Diversity of Mosquitoes (Diptera: Culicidae) in the Caatinga Biome, Brazil, from the Widespread to the Endemic

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
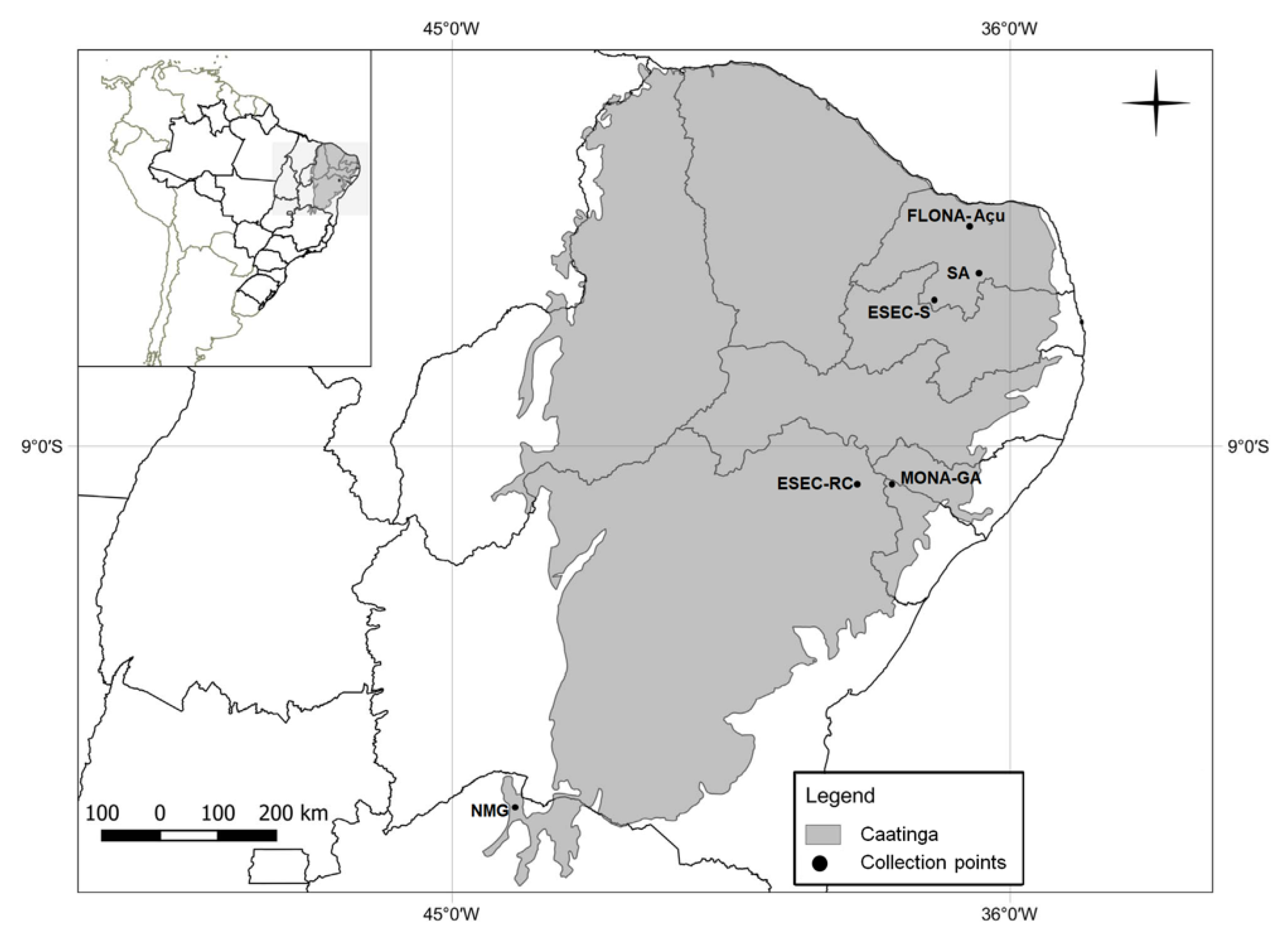
2.1. Study Area and Collection Techniques
2.2. DNA Barcoding
2.3. Data Analyses
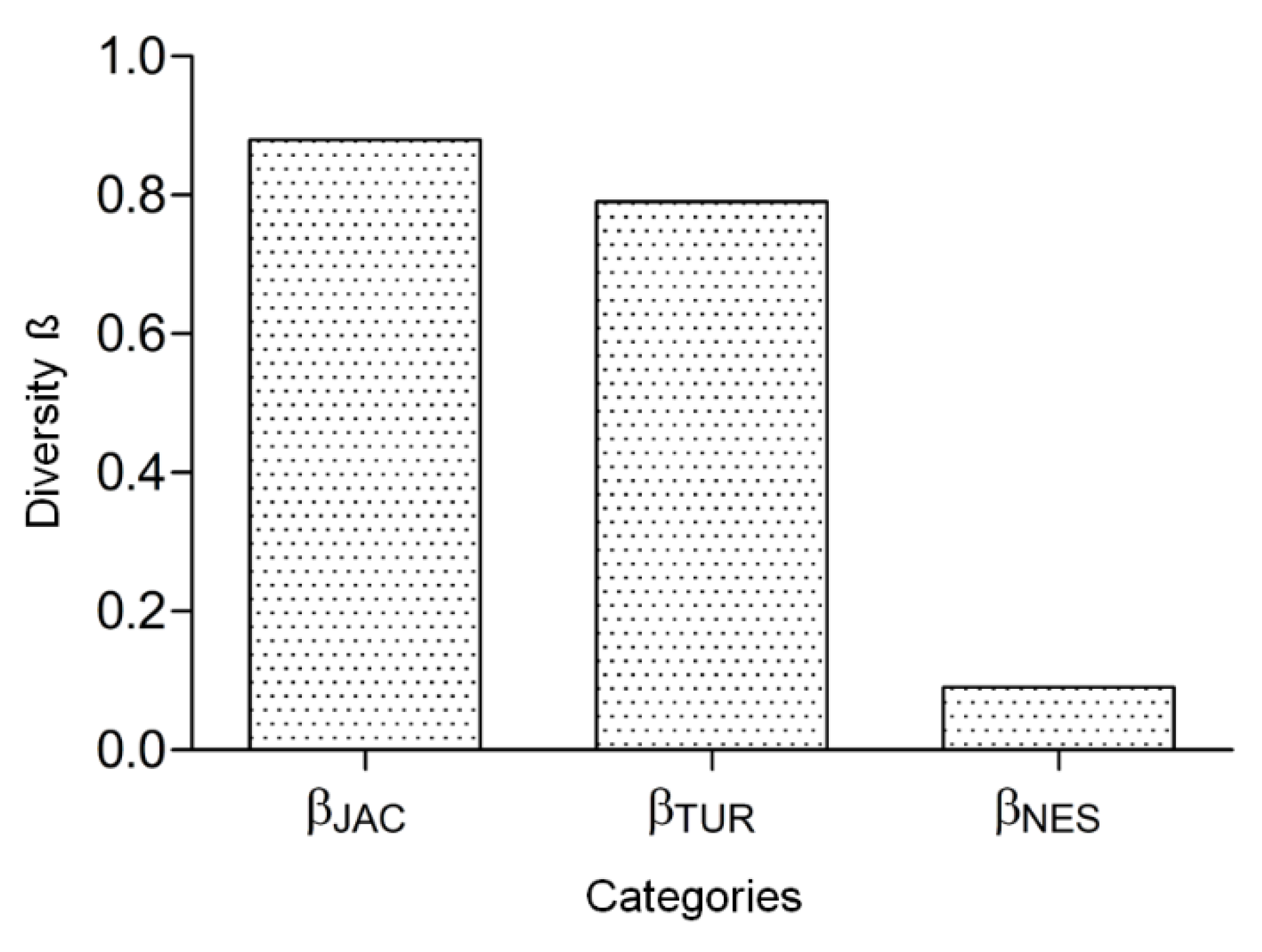
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Leal, I.R.; Tabarelli, M.; Silva, J.M.C. Ecologia e Conservação da Caatinga; Editora da Universidade Federal de Pernambuco: Recife, Brazil, 2003. [Google Scholar]
- Hauff, S.N. Representatividade do Sistema Nacional de Unidades de Conservação na Caatinga; Programa das Nações Unidas Para o Desenvolvimento: Brasília, Brazil, 2010. [Google Scholar]
- Ab’Sáber, A.N. Sertões e sertanejos: Uma geografia humana sofrida. Estudos Avançados 1999, 13, 7–59. [Google Scholar]
- Silva, J.M.C.; Tabarelli, M.; Fonseca, M.T.; Lins, L.V. Biodiversidade da Caatinga: Áreas e Ações Prioritárias Para Conservação; Ministério do Meio Ambiente: Brasília, Brazil, 2004.
