Scholar]
- Dobreva, M.; Simov, N.; Georgiev, G.; Mirchev, P.; Georgieva, M. First Record of Corythucha arcuata (Say) (Heteroptera: Tingidae) on the Balkan Peninsula. Acta Zool. Bulg. 2013, 3, 409–412. [Google Scholar]
- Hrašovec, B.; Posarić, D.; Lukić, I.; Pernek, M. Prvi nalaz hrastove mrežaste stjenice (Corythucha arcuata) u Hrvatskoj. Sumar. List 2013, 137, 499–503. [Google Scholar]
- Csóka, G.; Hirka, A.; Somlyai, M. A tölgy csipkéspoloska (Corythuca arcuata Say, 1832—Hemiptera, Tingidae) elsô észlelése Magyarországon. Növényvédelem 2013, 49, 293–296. [Google Scholar]
- Pap, P.; Drekić, M.; Poljaković-Pajnik, L.; Marković, M.; Vasić, V. Monitoring zdravstvenog stanja šuma na teritoriji Vojvodine u 2015. godini. Topola Poplar 2015, 188, 117–133. [Google Scholar]
- Don, I.; Don, C.D.; Sasu, L.R.; Vidrean, D.; Brad, M.L. Insect pests on the trees and shrubs from the Macea Botanical Garden. Studia Univ. Vasile Goldis Arad 2016, 11, 23–28. [Google Scholar]
- Chireceanu, C.; Theodoru, A.; Chiriloaie, A. First record of oak lace bug Corythucha arcuata (Tingidae: Heteroptera). In Proceedings of the 7th ESENIAS Workshop with Scientific Conference Networking and Regional Cooperation towards Invasive Alien Species Prevention and Management in Europe, Sofia, Bulgaria, 28–30 March 2017. [Google Scholar]
- Shchurov, V.I.; Bondarenko, A.S.; Skvortsov, M.M.; Shchurova, A.V. The alien invasive forest insect pests for the first time revealed in forest ecosystems of the Northwest Caucasus in 2014–2016. In The Kataev Memorial Readings—IX Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems, Proceedings of the International Conference, Saint Petersburg, Russia, 23–25 November 2016; Musolin, D.L., Selikhovkin, A.V., Eds.; Saint Petersburg State Forest Technical University: Saint Petersburg, Russia, 2016. [Google Scholar]
- Jurc, M.; Jurc, D. The first record and the beginning of the spread of oak lace bug, Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae), in Slovenia. Sumar. List 2017, 141, 485–488. [Google Scholar] [CrossRef] [Green Version]
- Dautbašić, M.; Zahirović, K.; Mujezinović, O.; Margaletić, J. First record of oak lace bug (Corythucha arcuata) in Bosnia and Herzegovina. Sumar. List 2018, 142, 179–181. [Google Scholar] [CrossRef] [Green Version]
- Streito, J.C.; Balmès, V.; Aversenq, P.; Weill, P.; Chapin, E.; Clément, M.; Piednoir, F. Corythucha arcuata (Say, 1832) et Stephanitis lauri Rietschel, 2014, deux espèces invasives nouvelles pour la faune de France (Hemiptera, Tingidae). L’Entomologiste 2018, 74, 133–136. [Google Scholar]
- Zúbrik, M.; Gubka, A.; Rell, S.; Kunca, A.; Vakula, J.; Galko, J.; Nikolov, C.; Leontovyč, R. First record of Corythucha arcuata in Slovakia—Short Communication. Plant. Prot. Sci. 2019, 55, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Sallmannshofer, M.; Ette, M.S.; Hinterstoisser, W.; Cech, T.L.; Hoch, G. Erstnachweis der Eichennetzwanze, Corythucha arcuata, in Österreich. Aktuell 2019, 66, 1–6. [Google Scholar]
- György Csóka, G.; Hirka, A.; Mutun, S.; Glavendeki, M.; Mikó, A.; Szőcs, L.; Paulin, M.; Csaba Béla Eötvös, C.B.; Gáspár, C.; Csepelényi, M.; et al. Spread and potential host range of the invasive oak lace bug [Corythucha arcuata (Say, 1832)—Heteroptera: Tingidae] in Eurasia. Agr. For. Entomol. 2019, 22, 61–74. [Google Scholar] [CrossRef]
- Bernardinelli, I. Potential host plants of Corythucha arcuata (Het., Tingidae) in Europe: A laboratory study. J. Appl. Entomol. 2006, 130, 480–484. [Google Scholar] [CrossRef]
- Connell, W.A.; Beacher, J.H. Life history and control of the oak lace bug. Bull. Univ. Del. Agric. Exp. Stn. 1947, 265, 28. [Google Scholar]
- Mutun, S.; Ceyhan, Z.; Sözen, C. Invasion by the oak lace bug, Corythucha arcuata (Say) (Heteroptera: Tingi¬dae), in Turkey. Turk. J. Zool. 2009, 49, 323–324. [Google Scholar] [CrossRef]
- UK Government Department for Environment, Food and Rural Affairs (Defra). Rapid Pest Risk Analysis (PRA) for: Corythucha arcuata. November 2018; UK government Department for Environment, Food and Rural Affairs (Defra): London, UK, 2018.
