Low Mismatch Rate between Double-Stranded RNA and Target mRNA Does Not Affect RNA Interference Efficiency in Colorado Potato Beetle


1
State Key Laboratory of Biocatalysis of Enzyme Engineering, School of Life Sciences, Hubei University, Wuhan 430062, China
2
Institute of Plant Protection, Xinjiang Academy of Agricultural Sciences, Urumqi 830000, China
3
Institute of Microbial Application, Xinjiang Academy of Agricultural Sciences, Urumqi 830000, China
*
Author to whom correspondence should be addressed.
†
These authors contribute equally to this work.
Insects 2020, 11(7), 449; https://doi.org/10.3390/insects11070449
Submission received: 11 June 2020
/
Revised: 10 July 2020
/
Accepted: 13 July 2020
/
Published: 16 July 2020
(This article belongs to the Special Issue Insecticide Resistance: The Genetic Basis and Underlying Mechanisms in Potato Pests)
{kind=link}
{kind=link}
{kind=link}
Abstract
:RNA interference (RNAi)-based technology has been proven as a novel approach for insect pest control. However, whether insects could evolve resistance to RNAi and the underlying mechanism is largely unknown. The target gene mutations were thought to be one of the potential ways to develop the resistance. Here we predicted the effective siRNA candidates that could be derived from dsRNA against the Colorado potato beetle (CPB) β-Actin gene (dsACT). By site-directed mutagenesis, we synthesized the dsRNAs with the defect in generation of effective siRNAs (and thus were supposed to have comparable low RNAi efficacy). We showed that, with mismatches to the target gene, all the dsRNA variants caused similar levels of silencing of target gene, mortality and larval growth retardation of CPB. Our results suggest that when the mismatch rate of dsACT and target β-Actin mRNA is less than 3%, the RNAi efficiency is not impaired in CPB, which might imply the low possibility of RNAi resistance evolving through the sequence mismatches between dsRNA and the target gene.
1. Introduction
The Colorado potato beetle (CPB, Leptinotarsa decemlineata Say) is one of the most notorious pests in the world and may cause huge yield losses to potato production [1]. CPB has a complicated and diverse life history, a strong fertility, can adapt to adverse conditions, and has the capability to tolerate toxins [2,3], which make CPB control a big challenge [4]. Using chemical insecticides is still the major way to control CPB. However, high selection pressure and capability of tolerance to toxins had resulted in the outbreak of large number of insecticide-resistant CPB populations [5,6]. The insecticide resistance of CPB even promoted the development of the insecticide industry, with hundreds of chemicals against it [5]. Another alternative method is using Bacillus thuringiensis (Bt) toxins, which have potent and insecticidal activity. Pests can be killed by spraying insecticidal Bt toxins, or by expressing it in transgenic crops [7,8]. An increasing number of transgenic Bt crops have been grown globally for many years and have a very effective performance on pest control [9]. Transgenic potato plants expressing Bt Cry3Aa were highly effective in suppressing the population of CPB [10,11]. Yet, evolution of insect resistance to Bt crops may threaten the success of transgenic Bt crops. And pest resistance had emerged in fields where Bt crops were planted [12,13,14].