- Marengo, J.A.; Alves, L.M.; Beserra, E.A.; Lacerda, F.F. Variabilidade e Mudanças Climáticas No Semiárido Brasileiro. Recursos Hídricos em Regiões Áridas e Semiáridas; Instituto Nacional do Semi-Árido: Campina Grande, Brazil, 2011.
- Vieira, R.M.S.P.; Tomasella, J.; Alvalá, R.C.S.; Sestini, M.F.; Affonso, A.G.; Rodriguez, D.A.; Barbosa, A.A.; Cunha, A.P.M.A.; Valles, G.F.; Crepani, E.; et al. Identifying areas susceptible to desertification in the Brazilian northeast. Solid Earth 2015, 6, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Tomasella, J.; Vieira, R.; Barbosa, A.; Rodriguez, D.; Santana, M.; Sestini, M. Desertification trends in the Northeast of Brazil over the period 2000–2016. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 197–206. [Google Scholar] [CrossRef]
- Giulietti, A.M.; Bocage Neta, A.L.; Castro, A.A.J.F.; Gamarra-Rojas, C.F.L.; Sampaio, E.V.S.B.; Virgínio, J.F.; Harley, R.M. Diagnóstico da Vegetação Nativa do Bioma Caatinga. Biodiversidade da Caatinga: Áreas e Ações Prioritárias Para a Conservação; Ministério do Meio Ambiente: Brasília, Brazil, 2004; pp. 48–90.
- Marteis, L.S.; Natal, D.; Sallum, M.A.M.; Medeiros-Sousa, A.R.; Oliveira, T.M.P.; La Corte, R. Mosquitoes of the Caatinga: 1. Adults stage survey and the emerge of seven news species endemics of a dry tropical forest in Brazil. Acta Trop. 2017, 166, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Marteis, L.S.; Natal, D.; Sallum, M.A.M.; Medeiros-Sousa, A.R.; La Corte, R. Mosquitoes of the Caatinga: 2. Species from periodic sampling of bromeliads and tree holes in a dry Brazilian forest. Acta Trop. 2017, 171, 114–123. [Google Scholar] [CrossRef] [PubMed]
- La Corte, R.; Maia, P.C.R.; Dolabella, S.S.; Cruz, D.E.R.; Marteis, L.S. Mosquitoes of the Caatinga. III. Larval Habitats, Frequency, and Dynamics of Immature and Adult Stages in a Dry Brazilian Forest. J. Med. Entomol. 2019, 56, 120–128. [Google Scholar] [CrossRef]
- Santos, J.C.; Leal, I.R.; Almeida-Cortez, J.S.; Fernandes, G.W.; Tabarelli, M. Caatinga: The scientific negligence experienced by a dry tropical forest. Trop. Conserv. Sci. 2011, 4, 276–286. [Google Scholar] [CrossRef]
- Leal, I.R.; Silva, J.D.; Tabarelli, M.; Lacher, T.E., Jr. Mudando o curso da conservação da biodiversidade na Caatinga do Nordeste do Brasil. Megadiversidade 2005, 1, 139–146. [Google Scholar]
- CNUC/MMA. Cadastro Nacional de Unidades de Conservação. Available online: www.mma.gov.br/cadastro/uc (accessed on 6 March 2017).
