
Picea wilsonii NAC31 and DREB2A Cooperatively Activate ERD1 to Modulate Drought Resistance in Transgenic Arabidopsis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
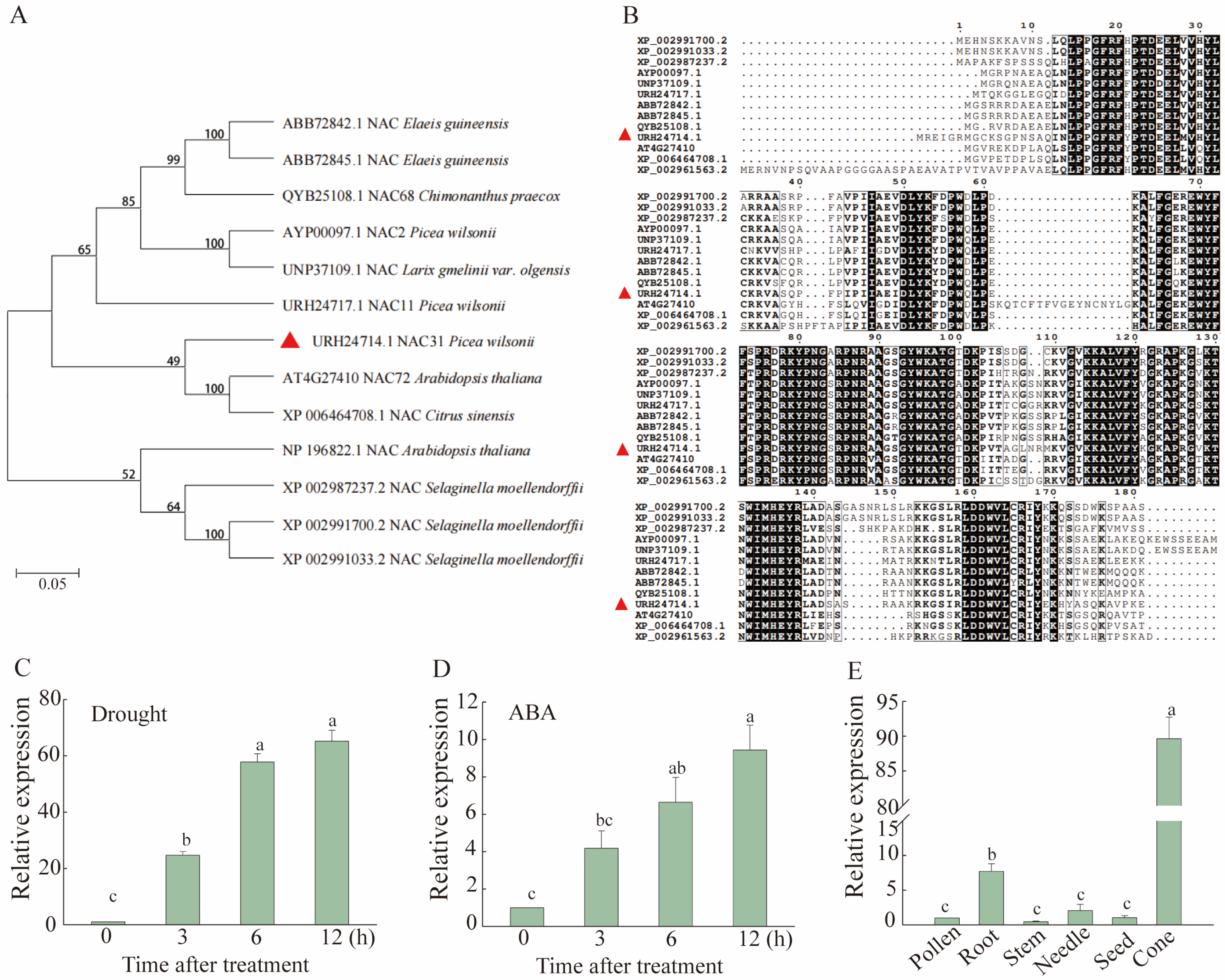
2.1. Identification and Analysis of PwNAC31
2.2. PwNAC31 Responds to Drought Stress and ABA
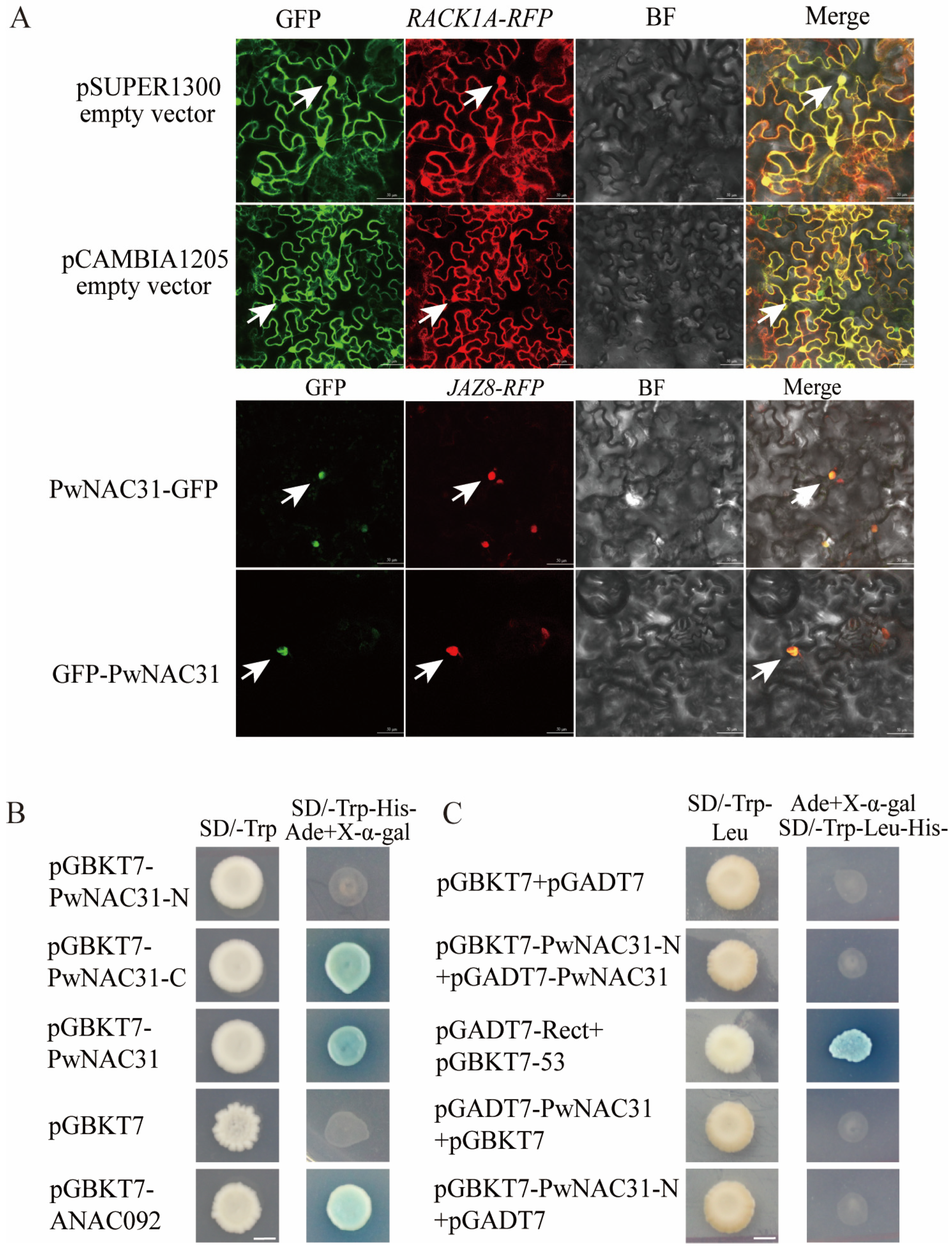
