The Roles of DNA Methyltransferases 1 (DNMT1) in Regulating Sexual Dimorphism in the Cotton Mealybug, Phenacoccus solenopsis

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Gene Identification and Bioinformatics Analysis
2.3. RNA Isolation and the Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
2.4. Rapid Amplification of cDNA Ends (RACE)
2.5. Gene Phylogeny
2.6. RNA Inference
2.7. Statistical Analysis
3. Results
3.1. The Cotton Mealybug Genome Encodes Two Copies of DNMT1 but Not DNMT3
3.2. The Cotton Mealybug PsDNMT1 Proteins Possess Conserved Domain Structures
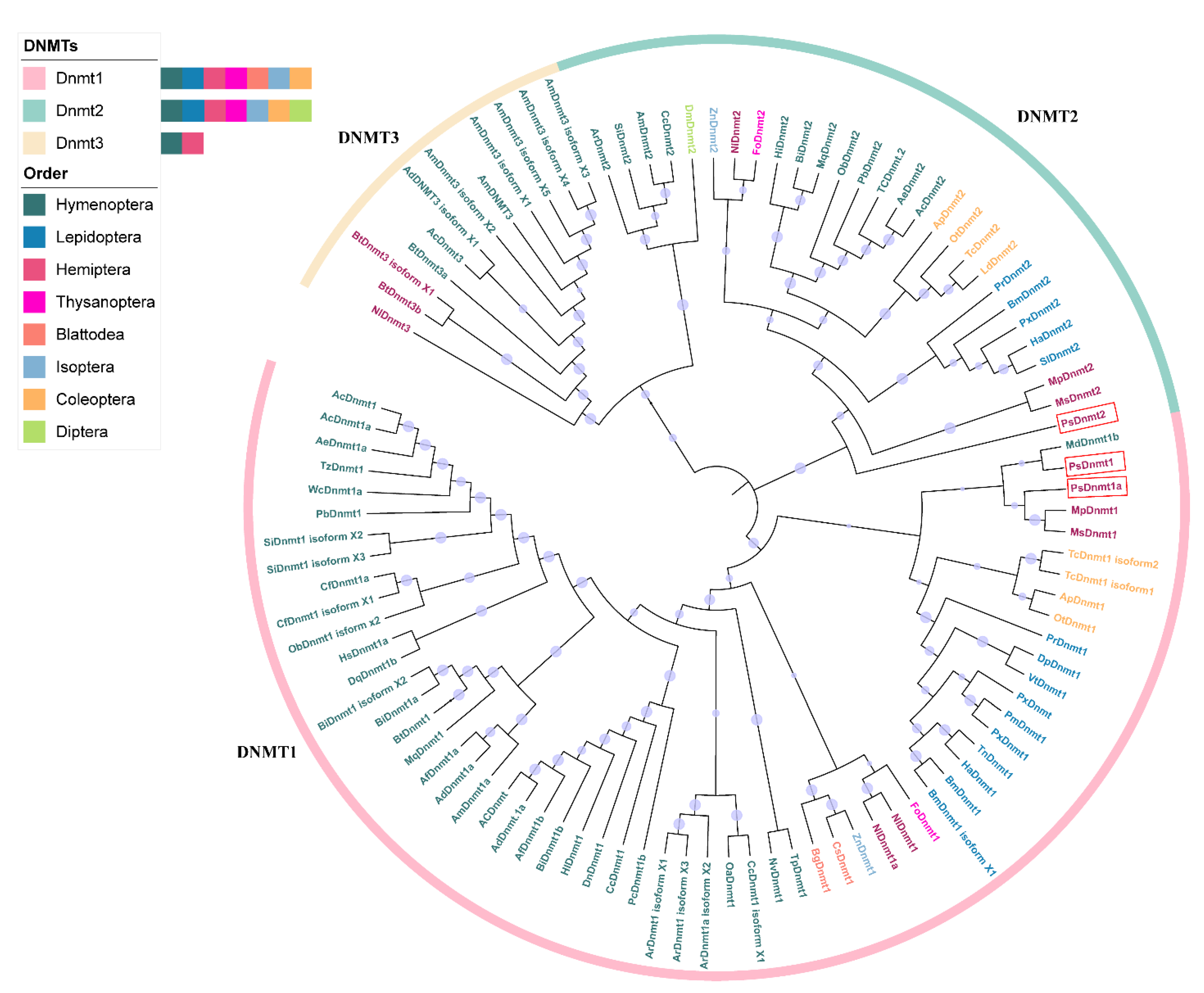
3.3. PsDNMT1A and PsDNMT1B Cluster into Two Different Phylogeny Sub-Clades
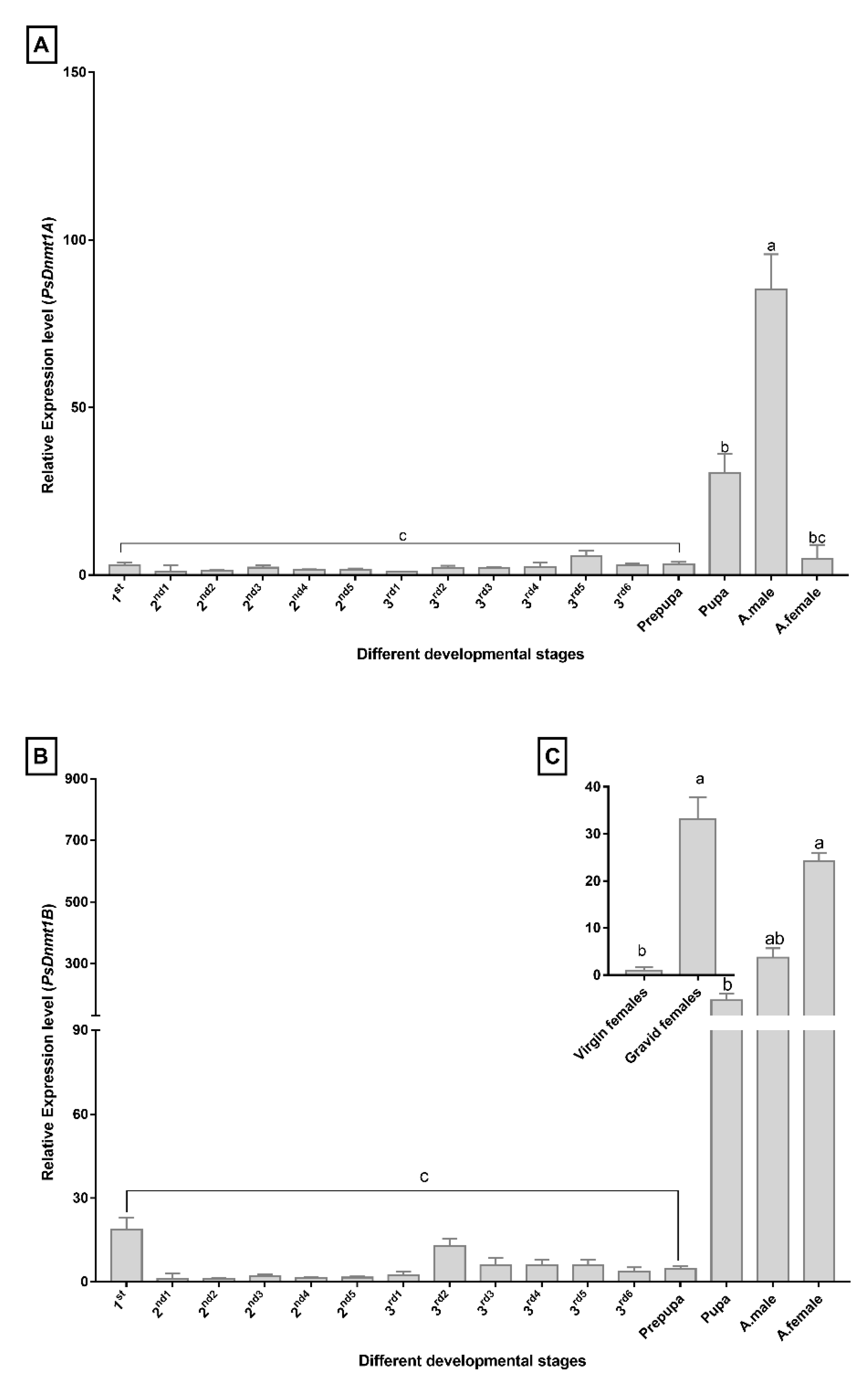
3.4. PsDnmt1A Is Highly Expressed in Males, While Psdnmt1b Is Abundant in Females
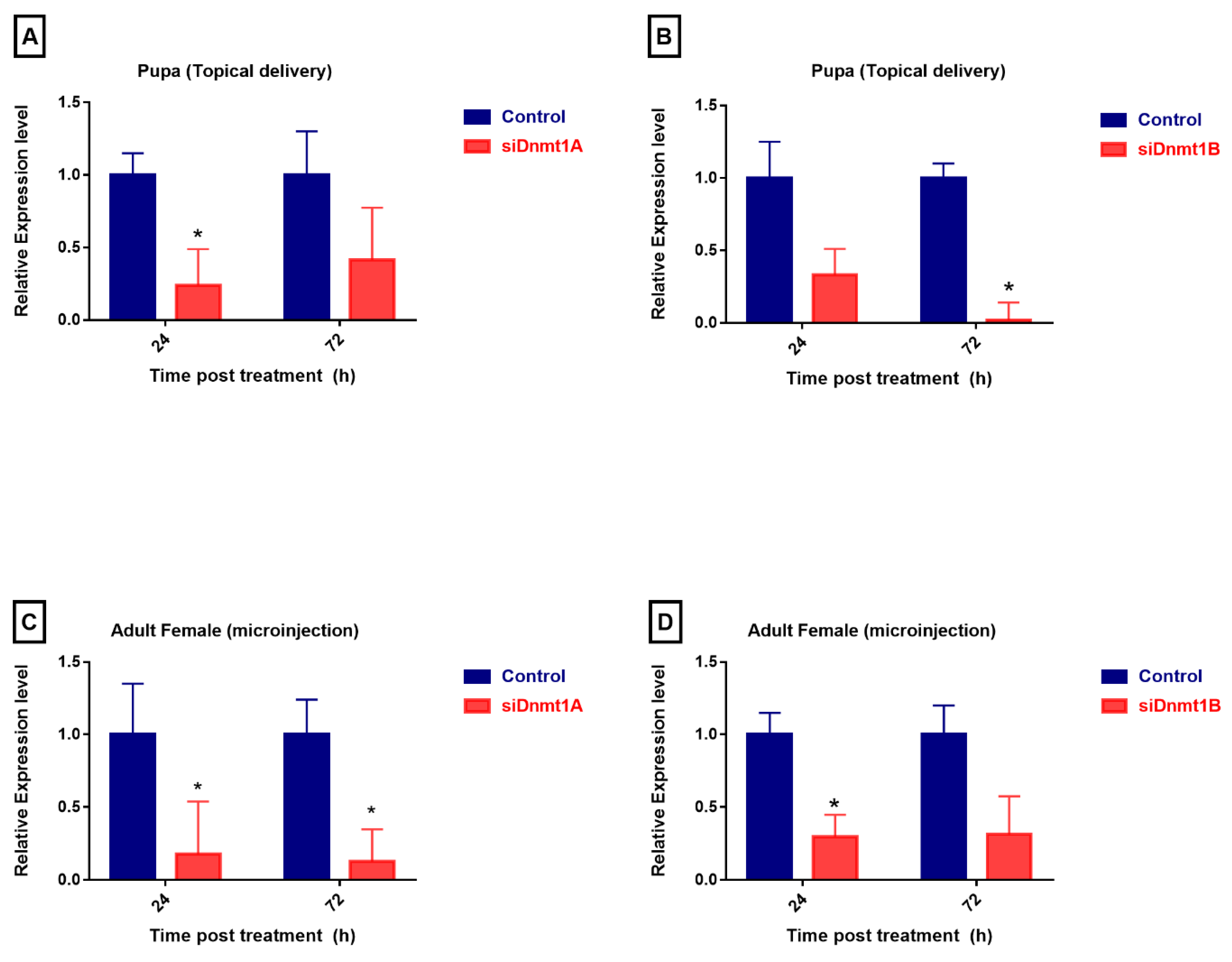
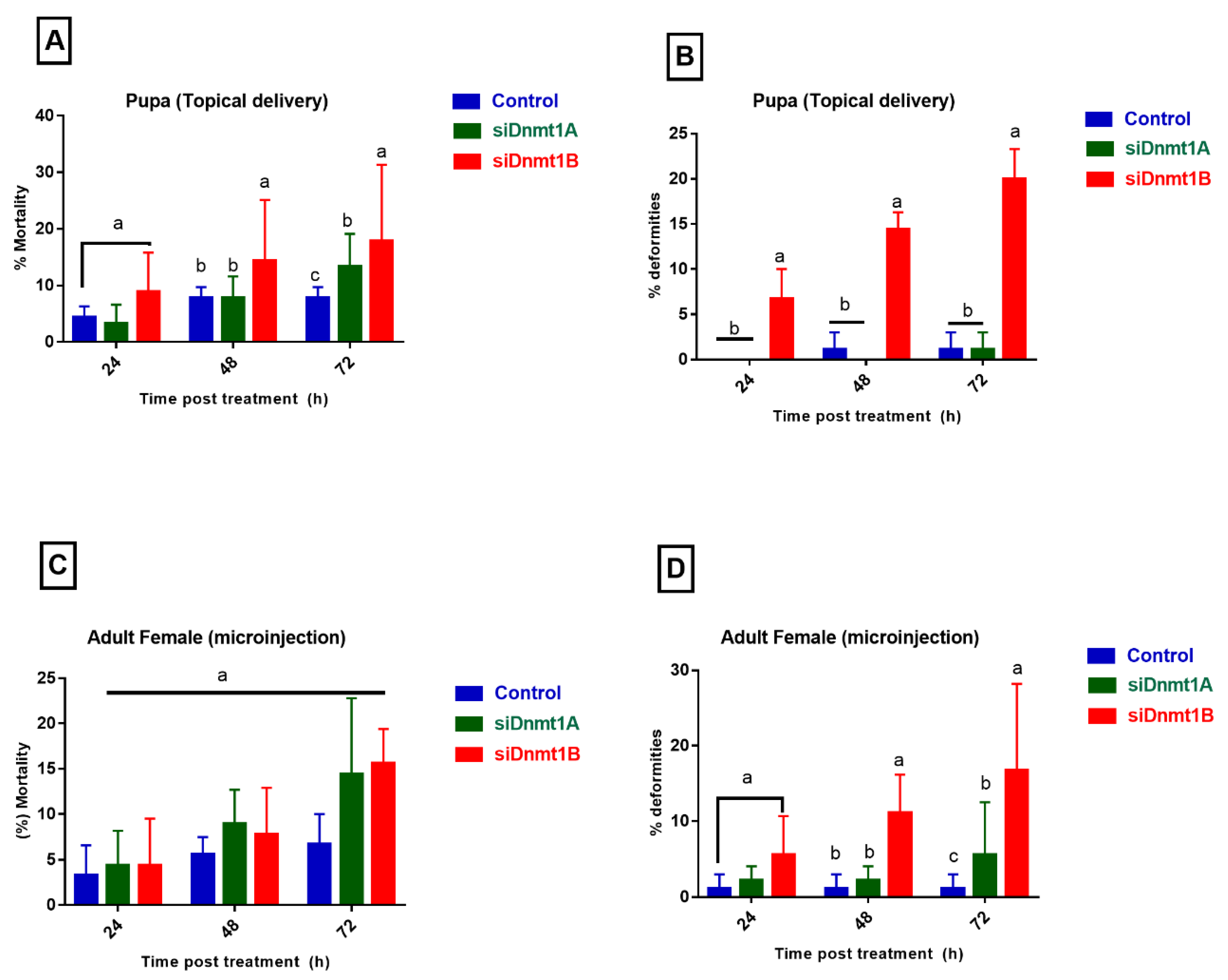
3.5. PsDnmt1B Regulates Wing Development in Adult Males, While Psdnmt1a and 1b Modulate Adult Female Reproduction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bird, A. DNA methylation patterns and epigenetic memory. Gene. Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Laird, P.W. Principles and challenges of genome-wide DNA methylation analysis. Nat. Rev. Genet. 2010, 11, 191–203. [Google Scholar] [CrossRef]
- Robertson, K.D. DNA methylation and human disease. Nat. Rev. Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef]
- Rakyan, V.K.; Down, T.A.; Thorne, N.P.; Flicek, P.; Kulesha, E.; Gräf, S.; Tomazou, E.M.; Bäckdahl, L.; Johnson, N.; Herberth, M. An integrated resource for genome-wide identification and analysis of human tissue-specific differentially methylated regions (tDMRs). Geno. Res. 2008, 18, 1518–1529. [Google Scholar] [CrossRef] [Green Version]
