Sex-Specific Wolbachia Infection Patterns in Populations of Polygraphus proximus Blandford (Coleoptera; Curculionidae: Scolytinae)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. DNA Extraction
2.3. PCR and Sequencing
2.4. Quantitative Real-Time PCR Assay
2.5. Data Analysis
3. Results
3.1. Wolbachia Infection Rates and Wolbachia Titer
3.1.1. Conventional PCR
3.1.2. Nested PCR
3.1.3. Estimation of Wolbachia Titer by qPCR
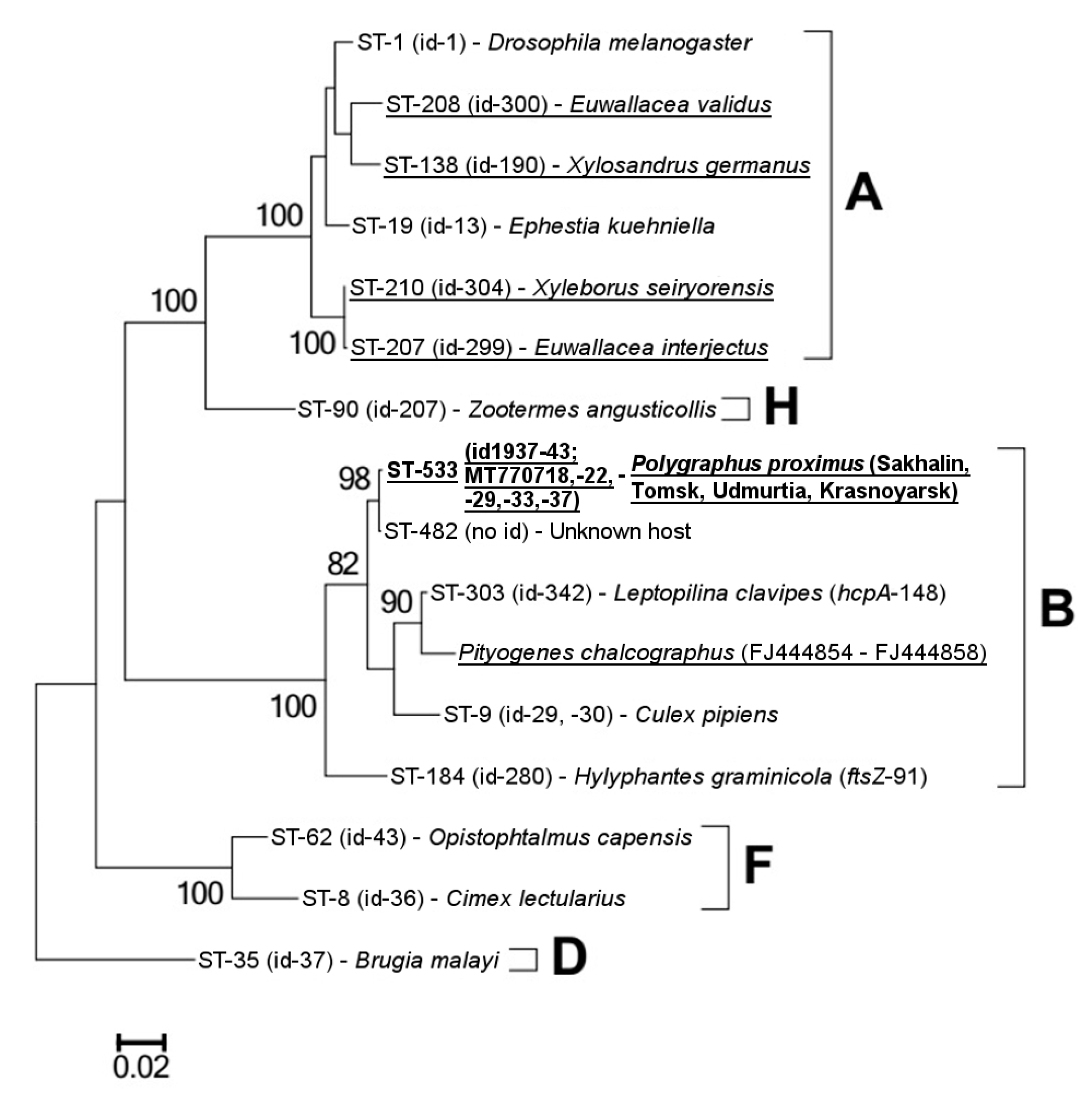
3.2. Characterization of Wolbachia Isolates and Mitochondrial DNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taylor, M.J.; Bandi, C.; Hoerauf, A. Wolbachia. Bacterial endosymbionts of filarial nematodes. Adv. Parasitol. 2005, 60, 245–284. [Google Scholar] [CrossRef] [PubMed]
- Haegeman, A.; Vanholme, B.; Jacob, J.; Vandekerckhove, T.T.; Claeys, M.; Borgonie, G.; Gheysen, G. An endosymbiotic bacterium in a plant-parasitic nematode: Member of a new Wolbachia supergroup. Int. J. Parasitol. 2009, 39, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werren, J.H. Biology of Wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vavre, F.; Fleury, F.; Lepetit, D.; Fouillet, P.; Boulétreau, M. Phylogenetic evidence for horizontal transmission of Wolbachia in host-parasitoid associations. Mol. Biol. Evol. 1999, 16, 1711–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, L.; Ayoub, N.A.; Hayashi, C.Y.; Russell, J.A.; Stahlhut, J.K.; Werren, J.H. Insight into the routes of Wolbachia invasion: High levels of horizontal transfer in the spider genus Agelenopsis revealed by Wolbachia strain and mitochondrial DNA diversity. Mol. Ecol. 2008, 17, 557–569. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Breinholt, J.W.; Kawahara, A.Y. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol. Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [Green Version]
- Ilinsky, Y.; Kosterin, O.E. Molecular diversity of Wolbachia in Lepidoptera: Prevalent allelic content and high recombination of MLST genes. Mol. Phylogenet. Evol. 2017, 109, 164–179. [Google Scholar] [CrossRef]
- Shaikevich, E.; Bogacheva, A.; Rakova, V.; Ganushkina, L.; Ilinsky, Y. Wolbachia symbionts in mosquitoes: Intra-and intersupergroup recombinations, horizontal transmission and evolution. Mol. Phylogenet. Evol. 2019, 134, 24–34. [Google Scholar] [CrossRef]
- De Barro, P.J.; Hart, P.J. Antibiotic curing of parthenogenesis in Eretmocerus mundus (Australian parthenogenic form). Entomol. Exp. Appl. 2001, 99, 225–230. [Google Scholar] [CrossRef]
