
Unseen Weapons: Bacterial Extracellular Vesicles and the Spread of Antibiotic Resistance in Aquatic Environments
 , , ,
, , , 
Abstract
:1. Importance and Multifaceted Nature of the Aquatic Environment
2. Antibiotic Resistance (ABR) in Aquatic Environments
3. Potential Negative Effects of ABR in Aquatic Environments on Human and Animal Health
4. Types of ARGs in Aquatic Environment
5. Bacterial Extracellular Vesicles (BEV)
6. Example of a Bacterial EV Responsible for Antibiotic Resistance
7. Potential Strategies to Address ABR in Aquatic Environments Using EVs
8. Future Perspectives
9. Summary and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and threats of contamination on aquatic ecosystems. Bioremediat. Biotechnol. 2020, 1–26. [Google Scholar] [CrossRef]
- Visbeck, M. Ocean science research is key for a sustainable future. Nat. Commun. 2018, 9, 690. [Google Scholar] [CrossRef] [PubMed]
- Nava, V.; Leoni, B. A critical review of interactions between microplastics, microalgae and aquatic ecosystem function. Water Res. 2021, 188, 116476. [Google Scholar] [CrossRef] [PubMed]
- Shivanna, K.R. Climate change and its impact on biodiversity and human welfare. Proc. Indian Natl. Sci. Acad. 2022, 88, 160–171. [Google Scholar] [CrossRef]
- Tang, X.; Xie, G.; Shao, K.; Tian, W.; Gao, G.; Qin, B. Aquatic bacterial diversity, community composition and assembly in the semi-arid inner Mongolia plateau: Combined effects of salinity and nutrient levels. Microorganisms 2021, 9, 208. [Google Scholar] [CrossRef]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in aquaculture. Sci. Rep. 2020, 10, 21878. [Google Scholar] [CrossRef]
- Boyd, C.E.; McNevin, A.A.; Davis, R.P. The contribution of fisheries and aquaculture to the global protein supply. Food Secur. 2022, 14, 805–827. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Rheman, S.; Debnath, N.; Delamare-Deboutteville, J.; Akhtar, Z.; Ghosh, S.; Parveen, S.; Islam, K.; Islam, M.A.; Rashid, M.M.; et al. Antibiotics usage practices in aquaculture in Bangladesh and their associated factors. One Health 2022, 15, 100445. [Google Scholar] [CrossRef] [PubMed]
- Tangcharoensathien, V.; Sommanustweechai, A.; Chanthong, B.; Sumpradit, N.; Sakulbumrungsil, R.; Jaroenpoj, S.; Sermsinsiri, V.; Thai SAC Working Group. Surveillance of antimicrobial consumption: Methodological review for systems development in Thailand. J. Glob. Health 2017, 7, 010307. [Google Scholar] [PubMed]
- Lenz, K.D.; Klosterman, K.E.; Mukundan, H.; Kubicek-Sutherland, J.Z. Macrolides: From toxins to therapeutics. Toxins 2021, 13, 347. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef] [PubMed]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in food and feedingstuffs: From regulation to analytical methods, bacterial resistance, and environmental and health implications. J. Anal. Methods Chem. 2017, 2017, 1315497. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Mao, Y.; Qiu, W.; Wu, Y.; Cai, B. The source and distribution of tetracycline antibiotics in China: A review. Toxics 2023, 11, 214. [Google Scholar] [CrossRef] [PubMed]
- Ben, Y.; Fu, C.; Hu, M.; Liu, L.; Wong, M.H.; Zheng, C. Human health risk assessment of antibiotic resistance associated with antibiotic residues in the environment: A review. Environ. Res. 2019, 169, 483–493. [Google Scholar] [CrossRef]
- Liang, X.; Chen, B.; Nie, X.; Shi, Z.; Huang, X.; Li, X. The distribution and partitioning of common antibiotics in water and sediment of the Pearl River Estuary, South China. Chemosphere 2013, 92, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Polianciuc, S.I.; Gurzău, A.E.; Kiss, B.; Ştefan, M.G.; Loghin, F. Antibiotics in the environment: Causes and consequences. Med. Pharm. Rep. 2020, 93, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Murata, A.; Takada, H.; Mutoh, K.; Hosoda, H.; Harada, A.; Nakada, N. Nationwide monitoring of selected antibiotics: Distribution and sources of sulfonamides, trimethoprim, and macrolides in Japanese rivers. Sci. Total Environ. 2011, 409, 5305–5312. [Google Scholar] [CrossRef]
- Baba, H.; Nishiyama, M.; Watanabe, T.; Kanamori, H. Review of antimicrobial resistance in wastewater in Japan: Current challenges and future perspectives. Antibiotics 2022, 11, 849. [Google Scholar] [CrossRef]