- Doi, A.; Park, I.H.; Wen, B.; Murakami, P.; Aryee, M.J.; Irizarry, R.; Herb, B.; Ladd-Acosta, C.; Rho, J.; Loewer, S. Differential methylation of tissue-and cancer-specific CpG island shores distinguishes human induced pluripotent stem cells, embryonic stem cells and fibroblasts. Nat. Genet. 2009, 41, 1350–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.A. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Goll, M.G.; Bestor, T.H. Eukaryotic cytosine methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glastad, K.M.; Hunt, B.G.; Yi, S.V.; Goodisman, M.A.D. DNA methylation in insects: on the brink of the epigenomic era. Ins. Mol. Bio. 2011, 20, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Bewick, A.J.; Vogel, K.J.; Moore, A.J.; Schmitz, R.J. Evolution of DNA methylation across insects. Mol. Biol. Evol. 2016, 34, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xing, Y.; Li, Y.; Yin, C.; Ge, C.; Li, F. DNA methyltransferases have an essential role in female fecundity in brown planthopper, Nilaparvata lugens. Biochem. Biophys. Res. Commun. 2015, 464, 83–88. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wheeler, D.; Avery, A.; Rago, A.; Choi, J.H.; Colbourne, J.K.; Clark, A.G.; Werren, J.H. Function and Evolution of DNA Methylation in Nasonia vitripennis. PLoS Genet. 2013, 9, e1003872. [Google Scholar] [CrossRef] [Green Version]
- Lyko, F.; Foret, S.; Kucharski, R.; Wolf, S.; Falckenhayn, C.; Maleszka, R. The Honey Bee Epigenomes: Differential Methylation of Brain DNA in Queens and Workers. PLoS Biol. 2010, 8, e1000506. [Google Scholar] [CrossRef] [Green Version]
- Lyko, F.; Maleszka, R. Insects as innovative models for functional studies of DNA methylation. Tren. Genet. 2011, 27, 127–131. [Google Scholar] [CrossRef]
- Marhold, J.; Rothe, N.; Pauli, A.; Mund, C.; Kuehle, K.; Brueckner, B.; Lyko, F. Conservation of DNA methylation in dipteran insects. Ins. Mol. Biol. 2004, 13, 117–123. [Google Scholar] [CrossRef]