2.3. The Subcellular Localization of PwNAC31 and Activity of Transcriptional Activation
2.4. PwNAC31 Enhances Seed Vigor and Germination under Drought Stress
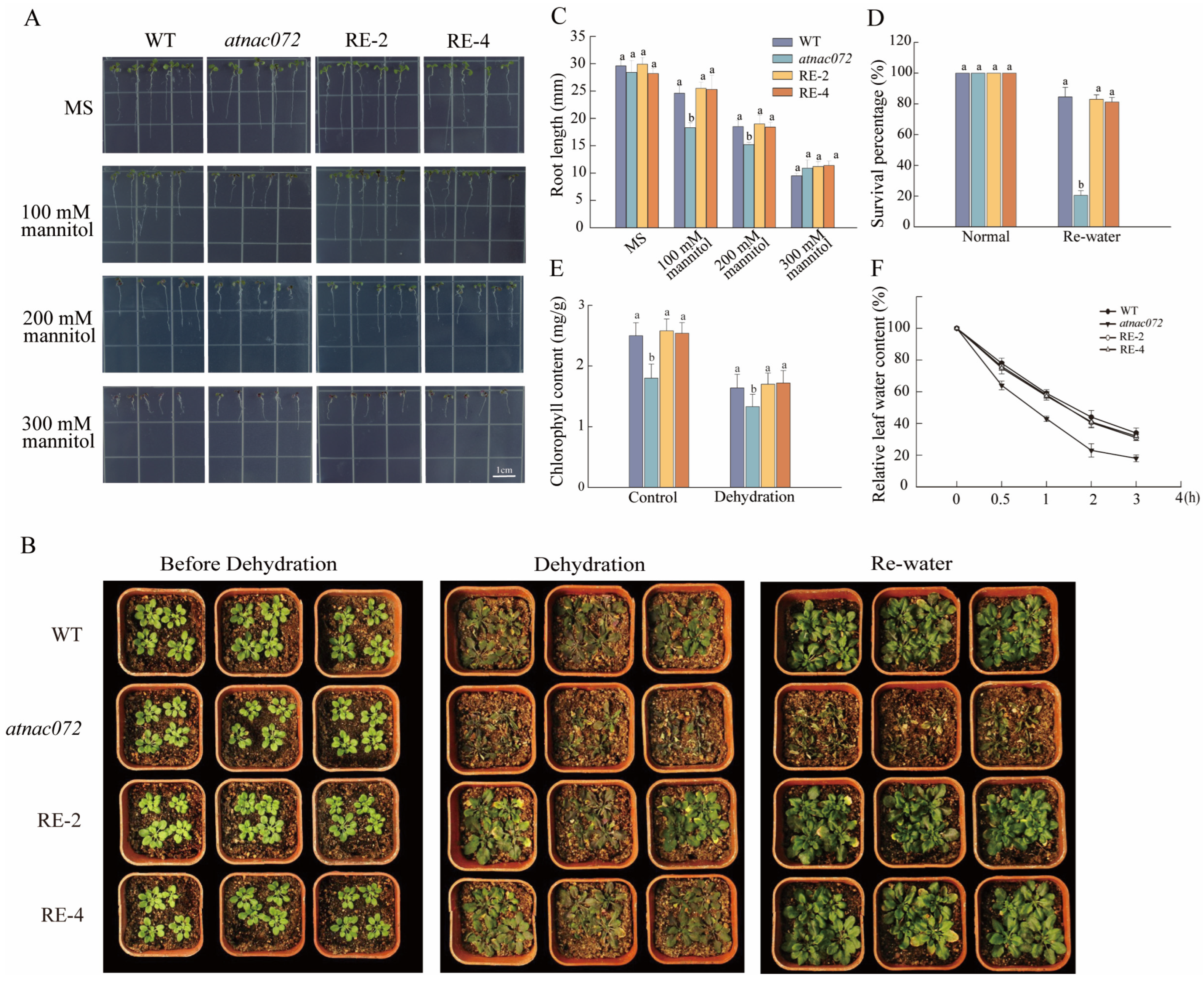
2.5. PwNAC31 Enhances Drought Tolerance in Transgenic Arabidopsis
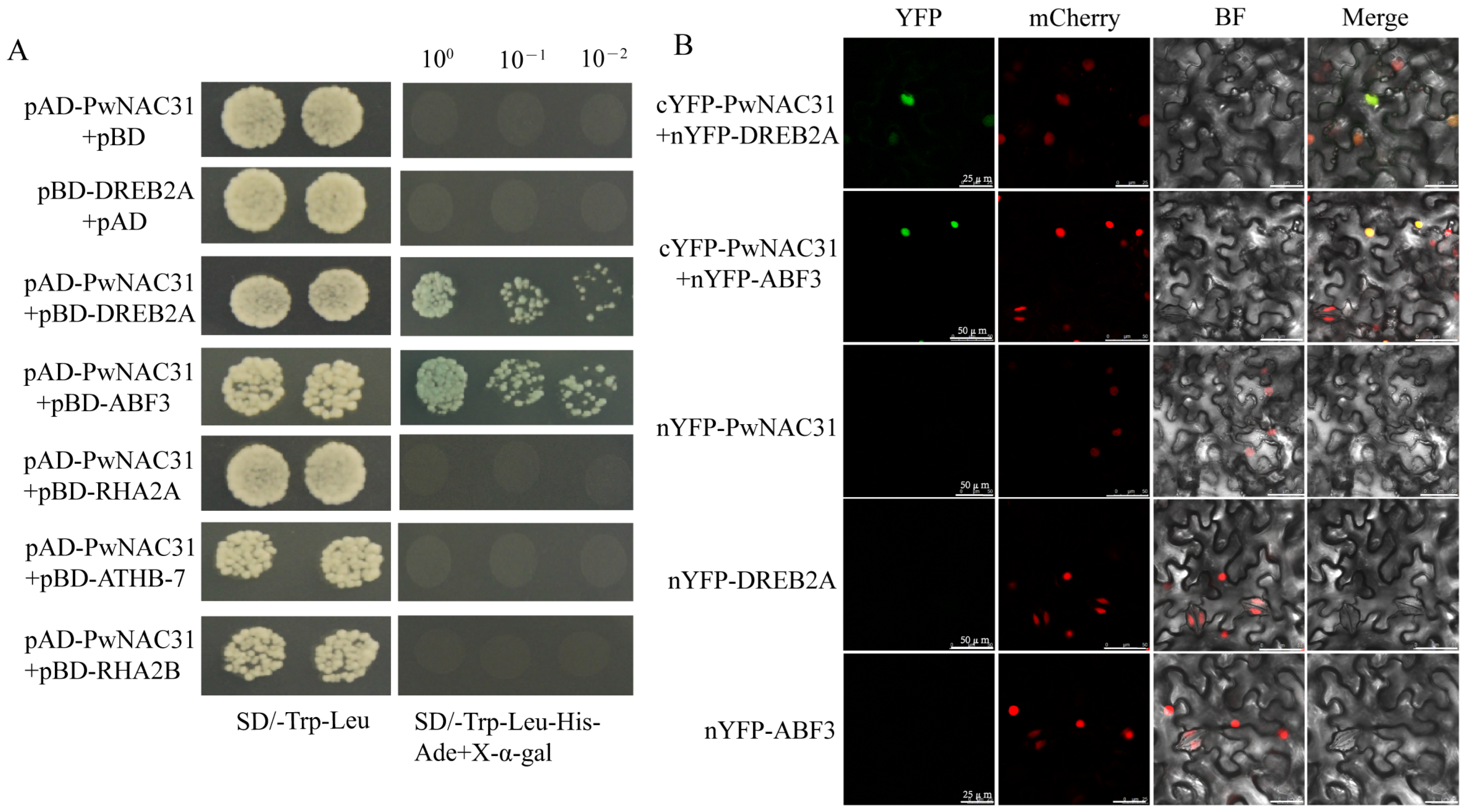
2.6. PwNAC31 Interacts with ABF3 and DREB2A
