Larval Host Preference and Suitability for the Sawfly Mesoneura rufonota among Five Cinnamomun Tree Species

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Insects Rearing
2.3. Larval Preference Experiments
2.3.1. Larval Feeding Rate across Host Plants
2.3.2. Larval Choice Behavior across Host Plants
2.4. Larval Performance Experiments
2.5. Statistical Analysis
3. Results
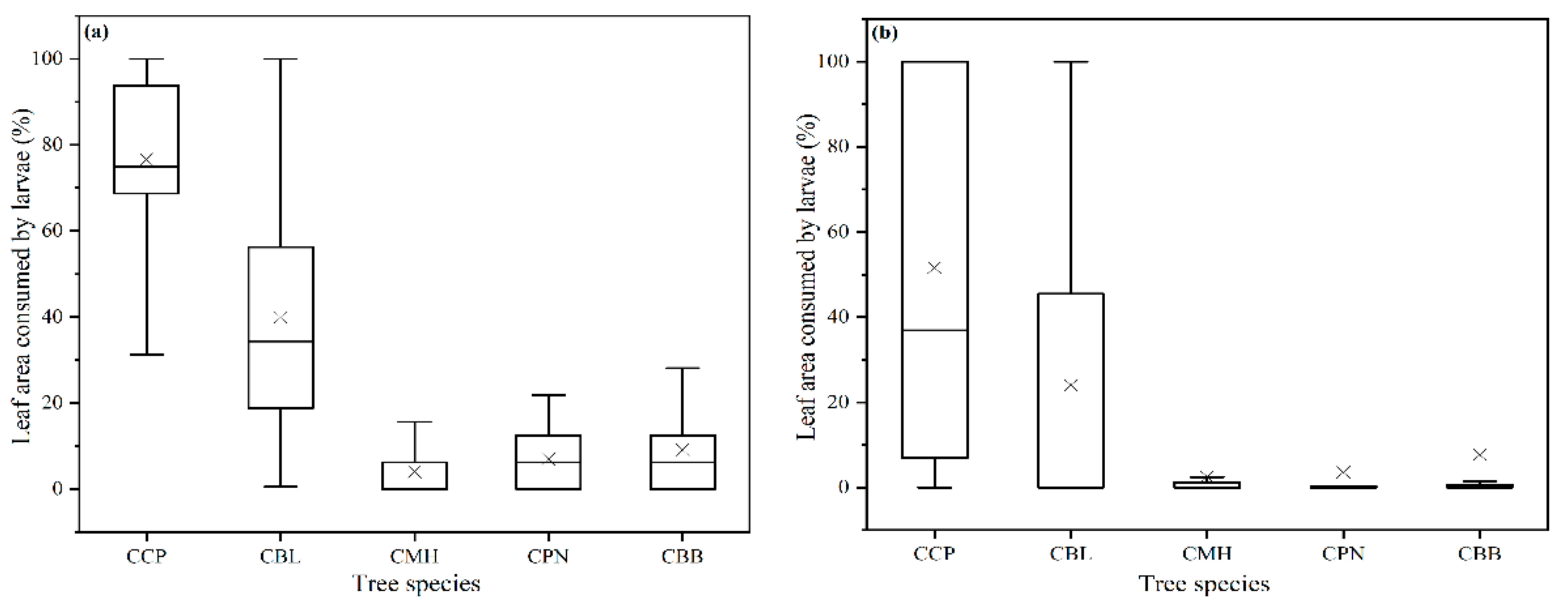
3.1. Larval Feeding Rate across Host Plants
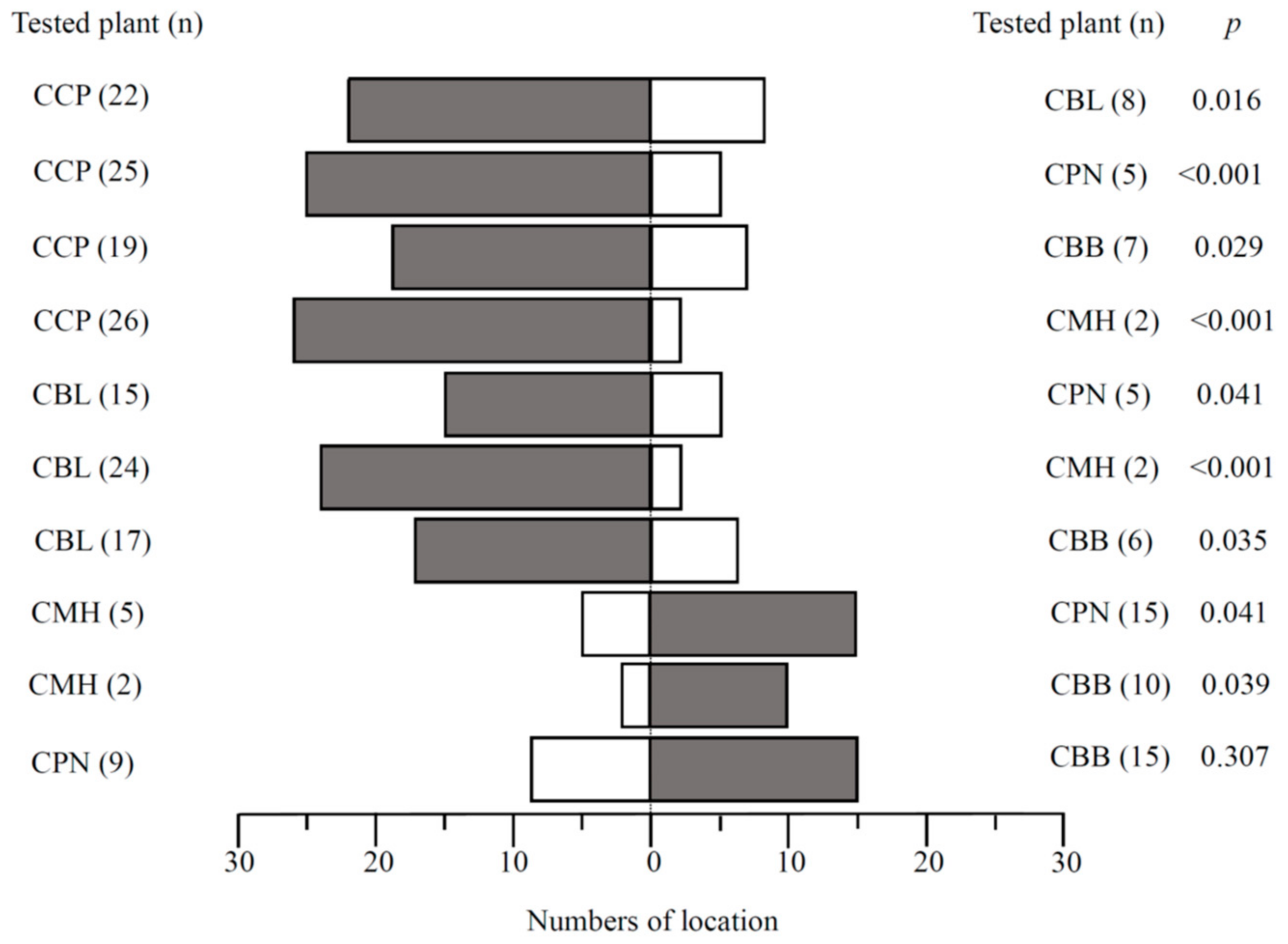
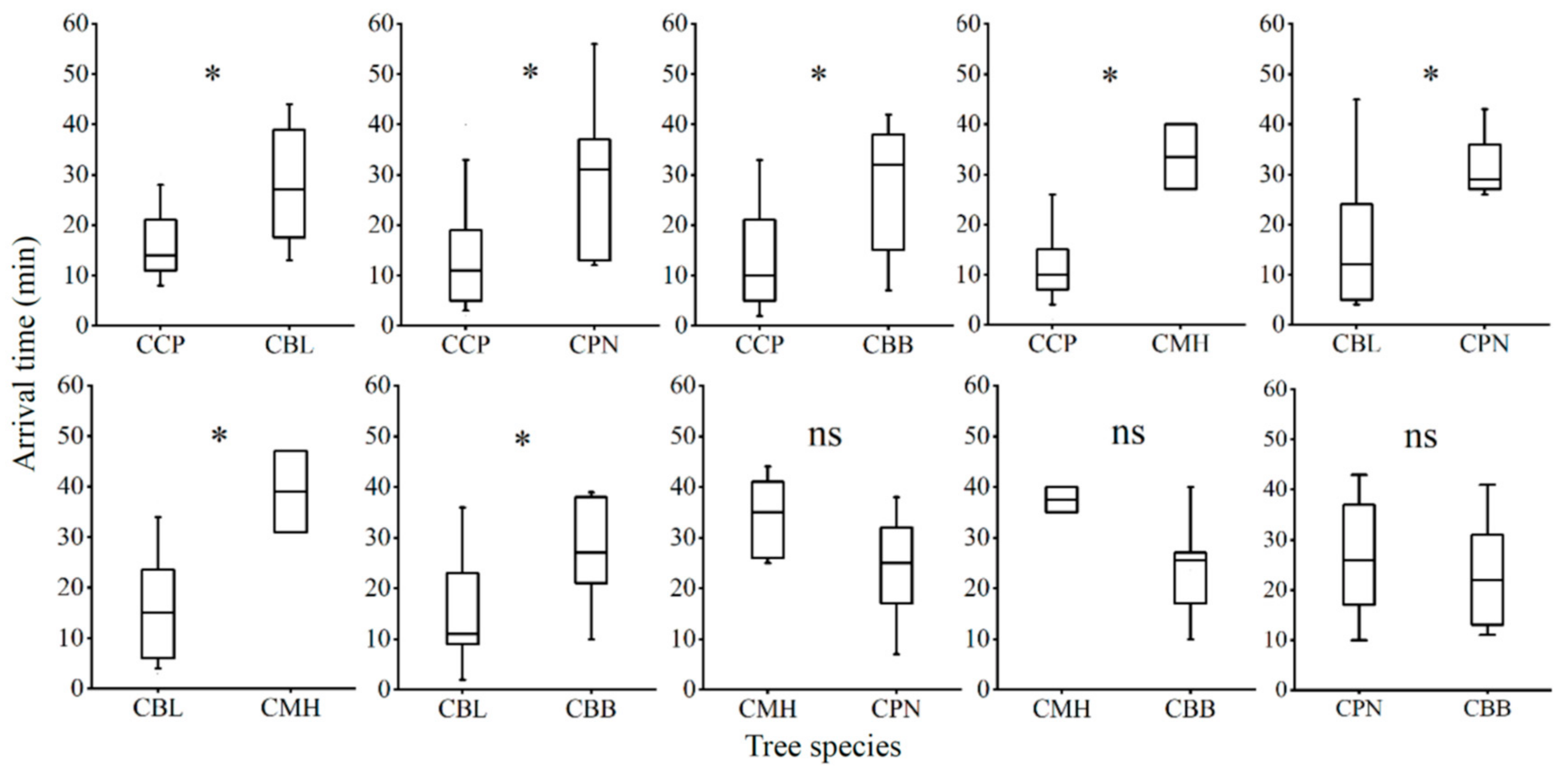
3.2. Larval Choice Behavior across Host Plants
3.3. Larval Performance across Host Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaenike, J. Host specialization in phytophagous insects. Annu. Rev. Ecol. Syst. 1990, 21, 243–273. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Thöming, G.; Larsson, M.C.; Hansson, B.S.; Anderson, P. Comparison of plant preference hierarchies of male and female moths and the impact of larval rearing hosts. Ecology 2013, 94, 1744–1752. [Google Scholar] [CrossRef]
- Luo, D.; Lai, M.; Xu, C.F.; Shi, H.N.; Liu, X.P. Life history traits in a capital breeding pine caterpillar: Effect of host species and needle age. BMC Ecol. 2018, 18, 24. [Google Scholar] [CrossRef] [Green Version]
- Novotny, V.; Basset, Y.; Miller, S.E.; Drozd, P.; Cizek, L. Host specialization of leaf-chewing insects in a New Guinea rainforest. J. Anim. Ecol. 2002, 71, 400–412. [Google Scholar] [CrossRef]
- Jermy, T. Evolution of insect-host plant relationships. Am. Nat. 1984, 124, 609–630. [Google Scholar] [CrossRef]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T. A meta-analysis of preference-performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef]
- Hafsi, A.; Facon, B.; Ravigné, V.; Chiroleu, F.; Quilici, S.; Chermiti, B.; Duyck, P.F. Host plant range of a fruit fly community (Diptera: Tephritidae): Does fruit composition influence larval performance? BMC Ecol. 2016, 16, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.P.; Raubenheimer, D.; Behmer, S.T.; Simpson, S.J. A correlation between macronutrient balancing and insect host-plant range: Evidence from the specialist caterpillar Spodoptera exempta (Walker). J. Insect Physiol. 2003, 49, 1161–1171. [Google Scholar] [CrossRef]