- Schaefer, M.; Lyko, F. Lack of evidence for DNA methylation of Invader4 retroelements in Drosophila and implications for Dnmt2-mediated epigenetic regulation. Nat. Genet. 2010, 42, 920–921. [Google Scholar] [CrossRef]
- Feliciello, I.; Parazajder, J.; Akrap, I.; Ugarković, Đ. First evidence of DNA methylation in insect Tribolium castaneum: Environmental regulation of DNA methylation within heterochromatin. Epigenetics 2013, 8, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Consortium, T.G.S. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar]
- Xiang, H.; Zhu, J.; Chen, Q.; Dai, F.; Li, X.; Li, M.; Zhang, H.; Zhang, G.; Li, D.; Dong, Y. Single base–resolution methylome of the silkworm reveals a sparse epigenomic map. Nat. Biot. 2010, 28, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Standage, D.S.; Berens, A.J.; Glastad, K.M.; Severin, A.J.; Brendel, V.P.; Toth, A.L. Genome, transcriptome and methylome sequencing of a primitively eusocial wasp reveal a greatly reduced DNA methylation system in a social insect. Mol. Ecol. 2016, 25, 1769–1784. [Google Scholar] [CrossRef]
- Mandrioli, M.; Volpi, N. The genome of the lepidopteran Mamestra brassicae has a vertebrate-like content of methyl-cytosine. Genetica 2003, 119, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Mendiratta, G.; Brahmachari, V. Genomic imprinting in the mealybugs. Cytogenetic Geno. Res. 2006, 113, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Scarbrough, K.; Hattman, S.; Nur, U. Relationship of DNA methylation level to the presence of heterochromatin in mealybugs. Mol. Cell. Biol. 1984, 4, 599–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, N.K.E.; Wagner, C.I.; Ebeling, J.; Raddatz, G.; Diddens-de Buhr, M.F.; Lyko, F.; Kurtz, J. Dnmt1 has an essential function despite the absence of CpG DNA methylation in the red flour beetle Tribolium castaneum. Sci. Rep. 2018, 8, 16462. [Google Scholar] [CrossRef]