- Medina-Pizzali, M.L.; Hartinger, S.M.; Salmon-Mulanovich, G.; Larson, A.; Riveros, M.; Mäusezahl, D. Antimicrobial resistance in rural settings in Latin America: A scoping review with a one health lens. Int. J. Environ. Res. Public Health 2021, 18, 9837. [Google Scholar] [CrossRef]
- Taneja, N.; Sharma, M. Antimicrobial resistance in the environment: The Indian scenario. Indian J. Med. Res. 2019, 149, 119–128. [Google Scholar] [CrossRef]
- Hanna, N.; Tamhankar, A.J.; Stålsby Lundborg, C. Antibiotic concentrations and antibiotic resistance in aquatic environments of the WHO Western Pacific and South-East Asia regions: A systematic review and probabilistic environmental hazard assessment. Lancet Planet. Health 2023, 7, e45–e54. [Google Scholar] [CrossRef]
- Waleng, N.J.; Nomngongo, P.N. Occurrence of pharmaceuticals in the environmental waters: African and Asian perspectives. Environ. Chem. Ecotoxicol. 2022, 4, 50–66. [Google Scholar] [CrossRef]
- Okeke, E.S.; Chukwudozie, K.I.; Nyaruaba, R.; Ita, R.E.; Oladipo, A.; Ejeromedoghene, O.; Atakpa, E.O.; Agu, C.V.; Okoye, C.O. Antibiotic resistance in aquaculture and aquatic organisms: A review of current nanotechnology applications for sustainable management. Environ. Sci. Pollut. Res. Int. 2022, 29, 69241–69274. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The rising tide of antimicrobial resistance in aquaculture: Sources, sinks and solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [PubMed]
- Maghsodian, Z.; Sanati, A.M.; Mashifana, T.; Sillanpää, M.; Feng, S.; Nhat, T.; Ramavandi, B. Occurrence and distribution of antibiotics in the water, sediment, and biota of freshwater and marine environments: A review. Antibiotics 2022, 11, 1461. [Google Scholar] [CrossRef]
- Yang, Q.; Gao, Y.; Ke, J.; Show, P.L.; Ge, Y.; Liu, Y.; Guo, R.; Chen, J. Antibiotics: An overview on the environmental occurrence, toxicity, degradation, and removal methods. Bioengineered 2021, 12, 7376–7416. [Google Scholar] [CrossRef] [PubMed]
- Ezzariai, A.; Hafidi, M.; Khadra, A.; Aemig, Q.; El Fels, L.; Barret, M.; Merlina, G.; Patureau, D.; Pinelli, E. Human and veterinary antibiotics during composting of sludge or manure: Global perspectives on persistence, degradation, and resistance genes. J. Hazard. Mater. 2018, 359, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Daeseleire, E.; De Graef, E.; Rasschaert, G.; De Mulder, T.; Van den Meersche, T.; Van Coillie, E.; Dewulf, J.; Heyndrickx, M. Antibiotic use and resistance in animals: Belgian initiatives. Drug Test. Anal. 2016, 8, 549–555. [Google Scholar] [CrossRef]
- Santos, L.; Ramos, F. Antimicrobial resistance in aquaculture: Current knowledge and alternatives to tackle the problem. Int. J. Antimicrob. Agents 2018, 52, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.; Kering, K.; Wairimu, C.; Onsare, R.; Mbae, C. Antimicrobial resistance rates and surveillance in Sub-Saharan Africa: Where are we now? Infect. Drug Resist. 2022, 15, 3589–3609. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Cheng, Z.; Zhang, X.; Lv, C.; Guo, C.; Liu, H.; Dong, K.; Zhang, Y.; Liu, C.; Chang, Y.F.; et al. Correction: Global antimicrobial resistance: A system-wide comprehensive investigation using the Global One Health Index. Infect. Dis. Poverty 2022, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, D. Antibiotic resistance, meat consumption and the harm principle. Ethics Policy Environ. 2023, 26, 53–68. [Google Scholar] [CrossRef]
- Suyamud, B.; Chen, Y.; Quyen, D.T.T.; Dong, Z.; Zhao, C.; Hu, J. Antimicrobial resistance in aquaculture: Occurrence and strategies in Southeast Asia. Sci. Total Environ. 2024, 907, 167942. [Google Scholar] [CrossRef] [PubMed]
- Nappier, S.P.; Liguori, K.; Ichida, A.M.; Stewart, J.R.; Jones, K.R. Antibiotic resistance in recreational waters: State of the science. Int. J. Environ. Res. Public Health. 2020, 17, 8034. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.C.; Rodrigues, S.; Afonso, A.; Nogueira, A.; Coutinho, P. Antibiotic resistance in the drinking water: Old and new strategies to remove antibiotics, resistant bacteria, and resistance genes. Pharmaceuticals 2022, 15, 393. [Google Scholar] [CrossRef]
- Zhang, K.; Li, K.; Xin, R.; Han, Y.; Guo, Z.; Zou, W.; Wei, W.; Cui, X.; Zhang, Z.; Zhang, Y. Antibiotic resistomes in water supply reservoirs sediments of central China: Main biotic drivers and distribution pattern. Environ. Sci. Pollut. Res. Int. 2022, 29, 37712–37721. [Google Scholar] [CrossRef]