RNA interference (RNAi) is a post-transcriptional gene silencing mechanism induced by double-stranded RNA (dsRNA) and widely distributed in eukaryotes [15]. RNAi was employed as a useful tool to study gene functions [16], and also repurposed as a novel approach for pest control [17]. Administration of dsRNA targeted against the essential genes of pests could result in the silencing of target gene and influence the growth and fecundity of pests, and even lead to the death of pests. Numerous studies have indicated that transgenic plants expressing dsRNAs targeted against insect genes showed significant reduction in feeding damage [18,19]. By expressing dsRNAs targeted against the β-Actin gene of CPB in potato plastids, full protection was achieved as evidenced by all the CPB larvae dying after feeding on transplastomic potatoes [20]. In 2017, the US Environmental Protection Agency (EPA) approved the first plant-incorporated protectant based on RNAi technology (targeting the western corn rootworm, WCR, Diabrotica virgifera virgifera LeConte) which may approach the market soon [21,22]. It was known that genetic mutation in this population can be a major cause of insect resistance evolution for chemical insecticides and Bt toxins [5,23]. For example, mutations in genes encoding carboxylesterase or acetylcholinesterase could enhance insecticide resistance of insect pests [24]. A point mutation is responsible for the resistance to insecticides acting on ionotropic GABA (G-aminobutyric acid) receptors [25]. Mutations in ABC (ATP-binding cassette) transporters confer resistance to the Bt toxin Cry2Ab in several insects [26,27,28]. Resistance to the Bt toxin Cry1Ac was found to be conferred by mutations in the toxin-binding cadherin proteins [29,30]. Similar to insecticide resistance and Bt resistance, there is no doubt that insects would also evolve resistance to RNAi. Several mechanisms, including mutations of target genes or RNAi core machinery genes, may contribute to RNAi resistance [31]. Researchers established a WCR population resistant to maize expressing DvSnf7 dsRNA and found that dsRNA uptake is changed in dsRNA-resistant WCR [32]. Another study found StaufenC, a dsRNA binding protein, is required for RNAi and is also a potential target for RNAi resistance [33]. These previous studies showed that the mutations of dsRNA uptake and processing could affect the RNAi responses of insects.
In this study, we set out to explore whether sequence mutation of RNAi targeted genes could be one of the possible causes that result in the resistance to RNAi. By selectively mutating the most effective small interfering RNA (siRNA) sites, we synthesized six dsRNA variants to a 200 bp dsACT and compared their insecticidal effects on CPB. We found that there was no significant reduction in RNAi efficacy induced by dsRNA variants compared with control.
2. Materials and Methods
2.1. Insect Material
CPB (L. decemlineata) larvae and adults were collected from a potato field in Urumqi (43.82 °N, 87.61 °E), Xinjiang Uygur Autonomous Region, China. CPBs were collected from the field and reared with wild-type potato plants in an insectary at 28 ± 1 ℃ and 50–60% relative humidity under a 14 h light–10 h dark photoperiod. Eggs from adult CPBs were collected and transferred onto fresh wild-type potato leaves before the bioassays.
2.2. In Vitro dsRNA Synthesis
The T7 RiboMAXTM Express RNAi System (Promega, cat. No. P1700, Madison, WI, USA) was used for in vitro dsRNA synthesis according to the manufacture’s instruction. A cDNA sequence of the ACT gene was acquired from previous research [34]. A 200 bp DNA fragment, covering nucleotides +579 to +778 of the ACT-coding region, was PCR amplified using specific primers (Supplementary Table S1) containing the T7 promoter sequence. The other DNA fragments with mutations were synthesized (Genecreate, Wuhan, Hubei, China) and used as template for dsRNA synthesis with primers containing the T7 promoter sequence (Supplementary Table S1). The reaction mixture contained 2 μL T7 Express Enzyme Mix, 10 μL RiboMAXTM Express T7 2 × Buffer, and 1 μg of DNA template. The mixture was incubated at 37 ℃ for 2–6 h and 70 ℃ for 10 min, then slowly cooled to room temperature (~20 min) to form the dsRNA. The DNA template and single-strand RNA (ssRNA) were removed by DNase (removing DNA template) and RNaseA (removing ssRNA) treatments, respectively. The yield of dsRNA was determined by a Nano Photometer (Implen, Munich, BY, Germany) and the integrity of the full length was detected by gel electrophoresis and stored at −80 ℃ until further use.
2.3. Insect Bioassays
The insect bioassays were performed as described previously [34]. Briefly, the third-instar CPB larvae were starved for 24 h before feeding assays. Equal amounts (16 ng, diluted in 50 μL ddH2O) of dsRNA were painted onto fresh potato leaves covering identical areas of 2 × 2 cm, thus the same dsRNA concentration of 4 ng cm−2 was obtained. ddH2O-painted leaves were used as control. Sufficient dsRNA-coated fresh leaves were supplied to insects and exchanged once per day. Three groups of insects per treatment were investigated, and each group contained 10 larvae. The body weight of larvae and the mortality were recorded daily and the β-Actin expression was detected at day 3.