- Sylla, S.; Brévault, T.; Monticelli, L.S.; Diarra, K.; Desneux, N. Geographic variation of host preference by the invasive tomato leaf miner Tuta absoluta: Implications for host range expansion. J. Pest Sci. 2019, 92, 1387–1396. [Google Scholar] [CrossRef]
- Bernays, E.A.; Chapman, R.F. Host-Plant Selection by Phytophagous Insects; Chapman & Hall: New York, NY, USA, 1994. [Google Scholar]
- Jiao, X.G.; Xie, W.; Wang, S.L.; Wu, Q.J.; Zhou, L.; Pan, H.P.; Liu, B.M.; Zhang, Y.J. Host preference and nymph performance of B and Q putative species of Bemisia tabaci on three host plants. J. Pest Sci. 2012, 85, 423–430. [Google Scholar] [CrossRef]
- Ahmad, S. Herbivorous Insects: Host-Seeking Behavior and Mechanisms; Academic Press: New York, NY, USA, 1983. [Google Scholar]
- Bruce, T.J.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]
- West, S.A.; Cunningham, J.P. A general model for host plant selection in phytophagous insects. J. Theor. Biol. 2002, 214, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Liu, T.X. Oviposition preference, larval performance and adaptation of Trichoplusia ni on cabbage and cotton. Insect Sci. 2014, 22, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Coapio, G.G.; Cruz-López, L.; Guerenstein, P.; Malo, E.A.; Rojas, J.C. Oviposition preference and larval performance and behavior of Trichoplusia ni (Lepidoptera: Noctuidae) on host and nonhost plants. Arthropod Plant Interact. 2017, 12, 267–276. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Clarke, A.R.; Malcolm, S.B. Ecology and behavior of first instar larval Lepidoptera. Ann. Rev. Entomol. 2002, 47, 361–393. [Google Scholar] [CrossRef] [PubMed]
- Soler, R.; Pineda, A.; Li, Y.; Ponzio, C.; van Loon, J.J.A.; Weldegergis, B.T.; Dicke, M. Neonates know better than their mothers when selecting a host plant. Oikos 2012, 121, 1923–1934. [Google Scholar] [CrossRef]