- Ahmed, H.; Zolfo, M.; Williams, A.; Ashubwe-Jalemba, J.; Tweya, H.; Adeapena, W.; Labi, A.K.; Adomako, L.A.B.; Addico, G.N.D.; Banu, R.A.; et al. Antibiotic-resistant bacteria in drinking water from the greater Accra region, Ghana: A cross-sectional study, December 2021–March 2022. Int. J. Environ. Res. Public Health 2022, 19, 12300. [Google Scholar] [CrossRef]
- Zhang, K.; Fan, Y.; Chang, S.; Fu, Q.; Zhang, Q.; Yang, G.; Sun, X. Characterization of antibiotic resistance genes in drinking water sources of the Douhe Reservoir, Tangshan, northern China: The correlation with bacterial communities and environmental factors. Environ. Sci. Eur. 2022, 34, 56. [Google Scholar] [CrossRef]
- Landrigan, P.J.; Stegeman, J.J.; Fleming, L.E.; Allemand, D.; Anderson, D.M.; Backer, L.C.; Brucker-Davis, F.; Chevalier, N.; Corra, L.; Czerucka, D.; et al. Human health and ocean pollution. Ann. Glob. Health 2020, 86, 151. [Google Scholar] [CrossRef]
- Fleming, L.E.; McDonough, N.; Austen, M.; Mee, L.; Moore, M.; Hess, P.; Depledge, M.H.; White, M.; Philippart, K.; Bradbrook, P.; et al. Oceans and human health: A rising tide of challenges and opportunities for Europe. Mar. Environ. Res. 2014, 99, 16–19. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Exp. Suppl. 2012, 101, 133–164. [Google Scholar] [PubMed]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic pollution in the environment: From microbial ecology to public policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sreekrishnan, T.R.; Ahammad, S.Z. Effects of heavy metals on the development and proliferation of antibiotic resistance in urban sewage treatment plants. Environ. Pollut. 2022, 308, 119649. [Google Scholar] [CrossRef]
- Tuvo, B.; Scarpaci, M.; Bracaloni, S.; Esposito, E.; Costa, A.L.; Ioppolo, M.; Casini, B. Microplastics and antibiotic resistance: The magnitude of the problem and the emerging role of hospital wastewater. Int. J. Environ. Res. Public Health 2023, 20, 5868. [Google Scholar] [CrossRef]
- Bong, C.W.; Low, K.Y.; Chai, L.C.; Lee, C.W. Prevalence and diversity of antibiotic resistant Escherichia coli from anthropogenic-impacted Larut river. Front. Public Health 2022, 10, 794513. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial resistance: A growing serious threat for global public health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef]
- Michaelis, C.; Grohmann, E. Horizontal gene transfer of antibiotic resistance genes in biofilms. Antibiotics 2023, 12, 328. [Google Scholar] [CrossRef]
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic resistance: One health one world outlook. Front. Cell. Infect. Microbiol. 2021, 11, 771510. [Google Scholar] [CrossRef]
- Monahan, C.; Nag, R.; Morris, D.; Cummins, E. Antibiotic residues in the aquatic environment—Current perspective and risk considerations. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2021, 56, 733–751. [Google Scholar] [CrossRef]
- Iwu, C.D.; Korsten, L.; Okoh, A.I. The incidence of antibiotic resistance within and beyond the agricultural ecosystem: A concern for public health. Microbiologyopen 2020, 9, e1035. [Google Scholar] [CrossRef] [PubMed]
- Pepi, M.; Focardi, S. Antibiotic-resistant bacteria in aquaculture and climate change: A challenge for health in the Mediterranean area. Int. J. Environ. Res. Public Health 2021, 18, 5723. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Meza, M.E.; Galarde-López, M.; Carrillo-Quiróz, B.; Alpuche-Aranda, C.M. Antimicrobial resistance: One health approach. Vet. World 2022, 15, 743–749. [Google Scholar] [CrossRef]
- Cella, E.; Giovanetti, M.; Benedetti, F.; Scarpa, F.; Johnston, C.; Borsetti, A.; Ceccarelli, G.; Azarian, T.; Zella, D.; Ciccozzi, M. Joining forces against antibiotic resistance: The one health solution. Pathogens 2023, 12, 1074. [Google Scholar] [CrossRef]
- Philippot, L.; Griffiths, B.S.; Langenheder, S. Microbial community resilience across ecosystems and multiple disturbances. Microbiol. Mol. Biol. Rev. 2021, 85, e00026-20. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of antimicrobial resistance in the environment and its relevance to environmental regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic resistance in the food chain: A developing country-perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The use of antibiotics and antimicrobial resistance in veterinary medicine, a complex phenomenon: A narrative review. Antibiotics 2023, 12, 487. [Google Scholar] [CrossRef]