2.4. Real-Time Quantitative PCR (qRT-PCR)
Total RNA samples were extracted using the RNAiso plus regent (Takara, Kyoto, Japan). Additional genomic DNA was digested by gDNA digester (Yeasen, Shanghai, China); 1.0 μg RNA was used for synthesizing cDNA by using the RevertAid First Strand cDNA Synthesis Kit (Yeasen, Shanghai, China). The RP18 was chosen as the reference gene [35,36]. The 10 μL reaction mixture consisted of 5 μL TB Green Premix Ex Tap Ⅱ (TliRNaseH Plus) (Takara, Kyoto, Japan), 3.5 μL of nuclease-free water, 1 μL of cDNA, and 0.25 μL of forward and reverse primers (Supplementary Table S1). The qPCR protocol included an initial denaturation step at 95 ℃ for 2 min, followed by 40 cycles of 95 ℃ for 5 s, 60 ℃ for 30 s and 72 ℃ for 30 s. The relative expression of β-Actin was calculated by the 2−ΔΔct method [37]. All experiments were repeated at least three times.
2.5. Statistical Analysis of Data
The effective candidates of siRNA were predicted by DNAMAN (v6.0.3.93) and two online tools: http://sidirect2.rnai.jp/ and https://www.invivogen.com/sirnawizard/design.php. The website for sequence alignment is http://multalin.toulouse.inra.fr/multalin/. All parameters are default parameters, such as: (i) A/U (adenine/uracil)at the 5’ end of the antisense strand; (ii) G/C (guanine/cytosine) at the 5’end of the sense strand; (iii) at least five A/U residues in the 5’ terminal one-third of the antisense strand; and (iv) the absence of any GC stretch of more than 9 nt in length [38]. The Kaplan-Meier method was used in the survival curves analysis. The log-rank test was used to evaluate the significance of differences between the two groups. Data of qRT-PCR and body weight were analyzed with one-way ANOVA coupled with Bonferroni’s (equal variances) or Dunnett’s T3 (unequal variances) correction multiple comparison test. A value of p < 0.05 was considered significantly different. Data were statistically analyzed by SPSS version 19.0. Figures were drawn by GraphPad Prism 7 and SnapGene 3.2.1.
3. Results