- Satnam, S.; Brar, J.; Suneet, P.; Gill, J.; Sukhjinder, K.; Jeyakumar, P.; Bambawale, O. Impact of area wide management of cotton mealybug, Phenacoccus solenopsis (Tinsely) in Faridkot district of Punjab. J. Cott. Res. Devel. 2012, 26, 214–218. [Google Scholar]
- Ma, L.; Cao, L.J.; Gong, Y.J.; Hoffmann, A.A.; Zeng, A.P.; Wei, S.J.; Zhou, Z.S. Development of novel microsatellites for population genetic analysis of Phenacoccus solenopsis Tinsley (Hemipeta: Pseudoccoccidae) based on genomic analysis. Int. J. Biol. Macromol. 2019, 121, 1135–1144. [Google Scholar] [CrossRef]
- Arya, S.K.; Jain, G.; Upadhyay, S.K.; Singh, H.; Dixit, S.; Verma, P.C. Reference genes validation in Phenacoccus solenopsis under various biotic and abiotic stress conditions. Sci. Rep. 2017, 7, 13520. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, C.; Abbas, G.; Arif, M.J.; Saeed, S.; Karar, H. Phenacoccus solenopsis Tinsley (Sternorrhyncha: Coccoidea: Pseudococcidae), an invasive mealybug damaging cotton in Pakistan and India, with a discussion on seasonal morphological variation. Zootaxa 2008, 1913, 1–35. [Google Scholar] [CrossRef]
- Omar, M.A.; Ao, Y.; Li, M.; He, K.; Xu, L.; Tong, H.; Jiang, M.; Li, F. The functional difference of eight chitinase genes between male and female of the cotton mealybug, Phenacoccus solenopsis. Ins. Mol. Biol. 2019, 28, 550–567. [Google Scholar] [CrossRef]
- Vea, I.M.; Tanaka, S.; Shiotsuki, T.; Jouraku, A.; Tanaka, T.; Minakuchi, C. Differential Juvenile Hormone Variations in Scale Insect Extreme Sexual Dimorphism. PLoS ONE 2016, 11, e0149459. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information (NCBI). Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 10 June 2019).
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Elsevier, 1965; pp. 97–166. [Google Scholar]
- GenePharma Co., Ltd. (Shanghai, China). Available online: http://www.genepharma.com/en/ (accessed on 16 July 2019).