- Wardhaugh, C.W. The spatial and temporal distributions of arthropods in forest canopies: Uniting disparate patterns with hypotheses for specialization. Biol. Rev. 2014, 89, 1021–1041. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Oxford, UK, 2004; 256p. [Google Scholar]
- Ward, J.V.; Tockner, K.; Schiemer, F. Biodiversity of floodplain river ecosystems: Ecotones and connectivity. Regul. Rivers Res. Manag. 1999, 15, 125–139. [Google Scholar] [CrossRef]
- Fernandes, G.O. Culicídeos Vetores em uma Unidade de Conservação da Caatinga na Região do Seridó no Rio Grande do Norte—Aspectos da Transmissão de Doenças. Master’s Thesis, Universidade Federal do Rio Grande do Norte, Natal, Brazil, 2011. [Google Scholar]
- Santos, C.F.; Silva, A.C.; Rodrigues, R.A.; Jesus, J.S.R.D.; Borges, M.A.Z. Inventory of mosquitoes (Diptera: Culicidae) in conservation units in Brazilian tropical dry forests. Rev. Inst. Med. Trop. São Paulo 2015, 57, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inácio, C.L.S. Mosquitos (Díptera, Culicidae) em Área de Caatinga Degradada na Região do Seridó, Estado do Rio Grande do Norte, Brasil. Master’s Thesis, Universidade Federal do Rio Grande do Norte, Natal, Brazil, 2016. [Google Scholar]
- Inácio, C.L.S.; Silva, J.H.T.; Freire, R.C.M.; Gama, R.A.; Marcondes, C.B.; Ximenes, M.F.F.M. Checklist of Mosquito Species (Diptera: Culicidae) in the Rio Grande do Norte State, Brazil: Contribution of Entomological Surveillance. J. Med. Entomol. 2017, 54, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Fay, R.W.; Eliason, D.A. A preferred oviposition site as a surveillance method for Aedes aegypti. Mosq News 1966, 26, 531–535. [Google Scholar]
- Maestre-Serrano, R.; Goenaga-Olaya, S. Vigilancia Entomológica para Culicidos a través de larvitrampas en el Departamento del Atlántico (Colombia) 2004–2008. Bol. Malariol. Salud Ambient. 2010, 50, 145–151. [Google Scholar]
- Service, M.W. Mosquito Ecology: Field Sampling Methods, 2nd ed.; Elsevier: London, UK, 1993. [Google Scholar]
- Reinert, J.F. Revised list of abbreviations for genera and subgenera of Culicidae (Diptera) and notes on generic and sub generic changes. J. Am. Mosq. Control Assoc. 2001, 17, 51–55. [Google Scholar]
- Mosquito Taxonomic Inventory. Available online: www.mosquito-taxonomic-inventory.info/ (accessed on 25 January 2020).
- Wilkerson, R.C.; Linton, Y.M.; Fonseca, D.M.; Schultz, T.R.; Price, D.C.; Strickman, D.A. Making mosquito taxonomy useful: A stable classification of tribe Aedini that balances utility with current know led geoevolutionary relationships. PLoS ONE 2015, 10, e0133602. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Porter, C.H.; Collins, F.H. Species-diagnostic differences in a ribosomal DNA internal transcribed spacer from the sibling species Anopheles freeborni and Anopheles hermsi (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 1991, 45, 271–279. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 9 April 2018).
- Baselga, A.; Orme, C.; David, L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Sokal, R.R.; Michener, D. A statistical method for evaluation systematic relationships. Univ. Kans. Sci. Bull. 1958, 28, 1409–1438. [Google Scholar]
- Hammer, O.; Harper, D.T.; Ryan, P. PAST: Paleontological Statistics Software Package for education and data analysis. Paleontol. Electron. 2001, 4, 9. [Google Scholar]