- Rosenwald, L.C.; Lill, J.T.; Lind, E.M.; Weiss, M.R. Dynamics of host plant selection and host-switching by silver spotted skipper caterpillars. Arthropod Plant Interact. 2017, 11, 833–842. [Google Scholar] [CrossRef]
- McCormick, A.C.; Arrigo, L.; Eggenberger, H.; Mescher, M.C.; De Moraes, C.M. Divergent behavioural responses of gypsy moth (Lymantria dispar) caterpillars from three different subspecies to potential host trees. Sci. Rep. 2019, 9, 8953. [Google Scholar] [CrossRef] [Green Version]
- Prasad, K.N.; Yang, B.; Dong, X.H.; Jiang, G.X.; Zhang, H.Y.; Xie, H.H.; Jiang, Y.M. Flavonoid contents and antioxidant activities from Cinnamomum species. Innov. Food Sci. Emerg. Technol. 2009, 4, 627–632. [Google Scholar] [CrossRef]
- Guo, S.S.; Geng, Z.F.; Zhang, W.J.; Liang, J.Y.; Wang, C.F.; Deng, Z.W.; Du, S.S. The chemical composition of essential oils from Cinnamomum camphora and their insecticidal activity against the stored product pests. Int. J. Mol. Sci. 2016, 17, 1836. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J. Study on the camphor sawfly Mesonurae rufonota (Rohwer). Sci. Silv. Sin. 1962, 8, 184–186, (In Chinese with English summary). [Google Scholar]
- Wu, J.F. Biology and control of the Mesoneura rufonota (Rohwer). Acta. Entomol. Sin. 1982, 25, 42–48, (In Chinese with English summary). [Google Scholar]
- Chang, K.P.; Hsiau, L.J.; Hsiau, W.F. The studies on the morphology and life cycle of camphor tree leaf bee (Mesonurae rufonota (Rohwer)). Plant Prot. Bull. 1998, 40, 287–295. [Google Scholar]
- Xu, C.F.; Luo, D.; Yin, L.X.; Liu, X.P. Influence of temperature on the development and reproduction of the camphor sawfly, Mesoneura rufonota (Hymenoptera: Tenthredinidae). Acta. Entomol. Sin. 2017, 60, 1216–1225, (In Chinese with English summary). [Google Scholar]
- Singer, M.C. The definition and measurement of oviposition preference in plant-feeding insects. In Insect-Plant Interactions; Miller, T.A., Miller, J., Eds.; Springer: New York, NY, USA, 1986; pp. 65–94. [Google Scholar]