3.1. Introduction of Point Mutations with Potentially Impaired Generation of Effective siRNAs on RNAi Efficacy in Colorado Potato Beetle
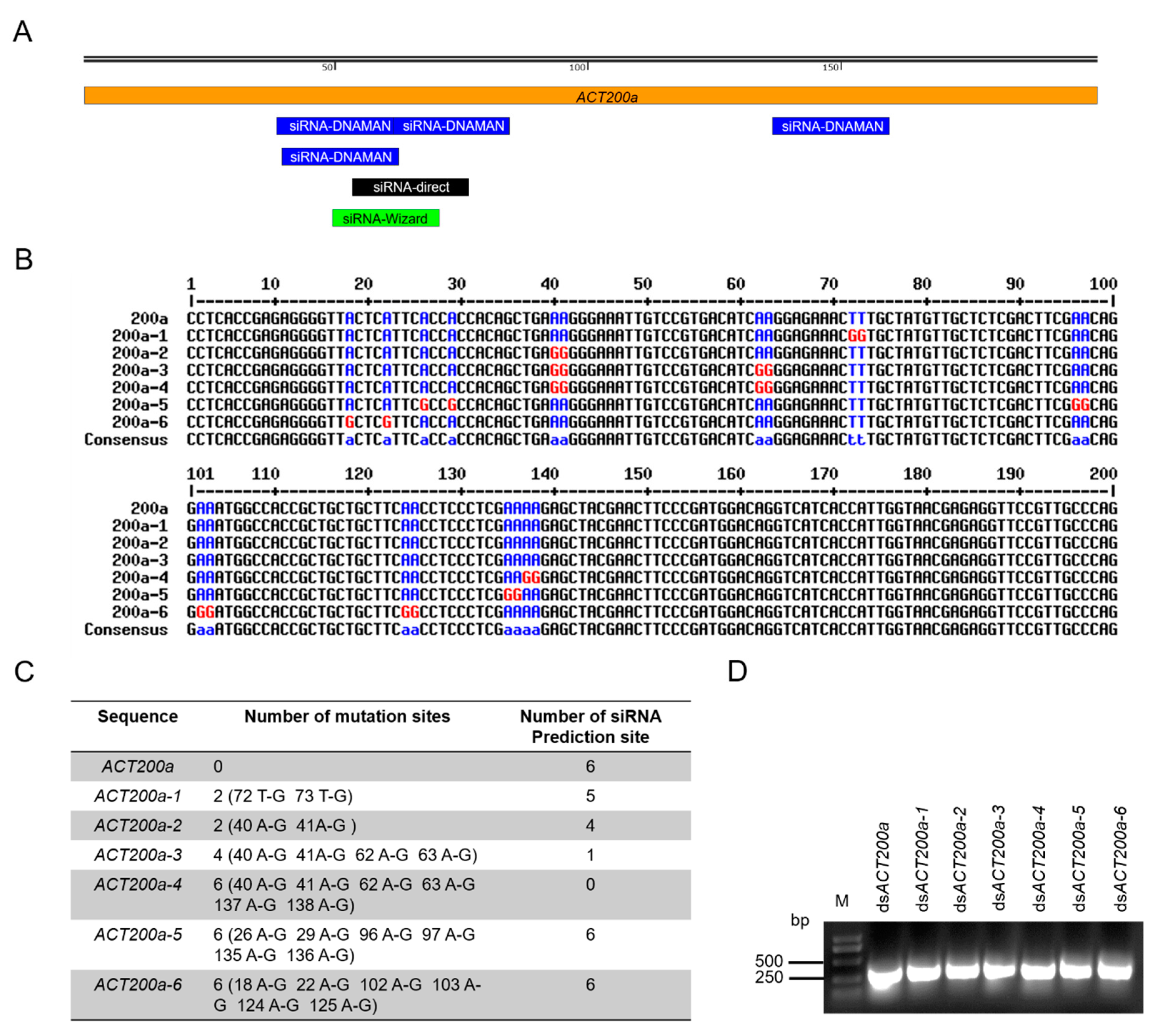
A total of six siRNA sites were predicted to be generated from a 200 bp of ACT fragment (ACT200a) (Figure 1A). Given that the mutation of the target gene sequence would result in the RNAi resistance of insects, there was a high possibility that mutations would occur at the sites which have the potential to derive those effective siRNAs. Therefore, we introduced the point mutations into regions of ACT200a that impaired the effective siRNA generation by T-G or A-G mutation (Figure 1B). For comparison, point mutations outside the effective-siRNA-generation region were introduced as control. Six dsACT200a variants were synthesized. dsACT200a-1 and dsACT200a-2 had mutations to generate five and four effective siRNAs, respectively. Mutations introduced in dsACT200a-3 resulted in the generation of only one effective siRNA, whilst dsACT200a-4 could not generate effective siRNAs. In dsACT200a-5 and dsACT200a-6, mutations were introduced to the sites which did not interfere the generation of effective siRNAs and served as control (Figure 1C). Six dsACT200a variants plus the original dsACT200a were in vitro synthesized and detected by gel electrophoresis. (Figure 1D and Figure S1).
3.2. Effect of dsACT with Different Mutations on RNAi Efficacy in Colorado Potato Beetle Larvae
We next tested whether the mutations of dsACT could affect the RNAi efficacy in CPB. Potato leaves painted with dsACT were fed to third-instar larvae of CPB. Compared with the control (fed with H2O-painted leaves), CPB larvae fed with all the dsACT-treated leaves had increased mortality, inhibited growth and suppressed β-Actin expression (Figure 2A–C). However, no significant differences in mortality, weight gain of survivors and level of gene downregulation were observed among the seven dsACTs treatments (Figure 3). These results revealed that the mutations introduced into dsACT sequences did not markedly affect RNAi efficiency in controlling CPB.