- Niu, J.; Yang, W.J.; Tian, Y.; Fan, J.Y.; Ye, C.; Shang, F.; Ding, B.Y.; Zhang, J.; An, X.; Yang, L.; et al. Topical dsRNA delivery induces gene silencing and mortality in the pea aphid. Pest Manag. Sci. 2019, 75, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Callebaut, I.; Courvalin, J.C.; Mornon, J.P. The BAH (bromo-adjacent homology) domain: A link between DNA methylation, replication and transcriptional regulation. FEBS Lett. 1999, 446, 189–193. [Google Scholar] [CrossRef]
- Arif, M.I.; Rafiq, M.; Ghaffar, A. Host plants of cotton mealybug (Phenacoccus solenopsis): A new menace to cotton agroecosystem of Punjab, Pakistan. Inter. J. Agri. Bio. 2009, 11, 163–167. [Google Scholar]
- Ibrahim, S.S.; Moharum, F.A.; El-Ghany, N.M.A. The cotton mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) as a new insect pest on tomato plants in Egypt. J. Pl. Prot. Res. 2015, 55, 48–51. [Google Scholar] [CrossRef]
- Tinsley, J. Notes on Coccidae with descriptions of new species. Canad. Entom. 1898, 30, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jorda, M.; Jones, P.L.; Maleszka, R.; Ling, X.; Robertson, H.M.; Mizzen, C.A.; Peinado, M.A.; Robinson, G.E. Functional CpG methylation system in a social insect. Science 2006, 314, 645–647. [Google Scholar] [CrossRef]
- Bewick, A.J.; Sanchez, Z.; McKinney, E.C.; Moore, A.J.; Moore, P.J.; Schmitz, R.J. Dnmt1 is essential for egg production and embryo viability in the large milkweed bug. Oncopeltus Fasciatus. Epig. Chrom. 2019, 12, 6. [Google Scholar] [CrossRef]
- Mitsudome, T.; Mon, H.; Xu, J.; Li, Z.; Lee, J.M.; Patil, A.A.; Masuda, A.; Iiyama, K.; Morokuma, D.; Kusakabe, T. Biochemical characterization of maintenance DNA methyltransferase DNMT-1 from silkworm, Bombyx mori. Ins. Bioch. Mol. Biol. 2015, 58, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Patalano, S.; Vlasova, A.; Wyatt, C.; Ewels, P.; Camara, F.; Ferreira, P.G.; Asher, C.L.; Jurkowski, T.P.; Segonds-Pichon, A.; Bachman, M. Molecular