Habitat and Seasonality Affect Mosquito Community Composition in the West Region of Cameroon

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
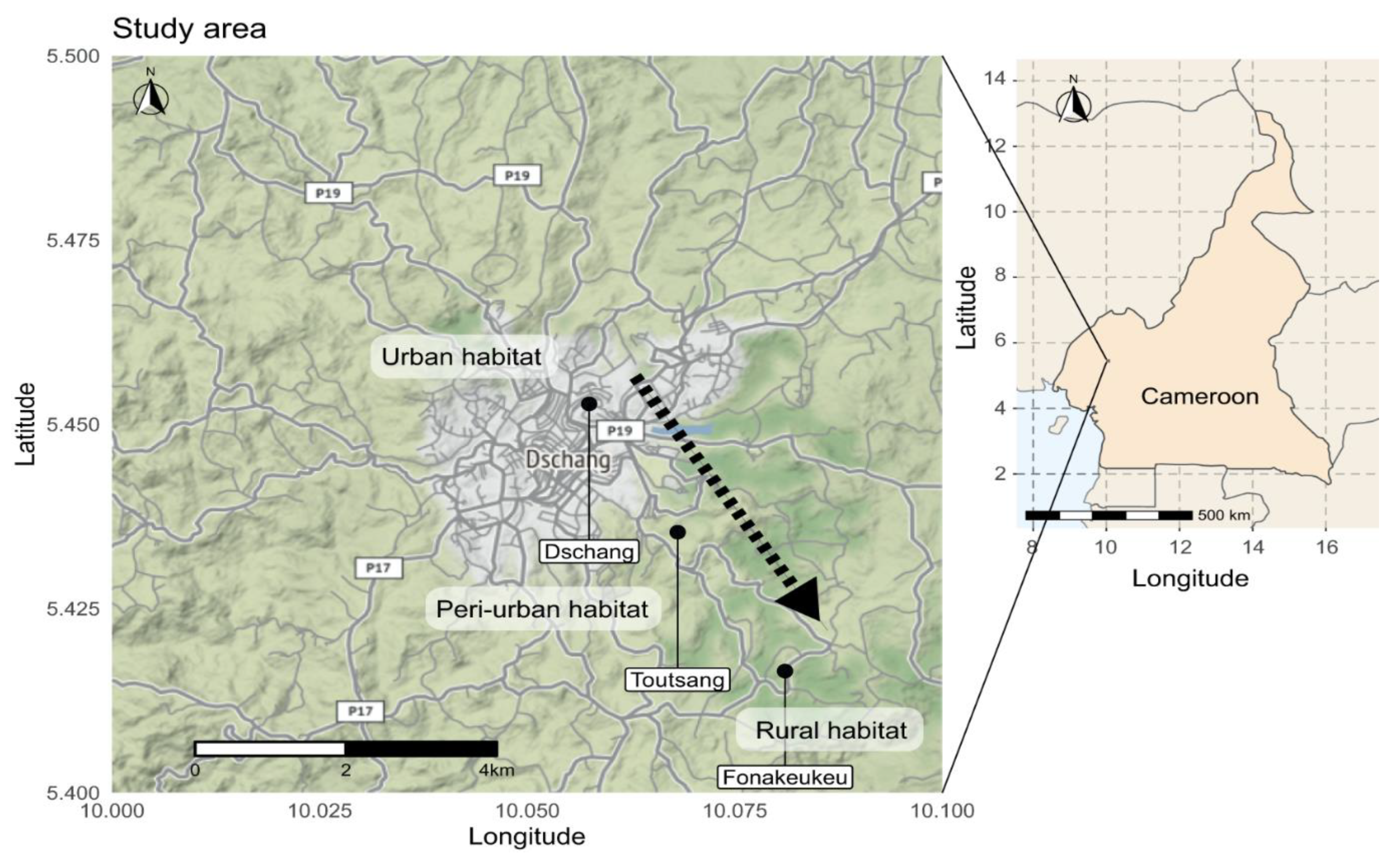
2.1. Description of the Study Sites
2.2. Mosquito Sampling, Identification and Preservation
2.3. Data Analysis
3. Results
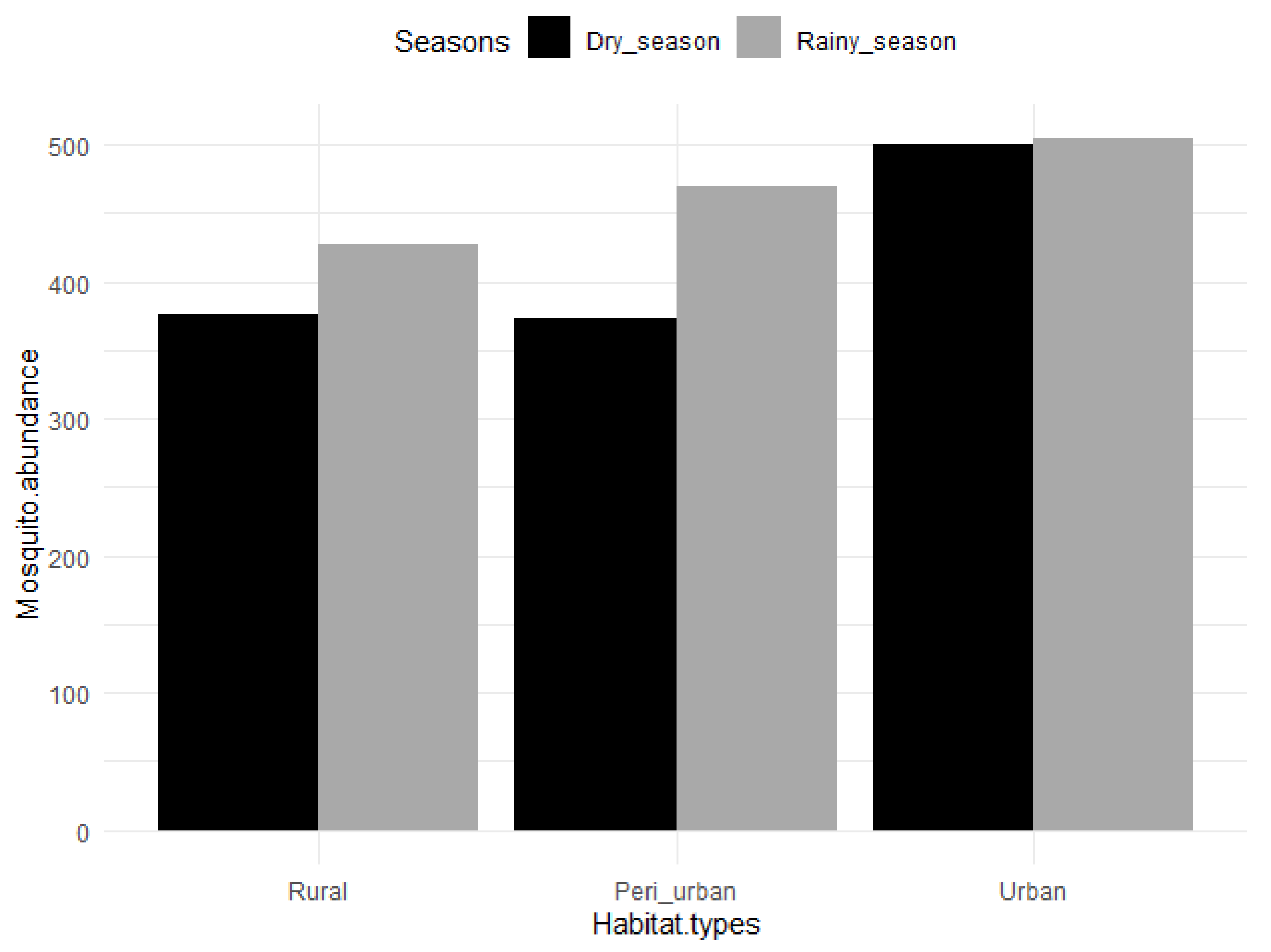
3.1. Mosquito Abundance across Habitats and Seasons
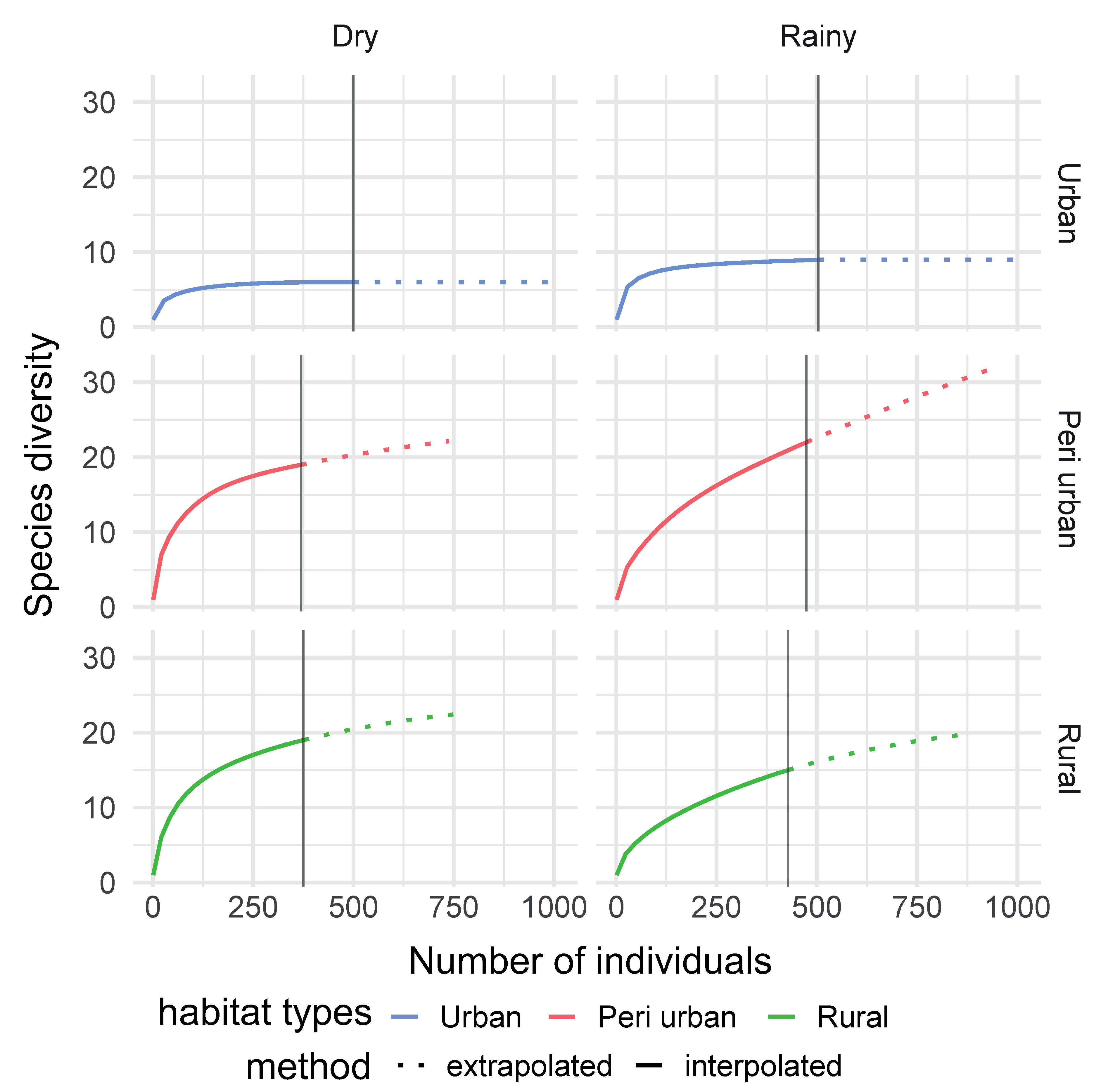
3.2. Mosquito Species Richness across Habitats and Seasons
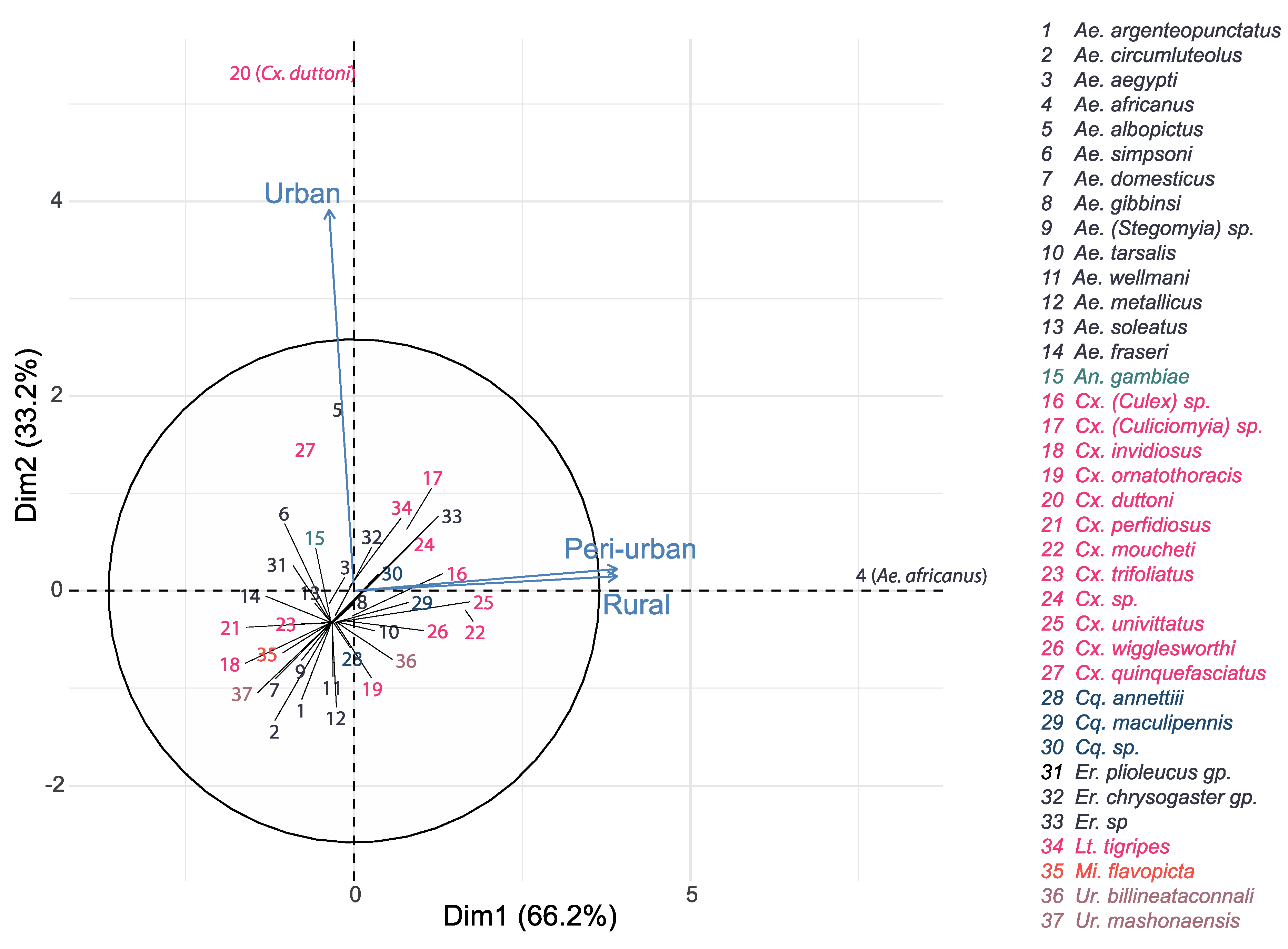
3.2.1. Habitat Type Effect
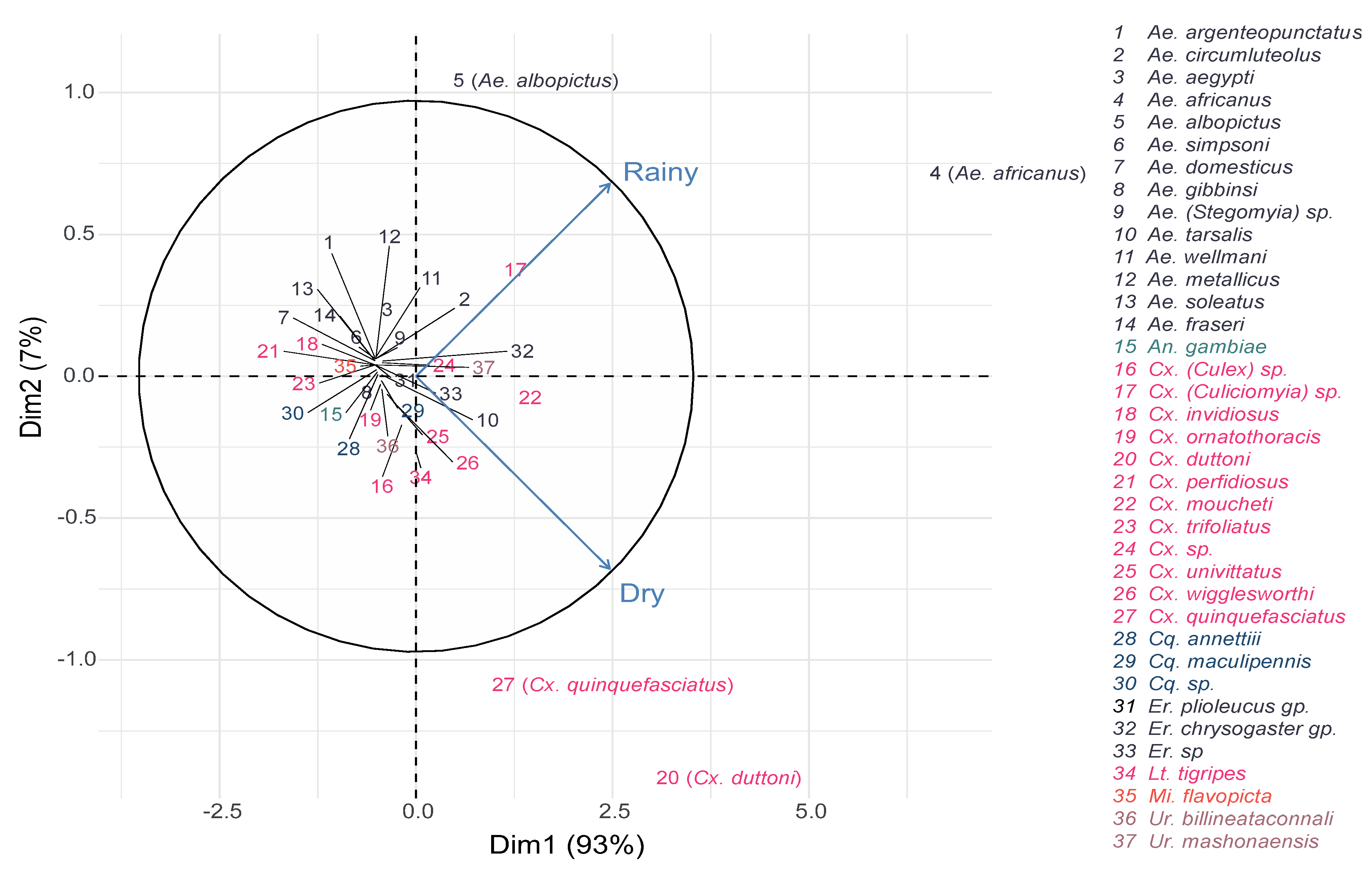
3.2.2. Seasonality Effect
3.2.3. Habitat: Seasonality Effect
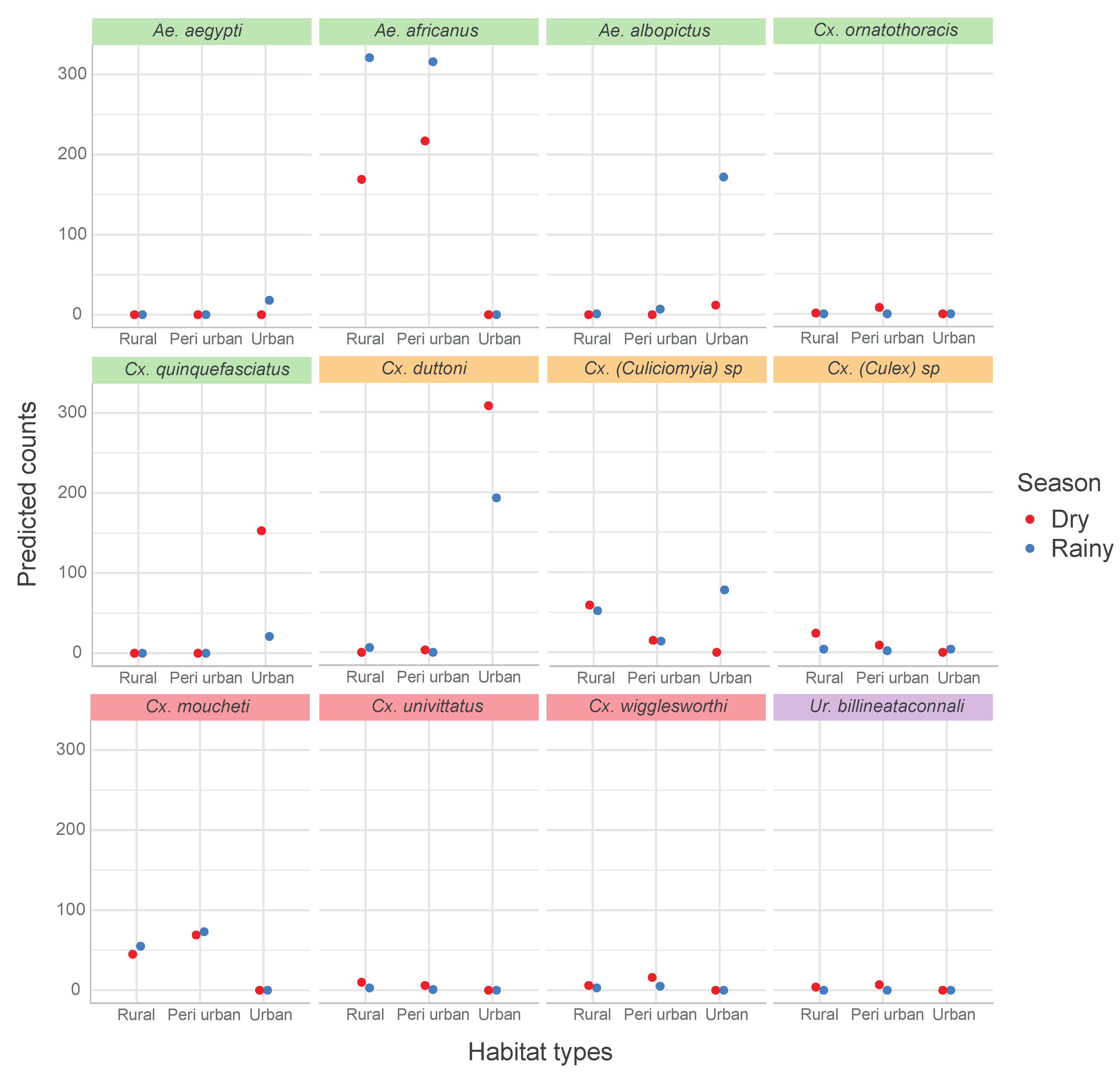
3.3. Potential Medical Importance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C.; Barrett, A.D. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2. [Google Scholar] [CrossRef]
- Chandrasegaran, K.; Lahondère, C.; Escobar, L.E.; Vinauger, C. Linking mosquito ecology, traits, behavior, and disease transmission. Trends Parasitol. 2020, 36, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Franklinos, L.H.; Jones, K.E.; Redding, D.W.; Abubakar, I. The effect of global change on mosquito-borne disease. Lancet Infect. Dis. 2019, 19, e302–e312. [Google Scholar] [CrossRef]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological control of mosquito vectors: Past, present, and future. Insects 2016, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Mwangangi, J.M.; Muturi, E.J.; Muriu, S.M.; Nzovu, J.; Midega, J.T.; Mbogo, C. The role of anopheles arabiensis and anopheles coustani in indoor and outdoor malaria transmission in Taveta district, Kenya. Parasites Vectors 2013, 6, 114. [Google Scholar] [CrossRef] [Green Version]
- Ochieng, C.; Lutomiah, J.; Makio, A.; Koka, H.; Chepkorir, E.; Yalwala, S.; Mutisya, J.; Musila, L.; Khamadi, S.; Richardson, J. Mosquito-borne arbovirus surveillance at selected sites in diverse ecological zones of Kenya, 2007–2012. Virol. J. 2013, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Grillet, M.E. Factors associated with distribution of anopheles aquasalis and anopheles oswaldoi (Diptera: Culicidae) in a malarious area, northeastern Venezuela. J. Med Entomol. 2000, 37, 231–238. [Google Scholar] [CrossRef]
- Preechaporn, W.; Jaroensutasinee, M.; Jaroensutasinee, K. Seasonal prevalence of Aedes aegypti and Ae. albopictus in three topographical areas of southern Thailand. World Acad. Sci. Eng. Technol. 2007, 36, 23–27. [Google Scholar]