4. Discussion
RNAi technology has been proven as an effective strategy for CPB control [20,39]. Whether and how CPB develops resistance to RNAi remains elusive. In the RNAi pathway, dsRNAs can be processed by Dicer endoribonuclease into 21–23 bp siRNAs, which then are loaded into the RNA-induced silencing complex (RISC) to cleave the target mRNA in a sequence-specific manner [40]. It is critical that the guide strand of siRNA is complementary to the target mRNA, while the mismatch of siRNA to the target mRNA might result in off-target effects and thus impede the RNAi efficiency. Under high pressure of insecticidal dsRNA, it is possible that CPB would accumulate the mutations at the target gene that were targeted by this specific dsRNA to reduce the RNAi efficiency. Sequence mutation is one direction of evolving resistance for CPB to RNAi.
Previously, we showed that transplastomic potato expressing dsRNA against β-Actin genes acquired high resistance to CPB [20]. Recently, naturally occurring mutations of western corn rootworm with resistance to RNAi was first reported in a transgenic corn field screen with subsequent laboratory selection. It showed that DvSnf7-dsRNA resistant WCR was also cross resistant to other dsRNAs, and dsRNA uptake rather than degradation was responsible for evolution of RNAi resistance [32]. This prompted us to investigate whether RNAi resistance could also occur in CPB fed on the dsACT-expressing transplastomic potato plants. Gene mutations are one of the driving forces to evolve the resistance. Potentially, target β-Actin gene mutation could be the resource of RNAi resistance. Under this circumstance, sequence mismatches between dsRNA and target genes could lead to the off-target effects. Since the gene editing tools for introducing mutations to the target gene are not available, we instead introduced the mutations to dsRNAs to mimic the mismatches between the dsRNA and target gene. Our results showed that dsACTs with sequence mutations were still effective and had no significant difference compared to the original dsACT, which demonstrated that the mismatches of dsRNA to target mRNA could not affect the RNAi efficiency in CPB. It was found that the target recognition process is highly sequence specific in the RNAi pathway, but not all positions of siRNA contribute equally to target recognition in a Drosophila melanogaster embryo lysate [41]. The sequence mutation rates of dsACT200 variants were all under 3%, so whether a higher mutation rate (>3%) could affect the RNAi efficiency remains to be investigated. It also should be mentioned that mutations introduced to dsACTs were based on siRNA prediction programs, which could not identify all the effective siRNAs with algorithms. Therefore, the position of a mutation may not be the key nucleotide for the generation of all the effective siRNAs.
5. Conclusions
Taking this together, our study demonstrates that the mismatch rate of dsRNA and a target mRNA lower than 3% in CPB does not reduce the RNAi efficiency in CPB, which may imply that the possibility of RNAi resistance evolving by sequence mutation of target genes would be low. This study will lay the foundation for studying the evolution of RNAi resistance in CPB.
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-4450/11/7/449/s1, Figure S1: The relative location of potential sites were predicted to generate effective siRNAs after mutation introduction to ACT200a (ACT200a-1 to ACT200a-6), Table S1: List of oligonucleotides used in this study.
Author Contributions
J.Z. conceived the project; W.H. and W.X. performed the research; W.H. and J.Z. analyzed the data. K.F. and W.G. support the research and provided the CPB resources. J.Z. wrote the article with contributions of all other authors. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31572071, 31872035).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hare, D.J. Impact of Defoliation by the Colorado Potato Beetle on Potato Yields. J. Econ. Entomol. 1980, 73, 369–373. [Google Scholar] [CrossRef]
- Cingel, A.; Savić, J.; Lazarević, J.; Ćosić, T.; Raspor, M.; Smigocki, A.; Ninković, S. Extraordinary Adaptive Plasticity of Colorado Potato Beetle: “Ten-Striped Spearman” in the Era of Biotechnological Warfare. Int. J. Mol. Sci. 2016, 17, 1538. [Google Scholar] [CrossRef] [PubMed]