- Dutto, M.; Bertero, M. Dermatosis caused by Corythuca ciliata (Say, 1932) (Heteroptera, Tingidae). diagnostic and clinical aspects of an unrecognized pseudoparasitosis. J. Prev. Med. Hyg. 2009, 54, 57–59. [Google Scholar]
- Pernek, M.; (Croatian Forest Research Institute, Jastrebarsko, Croatia). Personal communication, 2019.
- Ciceoi, R.; Radulovici, A. Facultative blood-sucking lace bugs, Corythucha sp. in Romania. In Joint ESENIAS and DIAS Scientific Conference and 8th ESENIAS Workshop, Book of Abstracts Management and Sharing of IAS Data to Support Knowledge Based Decision Making at Regional Level, Bucharest, Romania, 26–28 September 2018; Anastasiu, P., Trichkova, T., Uludag, A., Tomov, R., Eds.; Editura Universitatii din Bucuresti: Bucharest, Romania, 2018. [Google Scholar]
- Dreistadt, S.H.; Perry, E.J. Lace Bugs: Integrated pest management for home gardeners and lascape professionals. Pest Notes Univ. Calif. Agric. Nat. Resour. Bull. 2006, 74, 1–4. [Google Scholar]
- Dreistadt, S.H.; (UC Statewide IPM Program, Agriculture & Natural Resources, University of California, Davis, CA, USA). Personal communication, 2019.
- Dara, S.K.; Motlava, C.; Barta, M. Microbial control of invasive forest pests with entomopathogenic fungi: A review of the current situation. Insects 2019, 10, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khetan, S.K. Microbial Pest Control; Marcel Dekker: New York, NY, USA, 2001; p. 320. [Google Scholar]
- Augustyniuk-Kram, A.; Kram, K.J. Forest Ecosystems—More than Just Trees, Entomopathogenic Fungi as an Important Natural Regulator of Insect Outbreaks in Forests (Review); Blanco, J.A., Lo, Y.H., Eds.; InTech: Rijeka, Croatia, 2012; pp. 265–294. [Google Scholar]
- Kovač, M.; Lacković, N.; Pernek, M. Effect of Beauveria bassiana fungal infection on survival and feeding behavior of pine-tree lappet moth (Dendrolimus pini L.). Forests 2020, 11, 974. [Google Scholar]
- Sönmez, E.; Demirbag, Z.; Demir, I. Pathogenicity of selected entomopathogenic fungal isolates against the oak lace bug, Corythucha arcuata Say. (Hemipteran: Tingidae), under controlled conditions. Turk. J. Agric. 2016, 40, 715–722. [Google Scholar]
- Kovač, M.; Pernek, M. Field research on natural infestation of entomopathogenic fungi on overwintering Corythucha arcuata (Say) (Hemiptera: Tingidae). (manuscript in preparation).