- Heuer, O.E.; Kruse, H.; Grave, K.; Collignon, P.; Karunasagar, I.; Angulo, F.J. Human health consequences of use of antimicrobial agents in aquaculture. Clin. Infect. Dis. 2009, 49, 1248–1253. [Google Scholar] [CrossRef]
- Olmo, R.; Wetzels, S.U.; Armanhi, J.S.L.; Arruda, P.; Berg, G.; Cernava, T.; Cotter, P.D.; Araujo, S.C.; de Souza, R.S.C.; Ferrocino, I.; et al. Microbiome research as an effective driver of success stories in agrifood systems—A selection of Ccse studies. Front. Microbiol. 2022, 13, 834622. [Google Scholar] [CrossRef]
- Dadgostar, P. Antimicrobial resistance: Implications and costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed]
- Aminov, R.I. The role of antibiotics and antibiotic resistance in nature. Environ. Microbiol. 2009, 11, 2970–2988. [Google Scholar] [CrossRef] [PubMed]
- Samrot, A.V.; Wilson, S.; Preeth, R.S.S.; Prakash, P.; Sathiyasree, M.; Saigeetha, S.; Shobana, N.; Pachiyappan, S.; Rajesh, V.V. Sources of antibiotic contamination in wastewater and approaches to their removal—An overview. Sustainability 2023, 15, 12639. [Google Scholar] [CrossRef]
- Wang, T.; Luo, Y.; Kong, X.; Yu, B.; Zheng, P.; Huang, Z.; Mao, X.; Yu, J.; Luo, J.; Yan, H.; et al. Genetic- and fiber-diet-mediated changes in antibiotic resistance genes in pig colon contents and feces and their driving factors. Microorganisms 2023, 11, 2370. [Google Scholar] [CrossRef] [PubMed]
- Zarei-Baygi, A.; Smith, A.L. Intracellular versus extracellular antibiotic resistance genes in the environment: Prevalence, horizontal transfer, and mitigation strategies. Bioresour. Technol. 2021, 319, 124181. [Google Scholar] [CrossRef] [PubMed]
- Laht, M.; Karkman, A.; Voolaid, V.; Ritz, C.; Tenson, T.; Virta, M.; Kisand, V. Abundances of tetracycline, sulphonamide and beta-lactam antibiotic resistance genes in conventional wastewater treatment plants (WWTPs) with different waste load. PLoS ONE 2014, 9, e103705. [Google Scholar] [CrossRef] [PubMed]
- Warjri, I.; Dutta, T.K.; Lalzampuia, H.; Chandra, R. Detection and characterization of extended-spectrum β-lactamases (blaCTX-M-1 and blaSHV ) producing Escherichia coli, Salmonella spp. and Klebsiella pneumoniae isolated from humans in Mizoram. Vet. World 2015, 8, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic resistance genes in bacteria: Occurrence, spread, and control. J. Basic Microbiol. 2021, 61, 1049–1070. [Google Scholar] [CrossRef]
- Singh, N.S.; Singhal, N.; Virdi, J.S. Genetic environment of blaTEM-1, blaCTX-M-15, blaCMY-42 and characterization of integrons of Escherichia coli isolated from an Indian urban aquatic environment. Front. Microbiol. 2018, 9, 382. [Google Scholar] [CrossRef]
- Tacão, M.; Laço, J.; Teixeira, P.; Henriques, I. CTX-M-producing bacteria isolated from a highly polluted river system in Portugal. Int. J. Environ. Res. Public Health 2022, 19, 11858. [Google Scholar] [CrossRef]
- Mahdi, Z.M.; Hatif, Z.H.; Younis, M.H.; Abed Allah, R.A.A. Detection of resistance genes (Blandm and Blavim) of Escherichia coli in the aquatic environment. Iraqi J. Sci. 2023, 64, 6193–6203. [Google Scholar] [CrossRef]
- Gholami-Ahangaran, M.; Karimi-Dehkordi, M.; Miranzadeh-Mahabadi, E.; Ahmadi-Dastgerdi, A. The frequency of tetracycline resistance genes in Escherichia coli strains isolated from healthy and diarrheic pet birds. Iran. J. Vet. Res. 2021, 22, 337–341. [Google Scholar] [PubMed]
- Jahantigh, M.; Samadi, K.; Dizaji, R.E.; Salari, S. Antimicrobial resistance and prevalence of tetracycline resistance genes in Escherichia coli isolated from lesions of colibacillosis in broiler chickens in Sistan, Iran. BMC Vet. Res. 2020, 16, 267. [Google Scholar] [CrossRef]
- Grossman, T.H. Tetracycline antibiotics and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025387. [Google Scholar] [CrossRef] [PubMed]
- Zalewska, M.; Błażejewska, A.; Czapko, A.; Popowska, M. Antibiotics and antibiotic resistance genes in animal manure—Consequences of its application in agriculture. Front. Microbiol. 2021, 12, 610656. [Google Scholar] [CrossRef]
- Pazra, D.F.; Latif, H.; Basri, C.; Wibawan, I.W.T.; Rahayu, P. Distribution analysis of tetracycline resistance genes in Escherichia coli isolated from floor surface and effluent of pig slaughterhouses in Banten Province, Indonesia. Vet. World 2021, 16, 509–517. [Google Scholar] [CrossRef]