- Marteis, L.S.; Sallum, M.A.M.; Natal, D.; Oliveira, T.M.P.; Gama, R.A.; Dolabella, S.S.; Santos, R.L.C. First Record of Anopheles oryzalimnetes, Anopheles argyritarsis, and Anopheles sawyeri (Diptera: Culicidae) in the Caatinga Biome, Semiarid Scrubland of Sergipe State, Brazil. J. Med. Entomol. 2015, 52, 858–865. [Google Scholar] [CrossRef]
- Guedes, M.L.P. Culicidae (Diptera) no Brasil: Relações entre diversidade, distribuição e enfermidades. Rev. Oecol. Aust. 2012, 16, 283–296. [Google Scholar] [CrossRef]
- Andrade-Lima, D. Present-Day Forest Refuges in Northeastern Brazil. In Biological Diversification in the Tropics; Prance, G.T., Ed.; Columbia University Press: New York, NY, USA, 1982; pp. 245–251. [Google Scholar]
- Chandra, G.; Ghosh, A.; Biswas, D.; Chatterjee, S.N. Host plant preference of Mansonia mosquitoes. J. Aquat. Plant Manag. 2006, 44, 142–144. [Google Scholar]
- Nekola, J.C.; White, P.S. The distance decay of similarity in biogeography and ecology. J. Biogeogr. 1999, 26, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Robich, R.M.; Denlinger, D.L. Diapause in the mosquito Culex pipiens evokes a metabolic switch from blood feeding to sugar gluttony. Proc. Natl. Acad. Sci. USA 2005, 102, 15912–15917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcondes, C.B.; Alencar, J. Revisión de los mosquitos del género Haemagogus Williston (Diptera: Culicidae) de Brasil. Rev. Biomed. 2010, 21, 221–238. [Google Scholar]
- Junk, W.J.; Mello, J.A.S. Impactos ecológicos das represas hidrelétricas na bacia amazônica brasileira. Estudos Avançados 1990, 4, 126–143. [Google Scholar] [CrossRef]
- Civitello, D.J.; Cohen, J.; Fatima, H.; Halstead, N.T.; Liriano, J.; Mc Mahon, T.A.; Rohr, J.R. Biodiversity inhibits parasites: Broad evidence for the dilution effect. Proc. Natl. Acad. Sci. USA 2015, 112, 8667–8671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forattini, O.P.; Kakitani, I.; Massad, E.; Marucci, D. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment: 4-Survey of resting adults and synanthropic behaviour in South-Eastern, Brazil. Rev. Saude Publica 1993, 27, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.C.; Corseuil, E.; Barata, J.M.S. Culicinae (Diptera, Culicidae) ocorrentes no Estado do Rio Grande do Sul, Brasil. Rev. Bras. Entomol. 2005, 49, 275–287. [Google Scholar]
- Hutchings, R.S.G.; Hutchings, R.W.; Menezes, I.S.; Motta, M.A.; Sallum, M.A.M. Mosquitoes (Diptera: Culicidae) From the Northwestern Brazilian Amazon: Araçá River. J. Med. Entomol. 2018, 55, 1188–1209. [Google Scholar] [CrossRef]
- Linton, Y.M.; Pecor, J.E.; Porter, C.H.; Mitchell, L.B.; Garzón-Moreno, A.; Foley, D.H.; Pecor, D.B.; Wilkerson, R.C. Mosquitoes of eastern Amazonian Ecuador: Biodiversity, bionomics and barcodes. Memórias Inst. Oswaldo Cruz 2013, 108, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlin, O.G.W.; Belkin, J.N. Mosquito studies (Diptera, Culicidae) XXXVI. Subgenera Aedinus, Tinolestes and Anoedioporpa of Culex. Contrib. Am. Entomol. Inst. 1980, 17, 1–104. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Techniqueand Habitats | Survey Areas * and Time | |||||
|---|---|---|---|---|---|---|
| MONA–GA 2011–2014 | ESEC–RC 2013–2014 | FLONA–Açu 2011–2013 | ESEC–S 2009–2010 | SA/RN 2014–2015 | NMG 2008–2012 | |