- Mordecai, E.A.; Caldwell, J.M.; Grossman, M.K.; Lippi, C.A.; Johnson, L.R.; Neira, M.; Rohr, J.R.; Ryan, S.J.; Savage, V.; Shocket, M.S.; et al. Thermal biology of mosquito-borne disease. Ecol. Lett. 2019, 22, 1690–1708. [Google Scholar] [CrossRef] [Green Version]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.G.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Thongsripong, P.; Green, A.; Kittayapong, P.; Kapan, D.; Wilcox, B.; Bennett, S. Mosquito vector diversity across habitats in central Thailand endemic for dengue and other arthropod-borne diseases. PLoS Negl. Trop. Dis. 2013, 7, e2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, J.H.; Field, H.E.; Luby, S.; Pulliam, J.R.; Daszak, P. Nipah virus: Impact, origins, and causes of emergence. Curr. Infect. Dis. Rep. 2006, 8, 59–65. [Google Scholar] [CrossRef]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komono, B. La fièvrejaune en côte d’ivoire historique, actualité et perspectives de recherche pour la lutte. Médecine D’afrique Noire 2012, 59, 459–469. [Google Scholar]
- Yasuoka, J.; Levins, R. Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Am. J. Trop. Med. Hyg. 2007, 76, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchoumbou, M.A.; Mayi, M.P.A.; Malange, E.N.; Foncha, F.D.; Kowo, C.; Fru-cho, J.; Tchuinkam, T.; Awah-Ndukum, J.; Dorazio, R.; Anong, D.N. Effect of deforestation on prevalence of avian haemosporidian parasites and mosquito abundance in a tropical rainforest of Cameroon. Int. J. Parasitol. 2020, 50, 63–73. [Google Scholar] [CrossRef]
- Bamou, R.; Mbakop, L.R.; Kopya, E.; Ndo, C.; Awono-Ambene, P.; Tchuinkam, T.; Rono, M.K.; Mwangangi, J.; Antonio-Nkondjio, C. Changes in malaria vector bionomics and transmission patterns in the equatorial forest region of Cameroon between 2000 and 2017. Parasites Vectors 2018, 11, 464. [Google Scholar] [CrossRef]
- Johnson, M.T.; Munshi-South, J. Evolution of life in urban environments. Science 2017, 358, eaam8327. [Google Scholar] [CrossRef]
- Jia, P.; Chen, X.; Chen, J.; Lu, L.; Liu, Q.; Tan, X. How does the dengue vector mosquito Aedes albopictus respond to global warming? Parasites Vectors 2017, 10, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadee, D.D.; Martinez, R. Aedes aegypti (L.) in Latin American and Caribbean region: With growing evidence for vector adaptation to climate change? Acta Trop. 2016, 156, 137–143. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti, W., Jr.; Wilke, A.B.B.; Marrelli, M.T. Mosquitoes in urban green spaces: Using an island biogeographic approach to identify drivers of species richness and composition. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafferty, K.D. The ecology of climate change and infectious diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Divis, P.C.; Zakaria, S.K.; Matusop, A.; Julin, R.A.; Conway, D.J.; Cox-Singh, J.; Singh, B. Plasmodium knowlesi: Reservoir hosts and tracking the emergence in humans and macaques. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [Green Version]