- Greenfield, M.; Gómez-Jiménez, M.I.; Ortiz, V.; Vega, F.E.; Kramer, M.; Parsa, S. Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biol. Control 2016, 95, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E.P. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Humber, R.A. Manual of Techniques in Invertebrate Pathology, Identification of Entomopathogenic Fungi; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 151–187. [Google Scholar]
- Inglis, G.D.; Enkerli, J.; Goettel, M.S. Manual of Techniques in Invertebrate Pathology. Laboratory Techniques Used for Entomopathogenic Fungi: Hypocreales Chapter VII; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 189–253. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.H.; Hyten, A.S.; Spatafora, J.W. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am. J. Bot. 2002, 89, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to multilocus phylogenetic studies of the euascomycetes (Pezizomycotina, Fungi) with special emphasis on the lichen-forming Acarosporaceae and evolution of polyspory. Mol. Phylogenet. Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Posada, F.; Buckley, E.P.; Infante, F.; Castillo, A.; Vega, F.E. Phylogenetic origins of African and Neotropical Beauveria bassiana s.l. pathogens of the coffee berry borer, Hypothenemus hampei. J. Invertebr. Pathol. 2006, 93, 11–21. [Google Scholar] [CrossRef]
- Imoulan, A.; Li, Y.; Wang, W.J.; El Meziane, A.; Yao, Y.J. New record of Beauveria pseudobassiana from Morocco. Mycotaxon 2016, 131, 913–923. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Zhi, J.R.; Ye, M.; Zhang, Z.Y.; Yue, W.B.; Zou, X. Lecanicillium cauligalbarum sp. nov. (Cordycipitaceae, Hypocreales), a novel fungus isolated from a stemborer in the Yao Ren National Forest Mountain Park, Guizhou. MycoKeys 2018, 43, 59–74. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 23–69. [Google Scholar]
- Barta, M.; Lalík, M.; Rell, S.; Kunca, A.; Horáková, M.K.; Mudronceková, S.; Galko, J. Hypocrealean fungi associated with Hylobius abietis in Slovakia, their virulence against weevil adults and effect on feeding damage in laboratory. Insects 2019, 10, 634. [Google Scholar] [CrossRef] [Green Version]
- Schemmer, R.; Chládeková, P.; Medo, J.; Barta, M. Natural prevalence of entomopathogenic fungi in hibernating pupae of Cameraria ohridella (Lepidoptera: Gracillariidae) and virulence of selected isolates. Plant Prot. Sci. 2016, 52, 199–208. [Google Scholar]
- Medo, J.; Michalko, J.; Medová, J.; Cagáň, L. Phylogenetic structure and habitat associations of Beauveria species isolated from soils in Slovakia. J. Invertebr. Pathol. 2016, 140, 46–50. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, G.S. The genera Beauveria, Isaria, Tritirachium, and Acrodontium gen. nov. Stud. Mycol. 1972, 1, 1–41. [Google Scholar]
- Strasser, H.; Vey, A.; Butt, T.M. Are there any risks in using entomopathogenic fungi for pest control, with particular reference to the bioactive metabolites of Metarhizium, Tolypocladium and Beauveria species? Biocontrol Sci. Techn. 2000, 10, 717–735. [Google Scholar] [CrossRef]
- Goettel, M.S.; Koike, M.; Kim, J.J.; Aiuchi, D.; Shinya, R.; Brodeur, J. Potential of Lecanicillium spp. for management of insects, nematodes and plant diseases. J. Invertebr. Pathol. 2008, 98, 256–261. [Google Scholar] [CrossRef]
- Kope, H.H.; Leal, I. A new species of Lecanicillium isolated from the white pine weevil. Mycotaxon 2005, 94, 331–340. [Google Scholar]
- Chiriví-Salomón, J.S.; Danies, G.; Restrepo, S.; Sanjuan, T. Lecanicillium sabanense sp. nov. (Cordycipitaceae) a new fungal entomopathogen of coccids. Phytotaxa 2015, 234, 63–74. [Google Scholar]