- Dong, P.; Wang, J.J.; Hu, F.; Jia, F.X. Influence of Wolbachia infection on the fitness of the stored-product pest Liposcelis tricolor (Psocoptera: Liposcelididae). J. Econ. Entomol. 2007, 100, 1476–1481. [Google Scholar] [CrossRef]
- Mariño, Y.; Verle Rodrigues, J.; Bayman, P. Wolbachia affects reproduction and population dynamics of the coffee berry borer (Hypothenemus hampei): Implications for biological control. Insects 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, L.M.; Brownlie, J.C.; O'Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Ferreira, Á.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattel, J.; Martinez, J.; Jiggins, F.; Mouton, L.; Gibert, P. Wolbachia-mediated protection against viruses in the invasive pest Drosophila suzukii. Insect Mol. Biol. 2016, 25, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nouhuys, S.; Kohonen, M.; Duplouy, A. Wolbachia increases the susceptibility of a parasitoid wasp to hyperparasitism. J. Exp. Biol. 2016, 219, 2984–2990. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect–Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [Green Version]
- Starr, D.J.; Cline, T.W. A host–parasite interaction rescues Drosophila oogenesis defects. Nature 2002, 418, 76. [Google Scholar] [CrossRef]
- Clark, M.E.; Anderson, C.L.; Cande, J.; Karr, T.L. Widespread prevalence of Wolbachia in laboratory stocks and the implications for Drosophila research. Genetics 2005, 170, 1667–1675. [Google Scholar] [CrossRef] [Green Version]
- Ote, M.; Ueyama, M.; Yamamoto, D. Wolbachia protein TomO targets nanos mRNA and restores germ stem cells in Drosophila sex-lethal mutants. Curr. Biol. 2016, 26, 2223–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, M.; Shoubu, M.; Yamaguchi, D.; Yukawa, J. Defoliation and dieback of Abies firma (Pinaceae) trees caused by Parendaeus abietinus (Coleoptera: Curculionidae) and Polygraphus proximus (Coleoptera: Scolytidae) on Mount Unzen, Japan. Appl. Entomol. Zool. 2008, 43, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Baranchikov, Y.; Akulov, E.; Astapenko, S. Bark beetle Polygraphus proximus: A new aggressive Far Eastern invader on Abies species in Siberia and European Russia. In Proceedings of the 21st US Department of Agriculture Interagency Research Forum on Invasive Species 2010, Annapolis, MD, USA, 12–15 January 2010; pp. 64–65. [Google Scholar]
- Krivets, S.A.; Bisirova, E.M.; Kerchev, I.A.; Pats, E.N.; Chernova, N.A. Transformation of taiga ecosystems in the Western Siberian invasion focus of four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera: Curculionidae, Scolytinae). Russ. J. Biol. Inv. 2015, 6, 94–108. [Google Scholar] [CrossRef]