- Barrett, A.D.; Monath, T.P. Epidemiology and ecology of yellow fever virus. Adv. Virus Res. 2003, 61, 291–317. [Google Scholar]
- Strode, G.K. Yellow Fever; McGraw-Hill Book Co. Inc.: London, UK, 1951. [Google Scholar]
- Antonio-Nkondjio, C.; Defo-Talom, B.; Tagne-Fotso, R.; Tene-Fossog, B.; Ndo, C.; Lehman, L. High mosquito burden and malaria transmission in a district of the city of Douala, Cameroon. BMC Infect. Dis. 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Antonio-Nkondjio, C.; Simard, F.; Awono-Ambene, P.; Ngassam, P.; Toto, J.; Tchuinkam, T. Malaria vectors and urbanization in the equatorial forest region of south Cameroon. Trans. R Soc. Trop. Med. Hyg. 2005, 99. [Google Scholar] [CrossRef]
- Mayi, M.P.A.; Foncha, D.F.; Kowo, C.; Tchuinkam, T.; Brisco, K.; Anong, D.N.; Ravinder, S.; Cornel, A.J. Impact of deforestation on the abundance, diversity, and richness of culex mosquitoes in a southwest Cameroon tropical rainforest. J. Vector Ecol. 2019, 44, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Cornel, A.J.; Lee, Y.; Almeida, A.P.G.; Johnson, T.; Mouatcho, J.; Venter, M.; De Jager, C.; Braack, L. Mosquito community composition in South Africa and some neighboring countries. Parasites Vectors 2018, 11, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caglioti, C.; Lalle, E.; Castilletti, C.; Carletti, F.; Capobianchi, M.R.; Bordi, L. Chikungunya virus infection: An overview. New Microbiol. 2013, 36, 211–227. [Google Scholar] [PubMed]
- Vest, K.G. Zika virus: A basic overview of an emerging arboviral infection in the western hemisphere. Disaster Med. Public Health Prep. 2016, 10, 707–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braack, L.; de Almeida, A.P.G.; Cornel, A.J.; Swanepoel, R.; de Jager, C. Mosquito-borne arboviruses of African origin: Review of key viruses and vectors. Parasites Vectors 2018, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Kuniholm, M.H.; Wolfe, N.D.; Huang, C.Y.H.; Mpoudi-Ngole, E.; Tamoufe, U.; Burke, D.; Gubler, D. Seroprevalence and distribution of Flaviviridae, Togaviridae, and Bunyaviridae arboviral infections in rural Cameroonian adults. Am. J. Trop. Med. Hyg. 2006, 74, 1078–1083. [Google Scholar] [CrossRef] [Green Version]
- Simo, F.; Bigna, J.; Well, E.; Kenmoe, S.; Sado, F.; Weaver, S.; Moundipa, P.; Demanou, M. Chikungunya virus infection prevalence in Africa: A contemporaneous systematic review and meta-analysis. Public Health 2019, 166, 79–88. [Google Scholar] [CrossRef]
- Peyrefitte, C.; Rousset, D.; Pastorino, B.; Pouillot, R.; Bessaud, M.; Tock, F.; Mansaray, H.; Merle, O.; Pascual, A.; Paupy, C.; et al. Chikungunya virus, Cameroon, 2006. Emerg. Infect. Dis. 2007, 13, 768–771. [Google Scholar] [CrossRef]