- Kepler, R.M.; Luangsa-ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef]
- Wang, W.J.; Li, Y.; Wang, X.L.; Kirk, P.M.; Yao, Y.J. Neotypification of Paecilomyces hepiali (Hypocreales). Taxon 2015, 64, 147–150. [Google Scholar] [CrossRef]
- Dai, R.Q.; Shen, C.Y.; Li, X.M.; Lan, J.L.; Lin, S.F.; Shao, A.J. Response to neotypification of Paecilomyces hepiali (Hypocreales) (Wang et al. 2015). Taxon 2018, 67, 784–786. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A. Paecilomyces and some allied hyphomycetes. Stud. Mycol. 1974, 6, 1–119. [Google Scholar]
- Mongkolsamrit, S.; Noisripoom, W.; Thanakitpipattana, D.; Wutikhun, T.; Spatafora, J.W.; Luangsa-ard, J. Disentangling cryptic species with Isaria-like morphs in Cordycipitaceae. Mycologia 2018, 110, 230–257. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Locus | Forward | Reverse | Source |
|---|---|---|---|
| ITS | ITS1F | ITS4 | Gardes and Bruns, 1993 [40]; White et al., 1990 [41] |
| SSU | NS1 | NS6 | White et al., 1990 [41] |
| LSU | LR0R | LR5 | Vilgalys, unpublished; Vilgalys and Hester, 1990 [42] |
| TEF | EF1-983F | EF1-2218R | Rehner and Buckley, 2005 [36] |
| RPB1 | CRPB1 | RPB1-Cr | Castlebury et al., 2004 [43]; Matheny et al., 2002 [44] |
| RPB2 | RPB2-5F1 | RPB2-7cR | Sung et al., 2007 [45]; Reeb et al., 2004 [46] |
| Bloc | B5.1F | B3.1R | Rehner et al., 2006 [47] |
| Isolate | Species | Colonies | Conidiogenous Cell | Conidial Shape and Size |
|---|---|---|---|---|
| BBC1 BBC2 BBC3 BBC4 BBC6 BBC8 BBC10 BBNK1 | Beauveria pseudobassiana | white first and becoming yellowish-brown or pale yellow, aerial mycelium dense and farinaceous during sporulation, sometimes producing exudate, slightly changing color of medium to light purple; reverse side yellowish-brown with white margin | conidiophores solitary but usually consisting of dense spherical lateral clusters of globose to flask-shaped conidiogenous cells with an elongating sympodial denticulate rachis giving a zig-zag appearance, 3−6 × 1.3−2.5 μm | globose to subglobose, rarely ellipsoid, 1-celled, hyaline, smooth-walled; 2.5−3.7 × 1.4−3.1 μm |
| BBC5 | Samsoniella alboaurantium | white to light orange, cottony and floccose; reverse side deep yellow | conidiophores biverticillate with whorls of flask shaped phialides, 5.7–10 × 1.5–3 μm | conidia lemonshaped, 1-celled, hyaline, smooth walled, 2–3.5 × 1.9–3.5 μm, sometimes in chains |
| BBC7 BBNK2 | Lecanicillium pissodis | white; reverse side cream to pale yellow | phialides solitary, or 2 to 3 in whorls, 12.6–38 × 1–2 μm | formed in globose droplets containing up to more than 50 spores, cylindrical to oval, variable in size and shape, 2.6–7.2 × 1.7–2.5 μm |
| BBC9 | Akanthomyces attenuates | white; reverse side yellowish-white | up to 3–5 per node, 9.3–15.8 × 1–2 μm | cylindrical with slightly narrowed base, sometimes oval, very variable in size and shape 2.3–6 × 1.5–2.9 μm |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of Entomopathogenic Fungi as Naturally Occurring Enemies of the Invasive Oak Lace Bug, Corythucha arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. https://doi.org/10.3390/insects11100679
Kovač M, Gorczak M, Wrzosek M, Tkaczuk C, Pernek M. Identification of Entomopathogenic Fungi as Naturally Occurring Enemies of the Invasive Oak Lace Bug, Corythucha arcuata (Say) (Hemiptera: Tingidae). Insects. 2020; 11(10):679. https://doi.org/10.3390/insects11100679
Chicago/Turabian StyleKovač, Marta, Michał Gorczak, Marta Wrzosek, Cezary Tkaczuk, and Milan Pernek. 2020. "Identification of Entomopathogenic Fungi as Naturally Occurring Enemies of the Invasive Oak Lace Bug, Corythucha arcuata (Say) (Hemiptera: Tingidae)" Insects 11, no. 10: 679. https://doi.org/10.3390/insects11100679