- Thompson, J.N. Evolutionary ecology of the relationship between oviposition preference and performance of offspring in phytophagous insects. Entomol. Exp. Appl. 1988, 47, 3–14. [Google Scholar] [CrossRef]
- Rajapakse, C.N.K.; Walter, G.H. Polyphagy and primary host plants: Oviposition preference versus larval performance in the lepidopteran pest Helicoverpa armigera. Arthropod Plant Interact. 2007, 1, 17–26. [Google Scholar] [CrossRef]
- Shikano, I.; Akhtar, Y.; Isman, M.B. Relationship between adult and larval host plant selection and larval performance in the generalist moth, Trichoplusia ni. Arthropod Plant Interact. 2010, 4, 197–205. [Google Scholar] [CrossRef]
- Midega, C.A.O.; Khan, Z.R.; Pickett, J.A. Host plant selection behaviour of Chilo partellus and its implication for effectiveness of a trap crop. Entomol. Exp. Appl. 2011, 138, 40–47. [Google Scholar] [CrossRef]
- Dethier, V.G. Evolution of feeding preferences in phytophagous insects. Evolution 1954, 8, 33–54. [Google Scholar] [CrossRef]
- Rostás, M.; Eggert, K. Ontogenetic and spatio-temporal patterns of induced volatiles in glycine max in the light of the optimal defense hypothesis. Chemoecology 2008, 18, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhi, J.; Cong, C.; Margolies, D.C. Olfactory cues used in host selection by Frankliniella occidentalis (Thysanoptera: Thripidae) in relation to host suitability. J. Insect Behav. 2013, 27, 41–56. [Google Scholar] [CrossRef]
- Yang, J.; Sadof, C.S. Variegation in Coleus blumei and the life history of citrus mealybug (Homoptera: Pseudococcidae). Environ. Entomol. 1995, 24, 1650–1655. [Google Scholar] [CrossRef]
- Tanga, C.M.; Ekesi, S.; Govender, P.; Mohamed, S.A. Effect of six host plant species on the life history and population growth parameters of Rastrococcus iceryoides (Hemiptera: Pseudococcidae). Fla. Entomol. 2013, 96, 1030–1041. [Google Scholar] [CrossRef] [Green Version]