- Gake, B.; Vernet, M.A.; Leparc-Goffart, I.; Drexler, J.F.; Gould, E.A.; Gallian, P.; Lamballerie, X.D. Low seroprevalence of Zika virus in Cameroonian blood donors. Braz. J. Infect. Dis. 2017, 21, 481–483. [Google Scholar] [CrossRef]
- Fokam, E.; Levai, L.; Guzman, H.; Amelia, P.; Titanji, V.; Tesh, R.; Weaver, S. Silent circulation of arboviruses in Cameroon. East Afr. Med. J. 2010, 87, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Ndip, L.; Bouyer, D.; da Rosa, T.A.; Titanji, V.; Tesh, R.; Walker, D. Acute spotted fever rickettsiosis among febrile patients, Cameroon. Emerg. Infect. Dis. 2004, 10. [Google Scholar] [CrossRef]
- Tchuandom, S.B.; Tchouangueu, T.F.; Antonio-Nkondjio, C.; Lissom, A.; Djang, J.O.N.; Atabonkeng, E.P.; Kechia, A.; Nchinda, G.; Kuiate, J.-R. Seroprevalence of dengue virus among children presenting with febrile illness in some public health facilities in Cameroon. Pan. Afr. Med. J. 2018, 31, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Keiser, J.; Utzinger, J.; Caldas de Castro, M.; Smith, T.; Tanner, M.; Singer, B. Urbanization in sub-saharan Africa and implication for malaria control. Am. J. Trop. Med. Hyg. 2004, 71, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Hay, S.; Guerra, C.; Tatem, A.; Atkinson, P.; Snow, R. Urbanization, malaria transmission and disease burden in Africa. Nat. Rev. Microbiol. 2005, 3, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Klinkenberg, E.; McCall, P.; Wilson, M.; Amerasinghe, F.; Donnelly, M. Impact of urban agriculture on malaria vectors in Accra, Ghana. Malar. J. 2008, 7, 151. [Google Scholar] [CrossRef] [Green Version]
- Tchuandom, S.B.; Tchadji, J.C.; Tchouangueu, T.F.; Biloa, M.Z.; Atabonkeng, E.P.; Fumba, M.I.M.; Massom, E.S.; Nchinda, G.; Kuiate, J.-R. A cross-sectional study of acute dengue infection in paediatric clinics in Cameroon. BMC Public Health 2019, 19, 958. [Google Scholar] [CrossRef] [Green Version]
- Demanou, M.; Pouillot, R.; Grandadam, M.; Boisier, P.; Kamgang, B.; Hervé, J.P.; Rogier, C.; Rousset, D.; Paupy, C. Evidence of dengue virus transmission and factors associated with the presence of anti-dengue virus antibodies in humans in three major towns in Cameroon. PLoS Negl. Trop. Dis. 2014, 8, e2950. [Google Scholar] [CrossRef]
- Edwards, F.W. Mosquitoes of the Ethiopian Region. III.-Culicine Adults and Pupa; The Trustees of the British Museum: London, UK, 1941. [Google Scholar]
- Jupp, P.G. Mosquitoes of Southern Africa: Culicinae and Toxorhynchitinae; Ekogilde Publishers: Hartebeespoort, South Africa, 1996. [Google Scholar]