- Lyytinen, A.; Lindström, L.; Mappes, J. Genetic Variation in Growth and Development Time Under Two Selection Regimes in Leptinotarsa decemlineata. Entomol. Exp. Appl. 2008, 127, 157–167. [Google Scholar] [CrossRef]
- Alyokhin, A. Colorado Potato Beetle Management on Potatoes: Current Challenges and Future Prospects. In Fruit, Vegetable and Cereal Science and Biotechnology; Tennant, P., Benkeblia, N., Eds.; Global Science Books: Kagawa ken, Japan, 2009; Volume 3, Special Issue 1; pp. 10–19. ISBN 978-4-903313-26-9. [Google Scholar]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado Potato Beetle Resistance to Insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Margus, A.; Piiroinen, S.; Lehmann, P.; Tikka, S.; Karvanen, J.; Lindström, L. Sublethal Pyrethroid Insecticide Exposure Carries Positive Fitness Effects Over Generations in a Pest Insect. Sci. Rep. 2019, 9, 11320. [Google Scholar] [CrossRef] [Green Version]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A Century of Research, Development and Commercial Applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [Green Version]
- Carriere, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing Pyramided Transgenic Bt Crops for Sustainable Pest Management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef]
- Aldemita, R.R.; Reaño, I.M.E.; Solis, R.O.; Hautea, R.A. Trends in Global Approvals of Biotech Crops (1992–2014). GM Crops Food 2015, 6, 150–166. [Google Scholar] [CrossRef]
- Reed, G.L.; Jensen, A.S.; Riebe, J.; Head, G.; Duan, J.J. Transgenic Bt Potato and Conventional Insecticides for Colorado Potato Beetle Management: Comparative Efficacy and Non-Target Impacts. Entomol. Exp. Appl. 2001, 100, 89–100. [Google Scholar] [CrossRef]
- Amiri, A.N.; Bakhsh, A. An Effective Pest Management Approach in Potato to Combat Insect Pests and Herbicide. 3 Biotech 2019, 9, 16. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carriere, Y. Insect Resistance to Bt Crops: Evidence Versus Theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brevault, T.; Carriere, Y. Insect Resistance to Bt Crops: Lessons From the First Billion Acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Carrière, Y.; Brown, Z.S.; Downes, S.J.; Gujar, G.; Epstein, G.; Omoto, C.; Storer, N.P.; Mota-Sanchez, D.; Søgaard Jørgensen, P.; Carroll, S.P. Governing Evolution: A Socioecological Comparison of Resistance Management for Insecticidal Transgenic Bt Crops among Four Countries. Ambio 2020, 49, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maine, E.M. A Conserved Mechanism for Post-Transcriptional Gene Silencing? Genome Biol. 2000, 1, 1018.1–1018.4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannon, G.J. RNA Interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.A.; Heckel, D.G.; Bock, R. Next-Generation Insect-Resistant Plants: RNAi-Mediated Crop Protection. Trends Biotechnol. 2017, 35, 871–882. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of Coleopteran Insect Pests Through RNA Interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a Cotton Bollworm P450 Monooxygenase Gene by Plant-Mediated RNAi Impairs Larval Tolerance of Gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Pest Control. Full Crop Protection From an Insect Pest by Expression of Long Double-Stranded RNAs in Plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef]
- Zotti, M.; Dos Santos, E.A.; Cagliari, D.; Christiaens, O.; Taning, C.N.T.; Smagghe, G. RNA Interference Technology in Crop Protection Against Arthropod Pests, Pathogens and Nematodes. Pest Manag. Sci. 2018, 74, 1239–1250. [Google Scholar] [CrossRef]
- Schiemann, J.; Dietz-Pfeilstetter, A.; Hartung, F.; Kohl, C.; Romeis, J.; Sprink, T. Risk Assessment and Regulation of Plants Modified by Modern Biotechniques: Current Status and Future Challenges. Annu. Rev. Plant Biol. 2019, 70, 699–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Tian, W.; Zhao, J.; Jin, L.; Yang, J.; Liu, C.; Yang, Y.; Wu, S.; Wu, K.; Cui, J.; et al. Diverse Genetic Basis of Field-Evolved Resistance to Bt Cotton in Cotton Bollworm From China. Proc. Natl. Acad. Sci. USA 2012, 109, 10275–10280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatonnet, A. Insecticide Resistance Through Mutations in Cholinesterases or Carboxylesterases: Data Mining in the ESTHER Database. J. Pestic. Sci. 2010, 35, 315–320. [Google Scholar]