- Lin, X.; Ruan, J.; Huang, L.; Zhao, J.; Xu, Y. Comparison of the elimination effectiveness of tetracycline and AmpC β-lactamase resistance genes in a municipal wastewater treatment plant using four parallel processes. Ecotoxicology 2021, 30, 1586–1597. [Google Scholar] [CrossRef]
- Trif, E.; Cerbu, C.; Olah, D.; Zăblău, S.D.; Spînu, M.; Potârniche, A.V.; Pall, E.; Brudașcă, F. Old antibiotics can learn new ways: A systematic review of florfenicol use in veterinary medicine and future perspectives using nanotechnology. Animals 2023, 13, 1695. [Google Scholar] [CrossRef]
- Li, P.; Zhu, T.; Zhou, D.; Lu, W.; Liu, H.; Sun, Z.; Ying, J.; Lu, J.; Lin, X.; Li, K.; et al. Analysis of resistance to florfenicol and the related mechanism of dissemination in different animal-derived bacteria. Front. Cell. Infect. Microbiol. 2020, 10, 369. [Google Scholar] [CrossRef] [PubMed]
- Adesoji, A.T.; Call, D.R. Molecular analysis of florfenicol-resistant bacteria isolated from drinking water distribution systems in Southwestern Nigeria. J. Glob. Antimicrob. Resist. 2020, 23, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Deplano, A.; Meghraoui, A.; Dodémont, M.; Heinrichs, A.; Denis, O.; Nonhoff, C.; Roisin, S. Bacteria from animals as a pool of antimicrobial resistance genes. Antibiotics 2017, 6, 12. [Google Scholar] [CrossRef]
- Antunes, P.; Machado, J.; Sousa, J.C.; Peixe, L. Dissemination of sulfonamide resistance genes (sul1, sul2, and sul3) in Portuguese Salmonella enterica strains and relation with integrons. Antimicrob. Agents. Chemother. 2005, 49, 836–839. [Google Scholar] [CrossRef]
- Haenelt, S.; Wang, G.; Kasmanas, J.C.; Musat, F.; Richnow, H.H.; da Rocha, U.N.; Müller, J.A.; Musat, N. The fate of sulfonamide resistance genes and anthropogenic pollution marker intI1 after discharge of wastewater into a pristine river stream. Front. Microbiol. 2023, 14, 1058350. [Google Scholar]
- Jiang, H.; Cheng, H.; Liang, Y.; Yu, S.; Yu, T.; Fang, J.; Zhu, C. Diverse mobile genetic elements and conjugal transferability of sulfonamide resistance genes (sul1, sul2, and sul3) in Escherichia coli isolates From Penaeus vannamei and pork from large markets in Zhejiang, China. Front. Microbiol. 2019, 10, 1787. [Google Scholar] [CrossRef] [PubMed]
- Na, G.; Zhang, W.; Zhou, S.; Gao, H.; Lu, Z.; Wu, X.; Li, R.; Qiu, L.; Cai, Y.; Yao, Z. Sulfonamide antibiotics in the Northern Yellow Sea are related to resistant bacteria: Implications for antibiotic resistance genes. Mar. Pollut. Bull. 2014, 84, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.A.O.; Pavelquesi, S.L.S.; Monteiro, E.D.S.; Rodrigues, L.F.S.; Silva, C.M.S.; Silva, I.C.R.D.; Orsi, D.C. Prevalence and antimicrobial resistance of Salmonella spp. in aquacultured nile tilapia (Oreochromis niloticus) commercialized in federal district, Brazil. Foodborne. Pathog. Dis. 2021, 18, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.Y.; Araújo, S.; Folador, A.R.C.; Ramos, R.T.J.; Azevedo, J.S.N.; Tacão, M.; Silva, A.; Henriques, I.; Baraúna, R.A. Extended spectrum beta-lactamase-producing gram-negative bacteria recovered from an Amazonian Lake near the city of Belém, Brazil. Front. Microbiol. 2019, 10, 364. [Google Scholar] [CrossRef] [PubMed]
- Hassen, B.; Jouini, A.; Elbour, M.; Hamrouni, S.; Maaroufi, A. Detection of extended-spectrum β-lactamases (ESBL) producing Enterobacteriaceae from fish trapped in the Lagoon area of Bizerte, Tunisia. Biomed Res. Int. 2020, 2020, 7132812. [Google Scholar] [CrossRef]
- Ñahui Palomino, R.A.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota-host communications: Bacterial extracellular vesicles as a common language. PLoS Pathog. 2021, 17, e1009508. [Google Scholar] [CrossRef]
- Ghadami, S.; Dellinger, K. The lipid composition of extracellular vesicles: Applications in diagnostics and therapeutic delivery. Front. Mol. Biosci. 2023, 10, 1198044. [Google Scholar] [CrossRef]
- Briaud, P.; Carroll, R.K. Extracellular vesicle biogenesis and functions in gram-positive bacteria. Infect. Immun. 2020, 88, e00433-20. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, Z.; Liu, X.; Tyler, B.M. Biogenesis and biological functions of extracellular vesicles in cellular and organismal communication with microbes. Front. Mcrobiol. 2022, 13, 817844. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.H.K.; Aarestrup, F.M.; Petersen, T.N. Importance of mobile genetic elements for dissemination of antimicrobial resistance in metagenomic sewage samples across the world. PLoS ONE 2023, 18, e0293169. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Giv, N.; Basas, A.; Hicks, C.; El-Omar, E.; El-Assaad, F.; Hosseini-Beheshti, E. Bacterial extracellular vesicles and their novel therapeutic applications in health and cancer. Front. Cell. Infect. Microbiol. 2022, 12, 962216. [Google Scholar] [CrossRef] [PubMed]