- Wilkerson, R.C.; Linton, Y.-M.; Fonseca, D.M.; Schultz, T.R.; Price, D.C.; Strickman, D.A. Making mosquito taxonomy useful: A stable classification of tribe Aedini that balances utility with current knowledge of evolutionary relationships. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Chao, A.; Colwell, R.K.; Lin, C.-W.; Gotelli, N.J. Sufficient sampling for asymptotic minimum species richness estimators. Ecology 2009, 90, 1125–1133. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. Multivariate Linear Models in R; R Companion Applied Regression: Thousand Oaks, LA, USA, 2011. [Google Scholar]
- Kahle, D.; Wickham, H. Ggmap: Spatial visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar] [CrossRef] [Green Version]
- Salaün, J.J.; Rickenbach, A.; Brès, P.; Brottes, H.; Germain, M.; Eouzan, J.P.; Ferrara, L. Arboviruses isolated from mosquitoes in Cameroon. Bull. World Health Organ. 1969, 41, 233–241. [Google Scholar] [PubMed]
- Karabatsos, N. Supplement to international catalogue of arboviruses including certain other viruses of vertebrates. Am. J. Trop. Med. Hyg. 1978, 27, 372. [Google Scholar]
- Institut Pasteur de Dakar. Pertinence des Critères Oms 2000 de Paludisme Grave Chez L’enfant Non Immun à Dakar, Sénégal; Organization Mondiale de la Sante: Dakar, Senegal, 2000. [Google Scholar]
- Brottes, H.; Rickenbach, A.; Brès, P.; Salaün, J.J.; Ferrara, L. Arboviruses in the Cameroon. Isolation from mosquitoes. Bull. World Health Organ. 1966, 35, 811–825. [Google Scholar] [PubMed]
- Hubálek, Z.; Halouzka, J. West Nile fever-a reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; De Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rezende, R.H.; Romano, M.C.; Claro, M.I.; Caleiro, S.G.; Cerdeira-Sabino, E.; Felix, A.C.; Bissoli, J.; Hill, S.; Rodrigues Faria, N.; Cardoso da Silva, T.C.; et al. First report of Aedes albopictus infected by Dengue and Zika virus in a rural outbreak in Brazil. PLoS ONE 2020, 15, e0229847. [Google Scholar] [CrossRef]
- Hasty, J.M.; Felix, G.E.; Amador, M.; Barrera, R.; Santiago, G.S.; Nakasone, L.; Park, S.Y.; Okoji, S.; Honda, E.; Asuncion, B. Entomological investigation detects dengue virus type 1 in aedes (Stegomyia) albopictus (Skuse) during the 2015–2016 outbreak in Hawaii. Am. J. Trop. Med. Hyg. 2020, 102, 869–875. [Google Scholar] [CrossRef]
- Kamgang, B.; Vazeille, M.; Tedjou, A.N.; Wilson-Bahun, T.A.; Yougang, A.P.; Mousson, L.; Wondji, C.S.; Failloux, A.-B. Risk of dengue in central Africa: Vector competence studies with Aedes aegypti and Aedes albopictus (Diptera: Culicidae) populations and dengue 2 virus. PLoS Negl. Trop. Dis. 2019, 13, e0007985. [Google Scholar] [CrossRef] [Green Version]
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Mboui Ondo, S.; Jiolle, D. Zika virus in Gabon (Central Africa) 2007: A new threat from Aedes albopictus? PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Meegan, J.M.; Bailey, C.L. Rift valley fever. Arboviruses Epidemiol. Ecol. 1988, 4, 51–76. [Google Scholar]