- Buckingham, S.D.; Biggin, P.C.; Sattelle, B.M.; Brown, L.A.; Sattelle, D.B. Insect GABA Receptors: Splicing, Editing, and Targeting by Antiparasitics and Insecticides. Mol. Pharmacol. 2005, 68, 942–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Downes, S.; James, W.J.; Lee, S.F.; Reineke, A.; Williams, A.K.; Gordon, K.H. Insect Resistance to Bacillus thuringiensis Toxin Cry2Ab Is Conferred by Mutations in an ABC Transporter Subfamily A Protein. PLoS Genet. 2015, 11, e1005534. [Google Scholar] [CrossRef]
- Mathew, L.G.; Ponnuraj, J.; Mallappa, B.; Chowdary, L.R.; Zhang, J.; Tay, W.T.; Walsh, T.K.; Gordon, K.H.J.; Heckel, D.G.; Downes, S.; et al. ABC Transporter Mis-Splicing Associated With Resistance to Bt Toxin Cry2Ab in Laboratory- And Field-Selected Pink Bollworm. Sci. Rep. 2018, 8, 13531. [Google Scholar] [CrossRef]
- Yang, X.; Chen, W.; Song, X.; Ma, X.; Cotto-Rivera, R.O.; Kain, W.; Chu, H.; Chen, Y.R.; Fei, Z.; Wang, P. Mutation of ABC Transporter ABCA2 Confers Resistance to Bt Toxin Cry2Ab in Trichoplusia ni. Insect Biochem. Mol. Biol. 2019, 112, 103209. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, S.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Non-recessive Bt Toxin Resistance Conferred by an Intracellular Cadherin Mutation in Field-Selected Populations of Cotton Bollworm. PLoS ONE 2012, 7, e53418. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ma, Y.; Wan, P.; Liu, K.; Xiao, Y.; Wang, J.; Cong, S.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Resistance to Bacillus thuringiensis Linked With a Cadherin Transmembrane Mutation Affecting Cellular Trafficking in Pink Bollworm From China. Insect Biochem. Mol. Biol. 2018, 94, 28–35. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [Green Version]
- Khajuria, C.; Ivashuta, S.; Wiggins, E.; Flagel, L.; Moar, W.; Pleau, M.; Miller, K.; Zhang, Y.; Ramaseshadri, P.; Jiang, C.; et al. Development and Characterization of the First dsRNA-resistant Insect Population From Western Corn Rootworm, Diabrotica virgifera virgifera LeConte. PLoS ONE 2018, 13, e0197059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.S.; Mogilicherla, K.; Gurusamy, D.; Chen, X.; Chereddy, S.C.R.R.; Palli, S.R. Double-stranded RNA Binding Protein, Staufen, Is Required for the Initiation of RNAi in Coleopteran Insects. Proc. Natl. Acad. Sci. USA 2018, 115, 8334–8339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Xu, W.; Xu, L.; Fu, K.; Guo, W.; Bock, R.; Zhang, J. Length-dependent Accumulation of Double-Stranded RNAs in Plastids Affects RNA Interference Efficiency in the Colorado Potato Beetle. J. Exp. Bot. 2020, 71, 2670–2677. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: BestKeeper--Excel-based Tool Using Pair-Wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR Normalisation; Strategies and Considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Methods. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ui-Tei, K.; Naito, Y.; Takahashi, F.; Haraguchi, T.; Ohki-Hamazaki, H.; Juni, A.; Ueda, R.; Saigo, K. Guidelines for the Selection of Highly Effective SiRNA Sequences for Mammalian and Chick RNA Interference. Nucleic Acids Res. 2004, 32, 936–948. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-S.; Shukla, J.N.; Gong, Z.J.; Mogilicherla, K.; Palli, S.R. RNA Interference in the Colorado Potato Beetle, Leptinotarsa decemlineata: Identification of Key Contributors. Insect Biochem. Mol. Biol. 2016, 78, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Gordon, K.H.J.; Waterhouse, P.M. RNAi for Insect-Proof Plants. Nat. Biotechnol. 2007, 25, 1231–1232. [Google Scholar] [CrossRef]
- Elbashir, S.M.; Martinez, J.; Patkaniowska, A.; Lendeckel, W.; Tuschl, T. Functional Anatomy of siRNAs for Mediating Efficient RNAi in Drosophila melanogaster Embryo Lysate. EMBO J. 2001, 20, 6877–6888. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Introduction and in vitro synthesis of dsACTs with point mutations. (A) The relative location of effective siRNA candidates at ACT200a which were predicted by indicated bioinformatic toolboxes (DNAMAN, siDirect and siRNA Wizard). (B) Alignment of original ACT200a sequence with mutated ACT200a variants. The high consensus base pairs are represented as blue. The mutated base pairs are indicated as red. (C) Summary of the number and location of mutations in (B). (D) Analysis of the synthesized dsACTs by electrophoresis. M, 1 kb DNA ladder.