- Altesor, P.; González, A. Preference-performance in a specialist sawfly on congeneric host plants. Entomol. Exp. Appl. 2018, 166, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Nethononda, P.D.; Nofemela, R.S.; Modise, D.M. Development, survival, body weight and oviposition rates of Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae) when reared on seven cabbage cultivars. Afr. Entomol. 2016, 24, 162–169. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Saeed, R.; Sayyed, A.H.; Shad, S.A.; Zaka, S.M. Effect of different host plants on the fitness of diamond-back moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop Prot. 2010, 29, 178–182. [Google Scholar] [CrossRef]
- Strand, M.R.; Casas, J. Parasitoid and host nutritional physiology in behavioural ecology. In Parasitoid Behavioral Ecology; Wajnberg, E., van Alphen, J., Bernstein, C., Eds.; Blackwell Press: Oxford, UK, 2007; pp. 113–128. [Google Scholar]
- Winkler, K.; Wäckers, F.L.; Kaufman, L.V.; Larraz, V.; van Lenteren, J.C. Nectar exploitation by herbivores and their parasitoids is a function of flower species and relative humidity. Biol. Control. 2009, 50, 299–306. [Google Scholar] [CrossRef]
- Nestel, D.; Papadopoulos, N.T.; Pascacio-Villafán, C.; Righini, N.; Altuzar-Molina, A.R.; Aluja, M. Resource allocation and compensation during development in holometabolous insects. J. Insect Physiol. 2016, 95, 78–88. [Google Scholar] [CrossRef]
- Dostálek, T.; Rokaya, M.B.; Maršík, P.; Rezek, J.; Skuhrovec, J.; Pavela, R.; Münzbergová, Z. Trade-off among different anti-herbivore defence strategies along an altitudinal gradient. AoB Plants 2016, 8, plw026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maag, D.; Erb, M.; Köllner, T.G.; Gershenzon, J. Defensive weapons and defense signals in plants: Some metabolites serve both roles. BioEssays 2015, 37, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Züst, T.; Agrawal, A.A. Trade-offs between plant growth and defense against insect herbivory: An emerging mechanistic synthesis. Annu. Rev. Plant Biol. 2017, 68, 513–534. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.O.; Miller, J.R. Host-acceptance behavior in an herbivorous fly, Delia antiqua. J. Insect Physiol. 1988, 34, 179–190. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tree Species | CCP | CBL | CMH | CPN | |