signatures of plastic phenotypes in two eusocial insect species with simple societies. Proc. Nat. Acad. Sci. USA 2015, 112, 13970–13975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, B.G.; Glastad, K.M.; Yi, S.V.; Goodisman, M.A. Patterning and regulatory associations of DNA methylation are mirrored by histone modifications in insects. Geno. Biol. Evol. 2013, 5, 591–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Chen, X.M.; Liu, W.W.; Feng, Y.; Sun, T. Transcriptome analysis of sexually dimorphic Chinese white wax scale insects reveals key differences in developmental programs and transcription factor expression. Sci. Rep. 2015, 5, 8141. [Google Scholar] [CrossRef] [Green Version]
- Arakane, Y.; Dixit, R.; Begum, K.; Park, Y.; Specht, C.A.; Merzendorfer, H.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Analysis of functions of the chitin deacetylase gene family in Tribolium castaneum. Ins. Bioch. Mol. Biol. 2009, 39, 355–365. [Google Scholar] [CrossRef]
- Khan, A.M.; Ashfaq, M.; Khan, A.A.; Naseem, M.T.; Mansoor, S. Evaluation of potential RNA-interference-target genes to control cotton mealybug, Phenacoccus solenopsis (Hemiptera: Pseudococcuidae). Insect Sci. 2017, 25, 778–786. [Google Scholar] [CrossRef]
- Khan, A.M.; Ashfaq, M.; Kiss, Z.; Khan, A.A.; Mansoor, S.; Falk, B.W. Use of recombinant tobacco mosaic virus to achieve RNA interference in plants against the citrus mealybug, Planococcus citri (Hemiptera: Pseudococcidae). PLoS ONE 2013, 8, e73657. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, M.; Pandher, S.; Kaur, G.; Goel, N.; Rathore, P. Using de novo transcriptome assembly and analysis to study RNAi in Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Sci. Rep. 2019, 9, 13710. [Google Scholar] [CrossRef]