- Smithburn, K.; Haddow, A.; Gillett, J. Rift Valley fever. Isolation of the virus from wild mosquitoes. Br. J. Exp. Pathol. 1948, 29, 107. [Google Scholar] [PubMed]
- Williams, M.C.; Woodall, J.P.; Corbet, P.S.; Gillett, J.D. O’nyong-nyong fever: An epidemic virus disease in East Africa. 8. Virus isolations from anopheles mosquitoes. Trans. R Soc. Trop. Med. Hyg. 1965, 59. [Google Scholar] [CrossRef]
- Corbet, P.S.; Williams, M.; Gillett, J. O’nyong-nyong Fever: An epidemic virus disease in east Africa. Trans. R. Soc. Trop. Med. Hyg. 1961, 55, 463–480. [Google Scholar] [CrossRef]
- Diallo, M.A.; Diongue, K.; Ndiaye, M.; Gaye, A.; Deme, A.; Badiane, A.S. Evaluation of CareStart™ Malaria HRP2/pLDH (Pf/pan) combo test in a malaria low transmission region of Senegal. Malar. J. 2017, 16. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, J.; Chirico, J.; Boqvist, S.; Thu, H.T.V.; Magnusson, U. Occurrence of Japanese encephalitis virus mosquito vectors in relation to urban pig holdings. Am. J. Trop. Med. Hyg. 2012, 87, 1076–1082. [Google Scholar] [CrossRef] [Green Version]
- Ferraguti, M.; Martínez-de La Puente, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effects of landscape anthropization on mosquito community composition and abundance. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zahouli, J.B.; Utzinger, J.; Adja, M.A.; Müller, P.; Malone, D.; Tano, Y.; Koudou, B.G. Oviposition ecology and species composition of Aedes spp. and Aedes aegypti dynamics in variously urbanized settings in arbovirus foci in southeastern Côte d’Ivoire. Parasites Vectors 2016, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Steiger, D.B.M.; Ritchie, S.A.; Laurance, S.G. Mosquito communities and disease risk influenced by land use change and seasonality in the Australian tropics. Parasites Vectors 2016, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- Karim, M.R.; Islam, M.M.; Farid, M.S.; Rashid, M.A.; Akter, T.; Khan, H.R. Spatial distribution and seasonal fluctuation of mosquitoes in Dhaka City. Int. J. Fauna Biol. Stud. 2013, 1, 42–46. [Google Scholar]
- Santos, C.F.; Silva, A.C.; Rodrigues, R.A.; Jesus, J.S.R.D.; Borges, M.A.Z. Inventory of mosquitoes (Diptera: Culicidae) in conservation units in Brazilian tropical dry forests. Rev. Do Inst. De Med. Trop. De Sao Paulo 2015, 57, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Demanou, M.; Antonio-Nkondjio, C.; Ngapana, E.; Rousset, D.; Paupy, C.; Manuguerra, J.-C.; Zeller, H. Chikungunya outbreak in a rural area of Western Cameroon in 2006: A retrospective serological and entomological survey. BMC Res. Notes 2010, 3, 128. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mosquito Species | Rural | Peri-Urban | Urban | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Rainy | Dry | Total | Rainy | Dry | Total | Rainy | Dry | Total | Grand-Total | |