- Stauffer, C.; Van Meer, M.M.M.; Riegler, M. The presence of the protobacteria Wolbachia in European Ips typographus (Col., Scolytidae) populations and the consequences for genetic data. Proc. Ger. Soc. Gen. Appl. Entomol. 1997, 11, 709–711. [Google Scholar]
- Vega, F.E.; Benavides, P.; Stuart, J.A.; O’Neill, S.L. Wolbachia infection in the coffee berry borer (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 2002, 95, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Zchori-Fein, E.; Borad, C.; Harari, A.R. Oogenesis in the date stone beetle, Coccotrypes dactyliperda, depends on symbiotic bacteria. Physiol. Entomol. 2006, 31, 164–169. [Google Scholar] [CrossRef]
- Arthofer, W.; Markus, R.; Dimitrios, N.A.; Christian, S. Evidence for low-titre infections in insect symbiosis: Wolbachia in the bark beetle Pityogenes chalcographus (Coleoptera, Scolytinae). Environ. Microbiol. 2009, 11, 1923–1933. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Schuler, H.; Stauffer, C.; Lakatos, F.; Kajimura, H. Wolbachia endosymbionts in haplodiploid and diploid scolytine beetles (Coleoptera: Curculionidae: Scolytinae). Environ. Microbiol. Rep. 2016, 8, 680–688. [Google Scholar] [CrossRef]
- Baldo, L.; Hotopp, J.C.D.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [Green Version]
- Hurst, G.D.; Jiggins, F.M.; von der Schulenburg, J.H.G.; Bertrand, D.; West, S.A.; Goriacheva, I.I.; Zakharov, I.A.; Werren, J.H.; Stouthamer, R.; Majerus, M.E.N. Male–killing Wolbachia in two species of insect. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 735–740. [Google Scholar] [CrossRef] [Green Version]
- Richardson, M.F.; Weinert, L.A.; Welch, J.J.; Linheiro, R.S.; Magwire, M.M.; Jiggins, F.M.; Bergman, C.M. Population genomics of the Wolbachia endosymbiont in Drosophila melanogaster. PLoS Genet. 2012, 8, e1003129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilinsky, Y. Coevolution of Drosophila melanogaster mtDNA and Wolbachia genotypes. PLoS ONE 2013, 8, e54373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bykov, R.A.; Yudina, M.A.; Gruntenko, N.E.; Zakharov, I.K.; Voloshina, M.A.; Melashchenko, E.S.; Danilova, M.V.; Mazunin, I.O.; Ilinsky, Y.Y. Prevalence and genetic diversity of Wolbachia endosymbiont and mtDNA in Palearctic populations of Drosophila melanogaster. BMC Evol. Biol. 2019, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Kerchev, I.A. On monogyny of the four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera, Curculionidae: Scolytinae) and its reproductive behavior. Entomol. Rev. 2014, 94, 1059–1066. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Lo, N.; Casiraghi, M.; Salati, E.; Bazzocchi, C.; Bandi, C. How many Wolbachia supergroups exist? Mol. Biol. Evol. 2002, 19, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Che, L.H.; Zhang, S.Q.; Li, Y.; Liang, D.; Pang, H.; Ślipiński, A.; Zhang, P. Genome-wide survey of nuclear protein-coding markers for beetle phylogenetics and their application in resolving both deep and shallow-level divergences. Mol. Ecol. Resour. 2017, 17, 1342–1358. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kononov, A.; Ustyantsev, K.; Blinov, A.; Fet, V.; Baranchikov, Y.N. Genetic diversity of aboriginal and invasive populations of four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera, Curculionidae, Scolytinae). Agric. For. Entomol. 2016, 18, 294–301. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, H.; Hirano, T.; Akizuki, N.; Abe, A.; Nakai, M.; Kunimi, Y.; Inoue, M.N. Multiple infection and reproductive manipulations of Wolbachia in Homona magnanima (Lepidoptera: Tortricidae). Microb. Ecol. 2019, 77, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Dutton, T.J.; Sinkins, S.P. Strain-specific quantification of Wolbachia density in Aedes albopictus and effects of larval rearing conditions. Insect Mol. Biol. 2004, 13, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, P.; Charlat, S.; Labbe, P.; Dehecq, J.S.; Barré, H.; Weill, M. Wolbachia age-sex-specific density in Aedes albopictus: A host evolutionary response to cytoplasmic incompatibility? PLoS ONE 2010, 5, e9700. [Google Scholar] [CrossRef] [Green Version]
- Koehncke, A.; Telschow, A.; Werren, J.H.; Hammerstein, P. Life and death of an influential passenger: Wolbachia and the evolution of CI-modifiers by their hosts. PLoS ONE 2009, 4, e4425. [Google Scholar] [CrossRef] [Green Version]
- Jeyaprakash, A.; Hoy, M.A. Long PCR improves Wolbachia DNA amplification: Wsp sequences found in 76% of sixty-three arthropod species. Insect Mol. Biol. 2000, 9, 393–405. [Google Scholar] [CrossRef]
- Arthofer, W.; Riegler, M.; Schneider, D.; Krammer, M.; Miller, W.J.; Stauffer, C. Hidden Wolbachia diversity in field populations of the European cherry fruit fly, Rhagoletis cerasi (Diptera, Tephritidae). Mol. Ecol. 2009, 18, 3816–3830. [Google Scholar] [CrossRef]
- Hughes, G.L.; Allsopp, P.G.; Brumbley, S.M.; Woolfit, M.; McGraw, E.A.; O’Neill, S.L. Variable infection frequency and high diversity of multiple strains of Wolbachia pipientis in Perkinsiella planthoppers. Appl. Environ. Microbiol. 2011, 77, 2165–2168. [Google Scholar] [CrossRef] [Green Version]