- Gong, Y.H.; Yu, X.R.; Shang, Q.L.; Shi, X.Y.; Gao, X.W. Oral delivery mediated RNA interference of a carboxylesterase gene results in reduced resistance to organophosphorus insecticides in the cotton Aphid, Aphis gossypii Glover. PLoS ONE 2014, 9, e102823. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S.; Huvenne, H.; Smagghe, G. Delivery of dsRNA for RNAi in insects: an overview and future directions. Insect Sci. 2013, 20, 4–14. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, C.; von Wettstein, D.; Liu, B. Tissue-specific differences in cytosine methylation and their association with differential gene expression in sorghum. Plant Physi. 2011, 156, 1955–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, P.; Song, F.; Ghosh, S.; Morien, E.; Qin, M.; Mahmood, S.; Fujiwara, K.; Igarashi, J.; Nagase, H.; Held, W.A. Genome-wide survey reveals dynamic widespread tissue-specific changes in DNA methylation during development. BMC Geno. 2011, 12, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, F.; Smith, J.F.; Kimura, M.T.; Morrow, A.D.; Matsuyama, T.; Nagase, H.; Held, W.A. Association of tissue-specific differentially methylated regions (TDMs) with differential gene expression. Proc. Nat. Acad. Sci. USA 2005, 102, 3336–3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Zhang, J.; Lyu, H.; Song, Q.; Feng, Q.; Xiang, H.; Zheng, S. DNA methylation mediates BmDeaf1-regulated tissue-and stage-specific expression of BmCHSA-2b in the silkworm, Bombyx mori. Epig. Chrom. 2018, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sense (5′–3′) | Antisense (5′–3′) | Purpose |
|---|---|---|---|
| DNMT1A | AGGACATCTGTGCCCATTC | CGACCATAAGTTGAGTCGTATCT | PCR |
| DNMT1A (1) | ATGAAGCAGGACATCTGTGCCCATTCG | GGAACGCCTCCTGTTGCATCGGA | RACE “1st round” |
| DNMT1A (2) | ATGCAACAGGAGGCGTTCCTATTAGACG | CCTTTTACGGCGATTGGTTCTCCAATCC | RACE “2nd round” |
| qDT1A | TACGCTGCTGGTTACGTTAAA | ACCGCTTCTTCACCTTCATC | qRT-PCR |