- Biller, S.J.; Lundeen, R.A.; Hmelo, L.R.; Becker, K.W.; Arellano, A.A.; Dooley, K.; Heal, K.R.; Carlson, L.T.; Van Mooy, B.A.S.; Ingalls, A.E.; et al. Prochlorococcus extracellular vesicles: Molecular composition and adsorption to diverse microbes. Environ. Microbiol. 2022, 24, 420–435. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, N.E.; LeCleir, G.R.; Strzepek, R.F.; Ellwood, M.J.; Twining, B.S.; Roux, S.; Pennacchio, C.; Boyd, P.W.; Wilhelm, S.W. Bioavailable iron titrations reveal oceanic Synechococcus ecotypes optimized for different iron availabilities. ISME Commun. 2022, 2, 54. [Google Scholar] [CrossRef]
- Srivatsav, A.T.; Kapoor, S. The emerging world of membrane vesicles: Functional relevance, theranostic avenues and tools for investigating membrane function. Front. Mol. Biosci. 2021, 8, 640355. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef]
- Jan, A.T. Outer membrane vesicles (OMVs) of gram-negative bacteria: A perspective update. Front. Microbiol. 2021, 8, 1053. [Google Scholar] [CrossRef]
- Pérez-Cruz, C.; Delgado, L.; López-Iglesias, C.; Mercade, E. Outer-inner membrane vesicles naturally secreted by gram-negative pathogenic bacteria. PLoS ONE 2015, 10, e0116896. [Google Scholar] [CrossRef]
- Vitse, J.; Devreese, B. The contrhibution of membrane vesicles to bacterial pathogenicity in cystic fibrosis infections and healthcare associated pneumonia. Front. Microbiol. 2020, 11, 630. [Google Scholar] [CrossRef]
- Coelho, C.; Casadevall, A. Answers to naysayers regarding microbial extracellular vesicles. Biochem Soc Trans. 2019, 47, 1005–1012. [Google Scholar] [CrossRef]
- Pospíšil, J.; Vítovská, D.; Kofroňová, O.; Muchová, K.; Šanderová, H.; Hubálek, M.; Šiková, M.; Modrák, M.; Benada, O.; Barák, I.; et al. Bacterial nanotubes as a manifestation of cell death. Nat. Commun. 2020, 11, 4963. [Google Scholar] [CrossRef]
- Tian, C.M.; Yang, M.F.; Xu, H.M.; Zhu, M.Z.; Zhang, Y.; Yao, J.; Wang, L.S.; Liang, Y.J.; Li, D.F. Emerging role of bacterial outer membrane vesicle in gastrointestinal tract. Gut Pathog. 2023, 15, 20. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Sondak, T.; Kim, K.S. Characterization of increased extracellular vesicle-mediated tigecycline resistance in Acinetobacter baumannii. Pharmaceutics 2023, 15, 1251. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.T.; Kuo, S.C.; Chiang, M.H.; Lee, Y.T.; Sung, W.C.; Chen, Y.H.; Chen, T.L.; Fung, C.P. Acinetobacter baumannii extracellular OXA-58 is primarily and selectively released via outer membrane vesicles after sec-dependent periplasmic translocation. Antimicrob. Agents Chemother. 2015, 59, 7346–7354. [Google Scholar] [CrossRef]
- Collins, S.M.; Brown, A.C. Bacterial outer membrane vesicles as antibiotic delivery vehicles. Front. Immunol. 2021, 12, 733064. [Google Scholar] [CrossRef]
- Kim, S.W.; Park, S.B.; Im, S.P.; Lee, J.S.; Jung, J.W.; Gong, T.W.; Lazarte, J.M.S.; Kim, J.; Seo, J.S.; Kim, J.H.; et al. Outer membrane vesicles from β-lactam-resistant Escherichia coli enable the survival of β-lactam-susceptible E. coli in the presence of β-lactam antibiotics. Sci. Rep. 2018, 8, 5402. [Google Scholar] [CrossRef]
- Mao, H.; Guo, J.; Zhou, J.; Shi, J.; Cui, H.; Shi, R.; Yao, J.; Fang, X.; Wang, B.; Yan, F. Antimicrobial poly(ionic liquid)-induced bacterial nanotube formation and drug-resistance spread. Biomater. Sci. 2022, 10, 6460–6471. [Google Scholar] [CrossRef]
- Lucena, A.C.R.; Ferrarini, M.G.; de Oliveira, W.K.; Marcon, B.H.; Morello, L.G.; Alves, L.R.; Faoro, H. Modulation of Klebsiella pneumoniae outer membrane vesicle protein cargo under antibiotic treatment. Biomedicines. 2023, 11, 1515. [Google Scholar] [CrossRef]
- Tikhomirova, A.; Kidd, S.P. Haemophilus influenzae and Streptococcus pneumoniae: Living together in a biofilm. Pathog. Dis. 2013, 69, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Abebe, G.M. The role of bacterial biofilm in antibiotic resistance and food contamination. Int. J. Microbiol. 2020, 2020, 1705814. [Google Scholar] [CrossRef]