| Human landing catches | x | x | - | x | - | x |
| Shannon trap | x | x | x | x | x | x |
| Tree holes | x | x | - | - | - | - |
| Bromeliads | x | x | - | - | - | - |
| Ponds | x | x | - | - | - | - |
| Larvitrap | - | - | x | - | - | - |
| Ovitrap | x | - | x | x | x | - |
| CDC light trap | - | - | - | - | - | x |
| N | Species | Collections Sites a | CO1 Access Number | |||||
|---|---|---|---|---|---|---|---|---|
| MONAGA /SE | ESEC RC/BA | FLONA /RN | ESEC S/RN | SA RN | NMG | |||
| ANOPHELINAE | ||||||||
| 1 | Anopheles albitarsis s.l. Lynch Arribalzaga, 1878 | x | x | x | x | x | x | |
| b | An. argyritarsis s.l. Robineau-Desvoidy, 1827 | x | x | x | ||||
| 2 | An. argyritarsis s.s. Robineau-Desvoidy, 1827 | x | KT762353 d | |||||
| 3 | An. braziliensis (Chagas, 1907) | x | x | |||||
| 4 | An. darlingi Root, 1926 | x | ||||||
| 5 | An. deaneorum Rosa-Freitas, 1989 | x | x | |||||
| 6 | An. evansae (Brèthes, 1926) | x | ||||||
| 7 | An. marajoara Galvão & Damasceno, 1942 | x | ||||||
| 8 | An. oryzalimnetes Wilkerson & Motoki, 2009 | x c | KT762355 d | |||||
| 9 | An. sawyeri Causey, Deane, Deane & Sampaio, 1943 | x c | KT762356 d | |||||
| 10 | An. albimanus section/Oswaldoi Subgroup | x | ||||||
| 11 | An. triannulatus s.l. (Neiva & Pinto, 1922) | x | x | x | x | |||
| CULICINAE | ||||||||
| Aedomyiini | ||||||||
| 12 | Aedeomyia squamipennis (Lynch Arribalzaga, 1878) | x | x | x | x | |||
| Aedini | ||||||||
| 13 | Aedes fulvithorax (Lutz, 1904) | x c | x | MH118159 MH118160 MH118161 | ||||
| 14 | Ae. fulvus (Wiedemann, 1828) | x | ||||||
| 15 | Ae. hastatus (Dyar, 1922) | x | ||||||
| 16 | Ae. lepidus (Cerqueira & Paraense, 1945) | x | ||||||
| 17 | Ae. scapularis (Rondani,1848) | x | x c | x | x | x | x | MH118146 |
| 18 | Ae. serratus (Theobald, 1901) | x | ||||||
| 19 | Ae. stigmaticus (Edwards, 1922) | x | ||||||
| 20 | Ae. taeniorhynchus (Wiedemann, 1821) | x c | x | x | x | MH118152 | ||
| 21 | Ae. terrens (Walker, 1856) | x | x c | MH118164 MH118165 MH118166 | ||||
| 22 | Ae. aegypti (Linnaeus, 1762) | x | x | x | ||||
| 23 | Ae. albopictus (Skuse, 1895) | x | ||||||
| 24 | Haemagogus janthinomys Dyar, 1921 | x | ||||||
| 25 | Hg. spegazzinii Brethés, 1912 | x | x | x | ||||
| 26 | Hg. sp. near spegazzinii Brethés, 1912 | x c | x | MH118155 | ||||
| 27 | Hg. leucocelaenus (Dyar & Shannon, 1924) | x c | x | MH118162 MH118163 | ||||
| 28 | Psorophora cingulata (Fabricius, 1805) | x | ||||||
| 29 | Ps. discrucians (Walker, 1856) | x | ||||||
| 30 | Ps. ferox (Von Humboldt, 1819) | x | x | x | x | |||
| 31 | Ps. albigenu (Peryassú, 1908) | x | ||||||
| Mansoniini | ||||||||
| 32 | Coquillettidia shannoni (Lane & Antunes, 1937) | x | ||||||
| 33 | Cq. albicosta (Peryassú 1908) | x | ||||||
| 34 | Cq. hermanoi (Lane & Coutinho, 1940) | x | ||||||
| 35 | Cq. juxtamansonia (Chagas, 1907) | x | x | |||||
| 36 | Cq. lynchi (Shannon, 1931) | x | ||||||
| 37 | Cq. nigricans (Coquillett, 1904) | x | x | x | ||||
| 38 | Cq. venezuelensis (Theobald, 1912) | x | x | x | ||||
| 39 | Mansonia humeralis Dyar & Knab, 1916 | x | x | x | ||||
| 40 | Ma. indubitans Dyar & Shannon, 1925 | x c | x c | x | x | x | MH118158 | |
| 41 | Ma. pseudotitillans (Theobald, 1901) | x | x | |||||
| 42 | Ma. titillans (Walker, 1848) | x | x | x | ||||