|---|---|---|---|---|---|
| CBL | Percentage | 97.9 (71.8, 100); 63.9 (30.1, 76.1) | |||
| W | 61.0 ** | ||||
| CMH | Percentage | 100 (100, 100); 0 (0, 0.3) | 91.5 (78.1, 95.6); 0 (0, 0.1) | ||
| W | 0 ** | 9.5 ** | |||
| CPN | Percentage | 100 (100, 100); 2.8 (1.1, 3.8) | 82.5 (72.5, 90); 6.3 (0, 25) | 0 (0, 0); 1.8 (1.2, 4.2) | |
| W | 27.5 ** | 14 ** | 63 ** | ||
| CBB | Percentage | 98.8 (93.6, 100); 0.5 (0, 1.5) | 50 (27.6, 71.8); 0 (0, 2.8) | 0 (0, 0); 0.1 (0, 0.8) | 0.8 (0.4, 1.3); 0.6 (0, 1) |
| W | 41.0 ** | 46.5 ** | 92.5 * | 368 ns | |
| Tree Species | CCP | CBL | CMH | CPN | CBB |
|---|---|---|---|---|---|
| Larval survival (%) | 84.33 ± 2.98a | 45.00 ± 2.59b | 3.67 ± 1.16c | 10.33 ± 1.82c | 10.67 ± 1.71c |
| Pupal survival (%) | 88.88 ± 3.03a | 54.11 ± 3.94b | 0c | 0c | 0c |
| Larval developmental duration (d) | 8.60 ± 0.06b | 8.86 ± 0.08b | 9.27 ± 0.30a | 9.00 ± 0.15ab | 8.97 ± 0.12ab |
| Pupal developmental duration (d) | 7.25 ± 0.05b | 7.73 ± 0.12a | - | - | - |
| Pupal weight (g) | 0.23 ± 0.01a | 0.20 ± 0.01b | 0.16 ± 0.01c | 0.19 ± 0.01b | 0.19 ± 0.01b |
| Adult lifespan (d) | 4.01 ± 0.05a | 3.64 ± 0.10b | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Zhang, J.; Tregenza, T.; Pan, Y.; Wang, Q.; Shi, H.; Liu, X. Larval Host Preference and Suitability for the Sawfly Mesoneura rufonota among Five Cinnamomun Tree Species. Insects 2020, 11, 76. https://doi.org/10.3390/insects11020076
Zhou J, Zhang J, Tregenza T, Pan Y, Wang Q, Shi H, Liu X. Larval Host Preference and Suitability for the Sawfly Mesoneura rufonota among Five Cinnamomun Tree Species. Insects. 2020; 11(2):76. https://doi.org/10.3390/insects11020076
Chicago/Turabian StyleZhou, Jiaying, Jiangtao Zhang, Tom Tregenza, Youliang Pan, Qinzhao Wang, Haoni Shi, and Xingping Liu. 2020. "Larval Host Preference and Suitability for the Sawfly Mesoneura rufonota among Five Cinnamomun Tree Species" Insects 11, no. 2: 76. https://doi.org/10.3390/insects11020076