Figure 1.
Introduction and in vitro synthesis of dsACTs with point mutations. (A) The relative location of effective siRNA candidates at ACT200a which were predicted by indicated bioinformatic toolboxes (DNAMAN, siDirect and siRNA Wizard). (B) Alignment of original ACT200a sequence with mutated ACT200a variants. The high consensus base pairs are represented as blue. The mutated base pairs are indicated as red. (C) Summary of the number and location of mutations in (B). (D) Analysis of the synthesized dsACTs by electrophoresis. M, 1 kb DNA ladder.

Figure 2.
RNAi effects on Colorado potato beetle (CPB) larvae fed with different in vitro synthesized dsRNA variants. (A) Kaplan-Meier survival curves of third-instar CPB larvae fed with potato leaves that had been painted with identical amounts (4 ng/cm2) of indicated dsACTs. The log-rank test was used to assess the significance of differences between two survival curves. *** p < 0.001; NS, not significant. (B) Mean weight of surviving CPB larvae at the indicated days of feeding. Data are means ± SE (n = 30). (C) Relative expression levels of β-Actin in the CPB larvae in (A) at day 3. Gene expression levels were set as one in CPB larvae fed with H2O-painted control leaves. Data are means ± SE (n = 3). The letters above each bar in (B,C) indicate the significance of differences as determined by one-way ANOVA in SPSS (Bonferroni’s test, p < 0.05).
Figure 2.
RNAi effects on Colorado potato beetle (CPB) larvae fed with different in vitro synthesized dsRNA variants. (A) Kaplan-Meier survival curves of third-instar CPB larvae fed with potato leaves that had been painted with identical amounts (4 ng/cm2) of indicated dsACTs. The log-rank test was used to assess the significance of differences between two survival curves. *** p < 0.001; NS, not significant. (B) Mean weight of surviving CPB larvae at the indicated days of feeding. Data are means ± SE (n = 30). (C) Relative expression levels of β-Actin in the CPB larvae in (A) at day 3. Gene expression levels were set as one in CPB larvae fed with H2O-painted control leaves. Data are means ± SE (n = 3). The letters above each bar in (B,C) indicate the significance of differences as determined by one-way ANOVA in SPSS (Bonferroni’s test, p < 0.05).

Figure 3.
Leaves painted by H2O and dsACT200a and its mutations were fed to third-instar CPB after 3 days.
Figure 3.
Leaves painted by H2O and dsACT200a and its mutations were fed to third-instar CPB after 3 days.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
He, W.; Xu, W.; Fu, K.; Guo, W.; Zhang, J. Low Mismatch Rate between Double-Stranded RNA and Target mRNA Does Not Affect RNA Interference Efficiency in Colorado Potato Beetle. Insects 2020, 11, 449. https://doi.org/10.3390/insects11070449
AMA Style
He W, Xu W, Fu K, Guo W, Zhang J. Low Mismatch Rate between Double-Stranded RNA and Target mRNA Does Not Affect RNA Interference Efficiency in Colorado Potato Beetle. Insects. 2020; 11(7):449. https://doi.org/10.3390/insects11070449
Chicago/Turabian StyleHe, Wanwan, Wenbo Xu, Kaiyun Fu, Wenchao Guo, and Jiang Zhang. 2020. "Low Mismatch Rate between Double-Stranded RNA and Target mRNA Does Not Affect RNA Interference Efficiency in Colorado Potato Beetle" Insects 11, no. 7: 449. https://doi.org/10.3390/insects11070449
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.