- Schuler, H.; Arthofer, W.; Krumböck, S.; Bertheau, C.; Stauffer, C. Wolbachia infection in the Walnut-husk fly Rhagoletis completa Cresson 1929 (Diptera: Tephritidae). Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2012, 18, 243–245. [Google Scholar]
- Mee, P.T.; Weeks, A.R.; Walker, P.J.; Hoffmann, A.A.; Duchemin, J.B. Detection of low-level Cardinium and Wolbachia infections in Culicoides. Appl. Environ. Microbiol. 2015, 81, 6177–6188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, D.I.; Parker, A.G.; Abd-alla, A.M.; Miller, W.J. High-sensitivity detection of cryptic Wolbachia in the African tsetse fly (Glossina spp.). BMC Microbiol. 2018, 18, 140. [Google Scholar] [CrossRef] [PubMed]
- Uldig, C.K.S.; Wenzel, M.; Fernandez-Begne, P.; Bubnell, J.; Aquadro, C.F. The persistence of low-titer Wolbachia Pipientis Infection in Antibiotic-Treated Drosophila. TAGC 2020. Available online: https://tagc2020.figshare.com/articles/poster/The_persistence_of_low-titer_Wolbachia_pipientis_infection_in_antibiotic-treated_Drosophila/12150591/1 (accessed on 12 August 2020). [CrossRef]
- Yudina, M.A.; Bykov, R.A.; Kotti, B.K.; Vysochina, N.P.; Stakheev, V.V.; Broshkov, A.D.; Zakharov, I.K.; Ilinsky, Y.Y. Wolbachia Infection in Flea Populations (Insecta: Siphonaptera). Biol. Bull. Rev. 2019, 9, 403–411. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality No. | Sampling Sites, Coordinates (Where Available), and Date of Collection | N | Sex | ||
|---|---|---|---|---|---|
| Unknown | ♀ | ♂ | |||
| Sakhalin province (in total) | 60 | 23 | 24 | 13 | |
| 1 | Tomarinsky-1, 48°29′22.2″ N 142°01′49.7″ E, 8 June 2018 | 37 | - | 24 | 13 |
| 2 | Tomarinsky-2, 48°30′26.6″ N 142°00′43.9″ E, 8 June 2018 | 8 | 8 | - | - |
| 3 | Yuzhno-Sakhalinsk, 46°57′50.8″ N 142°45′16.7″ E, 11 June 2018 | 15 | 15 | - | - |
| Krasnoyarsk krai (in total) | 48 | - | 34 | 14 | |
| 4 | Bolshemurtinsky district, 25 December 2019 | 2 | - | 1 | 1 |
| 5 | Emelianovsk forestry, December 2019 | 16 | - | 8 | 8 |
| 6 | Kozulsk forestry, 12–17 February 2020 | 30 | - | 25 | 5 |
| Tomsk province (in total) | 271 | 148 | 63 | 60 | |
| 7 | Tomsky, 56°27′55.0″ N 85°06′46.0″ E, 28 August 2018 | 33 | 33 | - | - |
| 8 | Krivosheinsky, 57°24′27.0″ N 83°55′16.0″ E, 7 August 2018 | 26 | 26 | - | - |
| 9 | Molchanovsky, 57°29′34.0″ N 84°16′27.0″ E, 18 August 2018 | 31 | 31 | - | - |
| 10 | Verkhneketsky, 58°23′30.2″ N 84°06′35.8″ E, 29 August 2019 | 89 | - | 46 | 43 |
| 11 | Chainsky, 57°47′17.0″ N 82°12′34.0″ E, 10 August 2018 | 34 | 34 | - | - |
| 12 | Bakcharsky, 57°16′35.0″ N 81°30′18.0″ E, 15 August 2018 | 58 | 24 | 17 | 17 |
| Udmurtia | |||||