- Zaborowska, M.; Taulé Flores, C.; Vazirisani, F.; Shah, F.A.; Thomsen, P.; Trobos, M. Extracellular vesicles influence the growth and adhesion of Staphylococcus epidermidis under antimicrobial selective pressure. Front. Microbiol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Diggle, S.P.; Matthijs, S.; Wright, V.J.; Fletcher, M.P.; Chhabra, S.R.; Lamont, I.L.; Kong, X.; Hider, R.C.; Cornelis, P.; Cámara, M.; et al. The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem. Biol. 2007, 14, 87–96. [Google Scholar] [CrossRef]
- Wang, X.; Koffi, P.F.; English, O.F.; Lee, J.C. Staphylococcus aureus extracellular vesicles: A story of toxicity and the stress of 2020. Toxins 2021, 13, 75. [Google Scholar] [CrossRef]
- Parveen, S.; Subramanian, K. Emerging roles of extracellular vesicles in pneumococcal infections: Immunomodulators to potential novel vaccine candidates. Front. Cell. Infect. Microbiol 2022, 12, 836070. [Google Scholar] [CrossRef]
- Rivera, J.; Cordero, R.J.; Nakouzi, A.S.; Frases, S.; Nicola, A.; Casadevall, A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 19002–19007. [Google Scholar] [CrossRef]
- Liu, S.; Wu, X.; Chandra, S.; Lyon, C.; Ning, B.; Jiang, L.; Fan, J.; Hu, T.Y. Extracellular vesicles: Emerging tools as therapeutic agent carriers. Acta. Pharmaceutica. Sinica. B 2022, 12, 3822–3842. [Google Scholar] [CrossRef]
- Mosquera-Heredia, M.I.; Morales, L.C.; Vidal, O.M.; Barceló, E.; Silvera-Redondo, C.; Vélez, J.I.; Garavito-Galofre, P. Exosomes: Potential disease biomarkers and new therapeutic targets. Biomedicines 2021, 9, 1061. [Google Scholar] [CrossRef]
- Huda, M.N.; Nafiujjaman, M.; Deaguero, I.G.; Okonkwo, J.; Hill, M.L.; Kim, T.; Nurunnabi, M. Potential use of exosomes as diagnostic biomarkers and in targeted drug delivery: Progress in clinical and preclinical applications. ACS Biomater. Sci. Eng. 2021, 7, 2106–2149. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Zimmermann, A.K.; Rivieccio, F.; Visser, C.; Blango, M.G. Host-derived extracellular vesicles for antimicrobial defense. Microlife 2021, 2, uqab003. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles. 2015, 4, 27066. [Google Scholar] [CrossRef]
- Makarova, M.; May, R.C. Fungal extracellular vesicles in interkingdom communication. Curr. Top. Microbiol. Immunol. 2021, 432, 81–88. [Google Scholar]
- Amillano-Cisneros, J.M.; Fuentes-Valencia, M.A.; Leyva-Morales, J.B.; Davizón, Y.A.; Marquéz-Pacheco, H.; Valencia-Castañeda, G.; Maldonado-Coyac, J.A.; Ontiveros-García, L.A.; Badilla-Medina, C.N. Prebiotics in global and Mexican fish aquaculture: A review. Animals 2023, 13, 3607. [Google Scholar] [CrossRef] [PubMed]
- Hadizadeh, N.; Bagheri, D.; Shamsara, M.; Hamblin, M.R.; Farmany, A.; Xu, M.; Liang, Z.; Razi, F.; Hashemi, E. Extracellular vesicles biogenesis, isolation, manipulation and genetic engineering for potential in vitro and in vivo therapeutics: An overview. Front. Bioeng. Biotechnol. 2022, 10, 1019821. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, M.L.; Ghosh, S.; Chen, Z. Phytoremediation: A promising approach for revegetation of heavy metal-polluted land. Front. Plant. Sci. 2020, 11, 359. [Google Scholar] [CrossRef]
- Okoye, C.O.; Nyaruaba, R.; Ita, R.E.; Okon, S.U.; Addey, C.I.; Ebido, C.C.; Opabunmi, A.O.; Okeke, E.S.; Chukwudozie, K.I. Antibiotic resistance in the aquatic environment: Analytical techniques and interactive impact of emerging contaminants. Environ. Toxicol. Pharmacol. 2022, 96, 103995. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Beveridge, T.J. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: A novel mechanism of enzyme secretion. J. Bacteriol. 1995, 177, 3998–4008. [Google Scholar] [CrossRef]
- Huang, Y.; Nieh, M.P.; Chen, W.; Lei, Y. Outer membrane vesicles (OMVs) enabled bio-applications. Biotechnol. Bioeng. 2022, 119, 34–47. [Google Scholar] [CrossRef]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes. Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef]



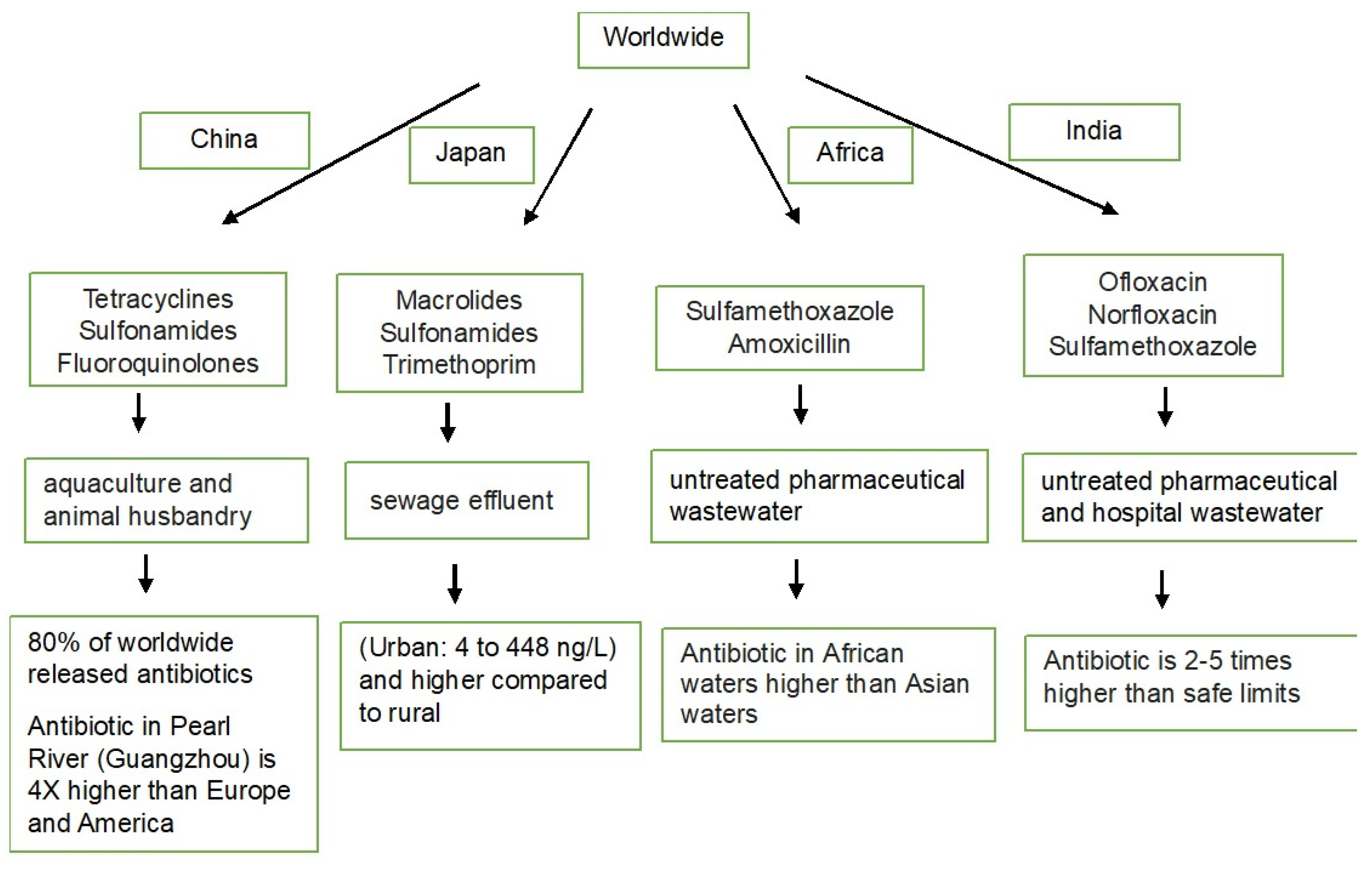{kind=link}
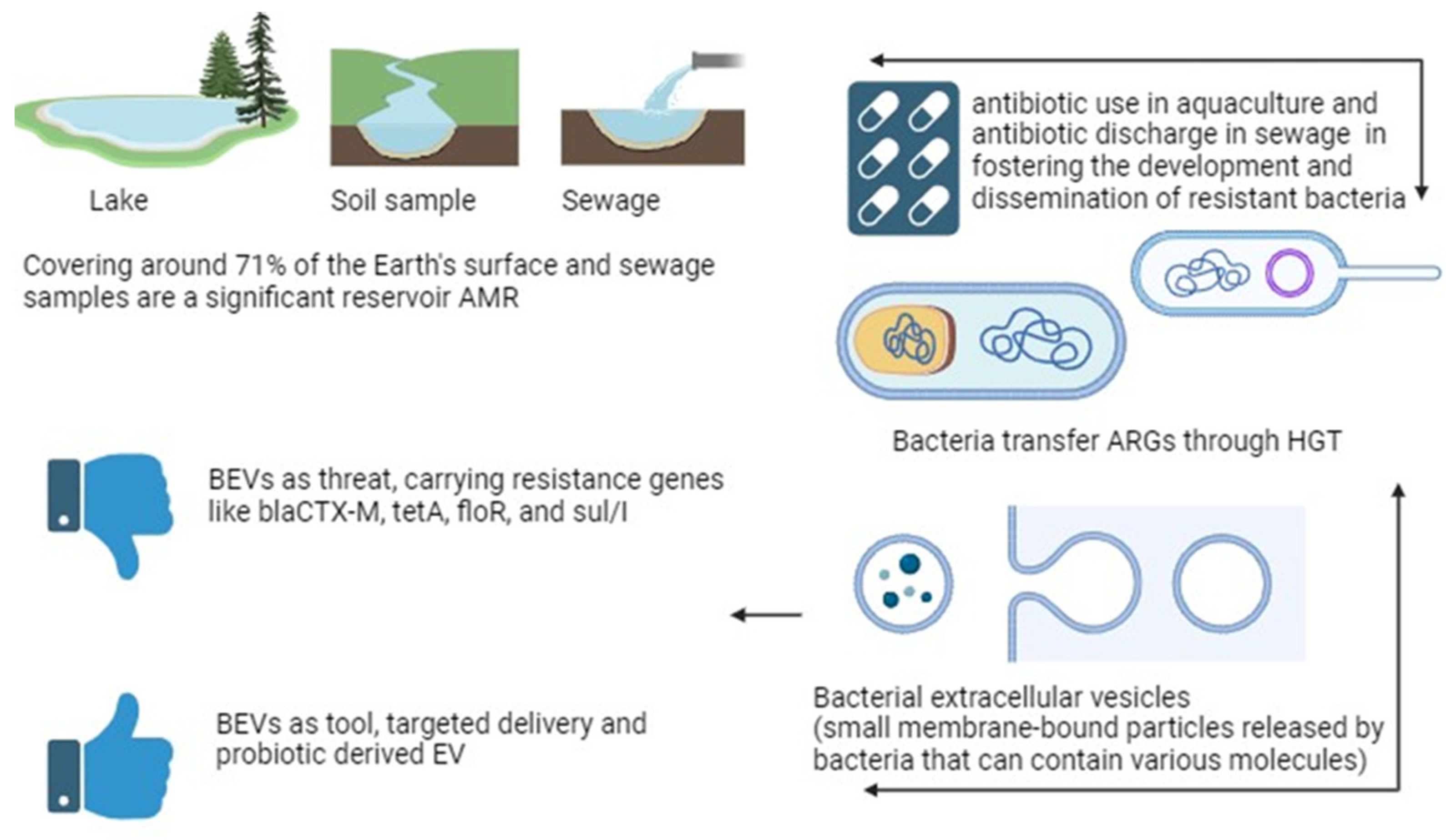{kind=link}
| ARGs | Antibiotic | Bacteria | Mechanism | Source | Citation |
|---|---|---|---|---|---|
| blaCTX-M | β-lactams (penicillins, cephalosporin, carbapenems) | Klebsiella pneumoniae, Escherichia coli, Salmonella spp. | Transfer through HGT (conjugation, transduction, transformation) | River, wastewater (United States) | [67,68,69,70] |