| DNMT1A-phso-490 | GCCAGAGAAUUUAAAGGAUTT | AUCCUUUAAAUUCUCUGGCTT | siRNA synthesis |
| DNMT1A-phso-1542 | CCAAGAAUUGACGUCGAUUTT | AAUCGACGUCAAUUCUUGGTT | siRNA synthesis |
| DNMT1A-phso-2093 | GCUAGAUACGACUCAACUUTT | AAGUUGAGUCGUAUCUAGCTT | siRNA synthesis |
| DNMT1B | GAAGGTTTGCATCAAGCGGG | CCCATTGGTTCTGGTTGGGT | PCR |
| DNMT1B (1) | TCAGAGCAGGCGAAGGGCAATCA | TTGGCACGGAGGGCCACCGCACA | RACE “1st round” |
| DNMT1B (2) | GCATCAAGCGGGAATAGCGGAGTG | GCAAACCTTCTGTCAATCCTCCGCAAC | RACE “2nd round” |
| qDT1B | GGTCATCATCTGCTCCGTTAC | GGGATCGTGCTTATGAGGTATTT | qRT-PCR |
| DNMT1B-phso-1376 | GCCAAGGAUUCAGUGGGAUTT | AUCCCACUGAAUCCUUGGCTT | siRNA synthesis |
| DNMT1B-phso-1771 | GCUCCGUUACGGAUGCUAATT | UUAGCAUCCGUAACGGAGCTT | siRNA synthesis |
| DNMT1B-phso-2206 | GCUAAUCGGCAUGGUAAUUTT | AAUUACCAUGCCGAUUAGCTT | siRNA synthesis |
| Negative control | UUCUCCGAACGUGUCACGUTT | ACGUGACACGUUCGGAGAATT | siRNA synthesis |
| PsActin | TCGTACCACCGGTATCGTATTA | TTAAGTCACGACCAGCCAAG | qRT-PCR |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omar, M.A.A.; Li, M.; Liu, F.; He, K.; Qasim, M.; Xiao, H.; Jiang, M.; Li, F. The Roles of DNA Methyltransferases 1 (DNMT1) in Regulating Sexual Dimorphism in the Cotton Mealybug, Phenacoccus solenopsis. Insects 2020, 11, 121. https://doi.org/10.3390/insects11020121
Omar MAA, Li M, Liu F, He K, Qasim M, Xiao H, Jiang M, Li F. The Roles of DNA Methyltransferases 1 (DNMT1) in Regulating Sexual Dimorphism in the Cotton Mealybug, Phenacoccus solenopsis. Insects. 2020; 11(2):121. https://doi.org/10.3390/insects11020121
Chicago/Turabian StyleOmar, Mohamed A.A., Meizhen Li, Feiling Liu, Kang He, Muhammad Qasim, Huamei Xiao, Mingxing Jiang, and Fei Li. 2020. "The Roles of DNA Methyltransferases 1 (DNMT1) in Regulating Sexual Dimorphism in the Cotton Mealybug, Phenacoccus solenopsis" Insects 11, no. 2: 121. https://doi.org/10.3390/insects11020121