| Ae. argenteopunctatus | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 2 |
| Ae. circumluteolus | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 2 |
| Ae. aegypti | 0 | 0 | 0 | 0 | 0 | 0 | 18 | 0 | 18 | 18 |
| Ae. africanus | 316 | 217 | 533 | 321 | 169 | 490 | 0 | 0 | 0 | 1023 |
| Ae. albopictus | 7 | 0 | 7 | 1 | 0 | 1 | 172 | 12 | 184 | 192 |
| Ae. simpsoni | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 2 |
| Ae. domesticus | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 |
| Ae. gibbinsi | 0 | 0 | 0 | 0 | 5 | 5 | 0 | 0 | 0 | 5 |
| Ae. (Stegomyia) spp. | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 2 |
| Ae. tarsalis | 0 | 3 | 3 | 4 | 5 | 9 | 0 | 0 | 0 | 0 |
| Ae. wellmani | 2 | 0 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 3 |
| Ae. metallicus | 2 | 0 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 3 |
| Ae. soleatus | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 |
| Ae. fraseri | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 |
| An. gambiae s.l. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 4 | 4 |
| Cx. (Culex) spp. | 2 | 9 | 11 | 4 | 24 | 28 | 4 | 0 | 4 | 43 |
| Cx. (Culiciomyia) spp. | 14 | 15 | 29 | 52 | 59 | 111 | 78 | 0 | 78 | 218 |
| Cx. invidiosus | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cx. ornatothoracis | 0 | 8 | 8 | 0 | 1 | 1 | 0 | 0 | 0 | 9 |
| Cx. duttoni | 0 | 3 | 3 | 6 | 0 | 6 | 193 | 308 | 501 | 510 |
| Cx. perfidiosus | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Cx. moucheti | 73 | 69 | 142 | 55 | 45 | 100 | 0 | 0 | 0 | 242 |
| Cx. trifoliatus | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| Cx. spp. | 0 | 1 | 1 | 8 | 2 | 10 | 0 | 3 | 3 | 14 |
| Cx. univittatus | 1 | 6 | 7 | 3 | 10 | 13 | 0 | 0 | 0 | 20 |
| Cx. wigglesworthi | 5 | 16 | 21 | 3 | 6 | 9 | 0 | 0 | 0 | 30 |
| Cx. pipiens s.l. | 0 | 0 | 0 | 0 | 0 | 0 | 21 | 153 | 174 | 174 |
| Cq. annettii | 0 | 2 | 2 | 0 | 3 | 3 | 0 | 0 | 0 | 5 |
| Cq. maculipennis | 1 | 4 | 5 | 1 | 5 | 6 | 0 | 0 | 0 | 11 |
| Cq. spp. | 0 | 0 | 0 | 0 | 3 | 3 | 0 | 0 | 0 | 3 |
| Er. “plioleucus” gp. | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| Er. “chrysogaster” gp. | 1 | 2 | 3 | 2 | 4 | 6 | 6 | 0 | 6 | 15 |
| Er. spp. | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| Lt. tigripes | 1 | 9 | 10 | 5 | 21 | 26 | 11 | 20 | 31 | 67 |
| Mi. flavopicta | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Ur. billineata connali | 0 | 7 | 7 | 0 | 4 | 4 | 0 | 0 | 0 | 11 |
| Ur. mashonaensis | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Total | 427 | 376 | 803 | 470 | 373 | 843 | 504 | 500 | 1004 | 2650 |
| Rural Area | Peri-urban Area | Urban Area | p | |
|---|---|---|---|---|