| blaTEM-1, blaCTX-M-15, blaCMY-42 | β-lactams | Escherichia coli | Plasmid-mediated HGT | River (India) | [71] |
| blaCTX-M | β-lactams | Enterobacteriaceae | Plasmid-mediated HGT | River (Portugal) | [72] |
| tetA, tetK, tetC, tetE, tetM | Tetracyclines | Escherichia coli | Efflux pumps and ribosomal protection proteins | Pig slaughterhouses (Indonesia); wastewater, agricultural runoff (Guangzhou) | [76,77] |
| floR | Florfenicol | Escherichia coli, Pseudomonas, Serratia, Proteus, Acinetobacter, Providencia rettgeri | Florfenicol efflux pump | Drinking water (Southwestern Nigeria) | [80,81] |
| sul/I | Sulfonamides | Escherichia coli | Plasmid and/or chromosome transfer through conjugation | Sea and shrimp (China) | [82,83] |
| blaCTX, tetA, floR, sul2 | Multiple | Salmonella spp., E. coli | Plasmid-mediated HGT | Fish (Kenya) | [84,85] |
| blaCTX–M, blaTEM, blaSHV | β-lactams | Extended-spectrum beta-lactamase Gram-negative bacteria | HGT | Lake (Brazil) | [86] |
| blaCTX-M-1, blaOXA-1, blaTEM-1-a, qnrA, qnrB | β-lactams and quinolone | Extended-spectrum beta-lactamase Gram-negative bacteria | Plasmid-mediated, efflux pump, and horizontal gene transfer | Lagoon (Tunisia) | [87] |
| Mechanism | Bacteria Involved | Resistance Genes/Enzymes | Citation |
|---|---|---|---|
| BEVs carrying β-lactamase enzymes | Acinetobacter baumannii, Stenotrophomonas maltophilia, Escherichia coli, Moraxella catarrhalis, Bacteroides fragilis | OXA-58 (carbapenemase), β-lactamases | [105] |
| Bacterial nanotube formation induced by cationic poly(ionic liquid)-based materials | Escherichia coli, Staphylococcus aureus, Vibrio fischeri | ARGs (not specified) | [108] |
| OMVs carrying β-lactamase enzymes | Klebsiella pneumoniae HCD1, Haemophilus influenzae (potentially packaged by Streptococcus) | KPC-2 (carbapenemase), β-lactamase | [106,107,108,109,110] |
| EVs contributing to biofilm formation and structure | Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pneumoniae, Bacillus anthracis | PQS (signaling molecule), β-lactamases, superantigens, toxins, cytosolic pore-forming toxins, anthrax toxin components | [112,113,114,115,116] |
| Application | Description | Advantages | Challenges |
|---|---|---|---|
| BEVs as drug delivery vehicles | BEVs engineered to carry natural antimicrobial cargo or loaded with antimicrobial agents | Enhanced specificity and reduced side effects compared with conventional antibiotics | Lack of standardization |
| BEVs loaded with antibiotic-degrading enzymes or bacteria | Delivering BEVs loaded with enzymes or bacteria to plant roots for targeted antibiotic decontamination | Targeted approach to antibiotic removal | Ensuring BEV stability and efficient enzyme/bacteria activity |
| Plant EVs for antibiotic removal | Engineering plant EVs to specifically bind and internalize antibiotic molecules for improved removal efficiency | Environmentally friendly approach to antibiotic removal in water and soil | Understanding and manipulating plant EV–antibiotic interactions and ambiguous mechanisms |
| BEVs with probiotics | BEVs derived from beneficial bacteria that produce prebiotics or probiotics introduced into aquatic environments | Low immunogenicity and -probiotics can inhibit pathogens through various mechanisms | Ensuring BEV stability and viability in the environment |
| Modified host BEVs | Engineering EVs with specific ligands or surface proteins for targeted delivery to pathogens | Potential for developing new therapies against challenging-to-treat pathogens | Addressing potential safety concerns of modified EVs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barathan, M.; Ng, S.-L.; Lokanathan, Y.; Ng, M.H.; Law, J.X. Unseen Weapons: Bacterial Extracellular Vesicles and the Spread of Antibiotic Resistance in Aquatic Environments. Int. J. Mol. Sci. 2024, 25, 3080. https://doi.org/10.3390/ijms25063080
Barathan M, Ng S-L, Lokanathan Y, Ng MH, Law JX. Unseen Weapons: Bacterial Extracellular Vesicles and the Spread of Antibiotic Resistance in Aquatic Environments. International Journal of Molecular Sciences. 2024; 25(6):3080. https://doi.org/10.3390/ijms25063080
Chicago/Turabian StyleBarathan, Muttiah, Sook-Luan Ng, Yogeswaran Lokanathan, Min Hwei Ng, and Jia Xian Law. 2024. "Unseen Weapons: Bacterial Extracellular Vesicles and the Spread of Antibiotic Resistance in Aquatic Environments" International Journal of Molecular Sciences 25, no. 6: 3080. https://doi.org/10.3390/ijms25063080