| 43 | Ma. wilsoni (Barreto & Coutinho, 1944) | x | x | x | ||||
| Culicini | ||||||||
| 44 | Culex conservator Dyar & Knab 1906 | x c | x | MH118148 | ||||
| 45 | Cx. (And.) sp. 1 | x | ||||||
| 46 | Culex nr. (And.) sp. 1 | x c | MH118147 | |||||
| 47 | Cx. ameliae Casal, 1967 | x | ||||||
| 48 | Cx. bidens Dyar, 1922 | x | ||||||
| 49 | Cx. chidesteri Dyar, 1921 | x | x | |||||
| 50 | Cx. Group coronator Dyar & Knab, 1906 | x | x | |||||
| 51 | Cx. declarator Dyar & Knab, 1906 | x | ||||||
| 52 | Cx. habilitator Dyar & Knab, 1906 | x | x | |||||
| 53 | Cx. maxi Dyar, 1928 | x c | x | MH118167 | ||||
| 54 | Cx. nigripalpus Theobald, 1901 | x | x | |||||
| 55 | Cx. restuans Theobald, 1901 | x | ||||||
| 56 | Cx. salinarius Coquillett, 1904 | x | ||||||
| 57 | Cx. saltanensis Dyar, 1928 | x | x | x | ||||
| 58 | Cx. scimitar Branch & Seabrook, 1959 | x | ||||||
| 59 | Cx. imitator Theobald, 1903 | x | ||||||
| 60 | Cx. Gr. Imitator sp. 1 Theobald, 1903 | x c | MH118149 | |||||
| 61 | Cx. Gr. Pleuristriatus sp. 1 Theobald, 1903 | x c | MH118150 MH118151 | |||||
| 62 | Cx. sp. nr. xenophobus | x c | x | MH118153 MH118154 | ||||
| 63 | Cx. aureonotatus Duret & Barreto, 1956 | x | ||||||
| 64 | Cx. bastagarius Dyar & Knab, 1906 | x | ||||||
| 65 | Cx. Complex Vomerifer | x | ||||||
| 66 | Cx. Group Atratus | x | ||||||
| 67 | Cx. ribeirensis/cedecei | x | ||||||
| 68 | Cx. Section melanoconion | x | x | |||||
| Sabethini | ||||||||
| 69 | Limatus durhamii Theobald, 1901 | x | ||||||
| 70 | Li. paraensis (Theobald, 1903) | x | ||||||
| 71 | Runchomyia sp. 1 | x c | MH118156 | |||||
| 72 | Sabethes undosus (Coquillett, 1906) | x | ||||||
| 73 | Wyeomyia (Phoniomyia) sp. 1 | x | x c | MT152293 e | ||||
| 74 | Wy. (Pho.) sp. 2 | x | x c | MT152294 e | ||||
| 75 | Wy. (Pho.) sp. 3 | x c | MT152295 e | |||||
| 76 | Wy. sp. 1 | x c | MH118157 | |||||
| Toxorhynchitini | ||||||||
| 77 | Toxorhynchites (Lynchiella) sp. 1 | x | x c | MF537258 | ||||
| 78 | Tx. (Lyn.) sp. 2 | x | x c | MF537259 | ||||
| Uranotaeniini | ||||||||
| 79 | Uranotaenia geometrica Theobald, 1901 | x | ||||||
| 80 | Ur. lowii Theobald, 1901 | x | x | x | x | |||
| 81 | Ur. pulcherrima Lynch Arribalzaga, 1891 | x | ||||||
| 82 | Ur. apicalis Theobald, 1903 | x | ||||||
| Total | 33 | 26 | 13 | 11 | 13 | 49 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, D.C.; Morais, S.A.; Marteis, L.S.; Gama, R.A.; Freire, R.C.d.M.; Rekowski, B.S.; Ueno, H.M.; La Corte, R. Diversity of Mosquitoes (Diptera: Culicidae) in the Caatinga Biome, Brazil, from the Widespread to the Endemic. Insects 2020, 11, 468. https://doi.org/10.3390/insects11080468
Andrade DC, Morais SA, Marteis LS, Gama RA, Freire RCdM, Rekowski BS, Ueno HM, La Corte R. Diversity of Mosquitoes (Diptera: Culicidae) in the Caatinga Biome, Brazil, from the Widespread to the Endemic. Insects. 2020; 11(8):468. https://doi.org/10.3390/insects11080468
Chicago/Turabian StyleAndrade, David Campos, Sirlei Antunes Morais, Letícia Silva Marteis, Renata Antonaci Gama, Renato Cesar de Melo Freire, Belgrano Santiago Rekowski, Helene Mariko Ueno, and Roseli La Corte. 2020. "Diversity of Mosquitoes (Diptera: Culicidae) in the Caatinga Biome, Brazil, from the Widespread to the Endemic" Insects 11, no. 8: 468. https://doi.org/10.3390/insects11080468