| Observed species richness (S) | 25 | 30 | 11 | <0.05 |
| Chao 1 | 32 | 121 | 11 | <0.05 |
| ACE | 33.37 | 51.97 | 11.33 | <0.05 |
| Rainy Season | Dry Season | p | |
|---|---|---|---|
| Observed species richness (S) | 24 | 28 | <0.05 |
| Chao 1 | 24.6 | 64 | <0.05 |
| ACE | 26.04 | 38.33 | <0.05 |
| Mosquito Species | Arboviruses | References |
|---|---|---|
| Ae. argenteopunctatus | NKOV, SFV, BUNV, MIDV, PGAV, SHOV, WESV, CHIKV, NRIV | [35,56,57,58] |
| Ae. circumluteolus | SPOV, WESV, NDUV, RVFV, BUNV, SIMV, MIDV, GERV, LEBV, PGAV, SHOV, SPOV, WNV | [35,56,57,59,60,61,62] |
| Ae. aegypti | YFV, DENV, ZIKV, CHIKV, RVFV, BUNV, DUGV, ORUV, USUV, VEEV, WNV, SFV, WESV, BBKV | [35,57,60,62] |
| Ae. africanus | YFV, ZIKV, BOUV, BBKV, CHIKV, WESV, WNV, ORUV, BOZV | [35,56,63] |
| Ae. albopictus | DENV, CHIKV, ZIKV, JEV, USUV, YFV | [35,61,62,63,64,65] |
| Ae. simpsoni | YFV, BBKV, NRIV, WESV | [35,56,61,62] |
| Ae. “domesticus” group | WESV, BUNV | [57,62] |
| Ae. “tarsalis” group | MIDV, WESV, KEDV, PGAV, PATV, ZIKV, SHOV, RVFV | [35,56,57,59,62,63,66,67] |
| Ae. metallicus | YFV, ZIKV, WESV | [35,62] |
| An. gambiae s.l. | BUNV, BWAV, CHIKV, ILEV, MIDV, ONNV, ORUV, ZIKV, TATV, NRIV, NDOV, BGIV | [57,68,69] |
| Cx. (Culiciomyia) spp. | NTAV, YAOV, CHIKV, MIDV, BBKV, BGIV | [35,59,62,63] |
| Cx. duttoni | Arb11266 | [63] |
| Cx. moucheti | NTAV | [59] |
| Cx. univittatus | WNV, USUV, WESV, SINV, RVFV, SPOV | [35]. |
| Cx. pipiens s.l. | JEV, WNV, RVFV, USUV, CHIKV, EEEV, KUNV, MTBV, MURV, OROV, RRV, SLEV, SINV, VEEV, WANV, WEEV, WNV, BBKV | [35,57,60,61,62,70,71] |
| Cq. maculipennis | CHIKV | [62] |
| Er. “chrysogaster” group | SFV, MIDV, NTAV, SIMV, NKOV, RVFV | [56,57,59,66] |
| Lt. tigripes | NTAV, KAMV, MOSV, SINV, BBKV | [35,57,59,62,63] |
| Ur. mashonaensis | WESV | [62] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayi, M.P.A.; Bamou, R.; Djiappi-Tchamen, B.; Fontaine, A.; Jeffries, C.L.; Walker, T.; Antonio-Nkondjio, C.; Cornel, A.J.; Tchuinkam, T. Habitat and Seasonality Affect Mosquito Community Composition in the West Region of Cameroon. Insects 2020, 11, 312. https://doi.org/10.3390/insects11050312
Mayi MPA, Bamou R, Djiappi-Tchamen B, Fontaine A, Jeffries CL, Walker T, Antonio-Nkondjio C, Cornel AJ, Tchuinkam T. Habitat and Seasonality Affect Mosquito Community Composition in the West Region of Cameroon. Insects. 2020; 11(5):312. https://doi.org/10.3390/insects11050312
Chicago/Turabian StyleMayi, Marie Paul Audrey, Roland Bamou, Borel Djiappi-Tchamen, Albin Fontaine, Claire L. Jeffries, Thomas Walker, Christophe Antonio-Nkondjio, Anthony John Cornel, and Timoléon Tchuinkam. 2020. "Habitat and Seasonality Affect Mosquito Community Composition in the West Region of Cameroon" Insects 11, no. 5: 312. https://doi.org/10.3390/insects11050312