| 13 | Malopurginsky district, 56°38′40.0″ N 53°05′57.6″ E, 11 October 2019 | 38 | 5 | 17 | 16 |
| Total: | 417 | 176 | 138 | 103 | |
| Locality No. | Region, Locality | Conventional PCR | Conventional and Nested PCR | ||
|---|---|---|---|---|---|
| Nuninf/N♀inf/N♂inf | Total %inf (95%CI) | Nuninf/N♀inf/N♂inf | Total %inf (95%CI) | ||
| Sakhalin province (in total) | 8/23/4 | 58.33 (44.88–70.93) | 20/23/9 | 86.67 (75.41–94.06) | |
| 1 | Tomarinsky-1 | 0/23/4 | 72.97 (55.88–86.21) | 0/23/9 | 86.49 (71.23–95.46) |
| 2 | Tomarinsky-2 | 0/0/0 | - | 7/0/0 | 87.5 (47.35–99.68) |
| 3 | Yuzhno-Sakhalinsk | 8/0/0 | 53.33 (26.59–78.73) | 13/0/0 | 86.67 (59.54–98.34) |
| Krasnoyarsk Krai (in total) | 0/25/4 | 60.42 (45.27–74.23) | 0/25/6 | 64.58 (49.46–77.84) | |
| 4 | Bolshemurtinsky district | 0/1/0 | - | 0/1/0 | - |
| 5 | Emelianovsk forestry | 0/8/3 | 68.75 (41.34–88.98) | 0/8/5 | 81.25 (54.35–95.95) |
| 6 | Kozulsk forestry | 0/16/1 | 56.67 (37.43–74.54) | 0/16/1 | 56.67 (37.43–74.54) |
| Tomsk province (in total) | 74/41/15 | 47.97 (41.89–54.1) | 87/45/34 | 61.26 (55.17–67.09) | |
| 7 | Tomsky | 20/0/0 | 60.61 (42.14–77.09) | 22/0/0 | 66.67 (48.17–82.04) |
| 8 | Krivosheinsky | 20/0/0 | 76.92 (56.35–91.03) | 21/0/0 | 80.77 (60.65–93.45) |
| 9 | Molchanovsky | 12/0/0 | 38.71 (21.85–57.81) | 15/0/0 | 48.39 (30.15–66.94) |
| 10 | Verkhneketsky | 1/24/12 | 41.57 (31.21–52.51) | 1/28/24 | 59.55 (48.62–69.83) |
| 11 | Chainsky | 15/0/0 | 44.12 (27.19–62.11) | 19/0/0 | 55.88 (37.89–72.82) |
| 12 | Bakcharsky | 6/17/3 | 44.83 (31.74–58.46) | 9/17/10 | 62.07 (48.37–74.49) |
| Udmurtia | |||||
| 13 | Malopurginsky district | 3/17/1 | 55.26 (38.30–71.38) | 4/17/14 | 92.11 (78.62–98.34) |
| Total: | 85/106/24 | 51.56 (46.65–56.45) | 111/110/63 | 68.11 (63.40–72.56) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bykov, R.; Kerchev, I.; Demenkova, M.; Ryabinin, A.; Ilinsky, Y. Sex-Specific Wolbachia Infection Patterns in Populations of Polygraphus proximus Blandford (Coleoptera; Curculionidae: Scolytinae). Insects 2020, 11, 547. https://doi.org/10.3390/insects11080547
Bykov R, Kerchev I, Demenkova M, Ryabinin A, Ilinsky Y. Sex-Specific Wolbachia Infection Patterns in Populations of Polygraphus proximus Blandford (Coleoptera; Curculionidae: Scolytinae). Insects. 2020; 11(8):547. https://doi.org/10.3390/insects11080547
Chicago/Turabian StyleBykov, Roman, Ivan Kerchev, Marya Demenkova, Artem Ryabinin, and Yury Ilinsky. 2020. "Sex-Specific Wolbachia Infection Patterns in Populations of Polygraphus proximus Blandford (Coleoptera; Curculionidae: Scolytinae)" Insects 11, no. 8: 547. https://doi.org/10.3390/insects11080547