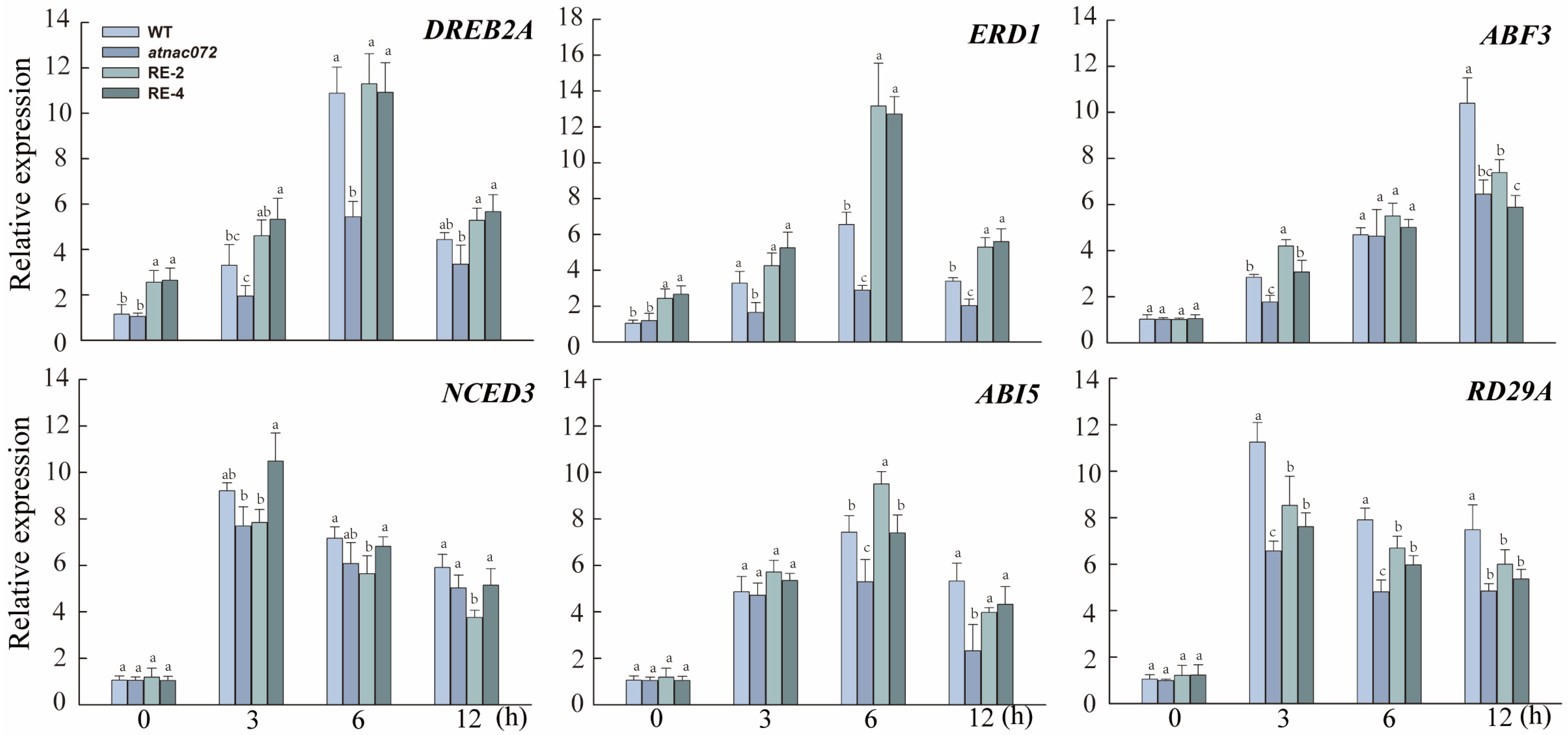
2.7. Expression Pattern Analysis of Genes Responding to Drought and ABA in PwNAC31 Transgenic Arabidopsis
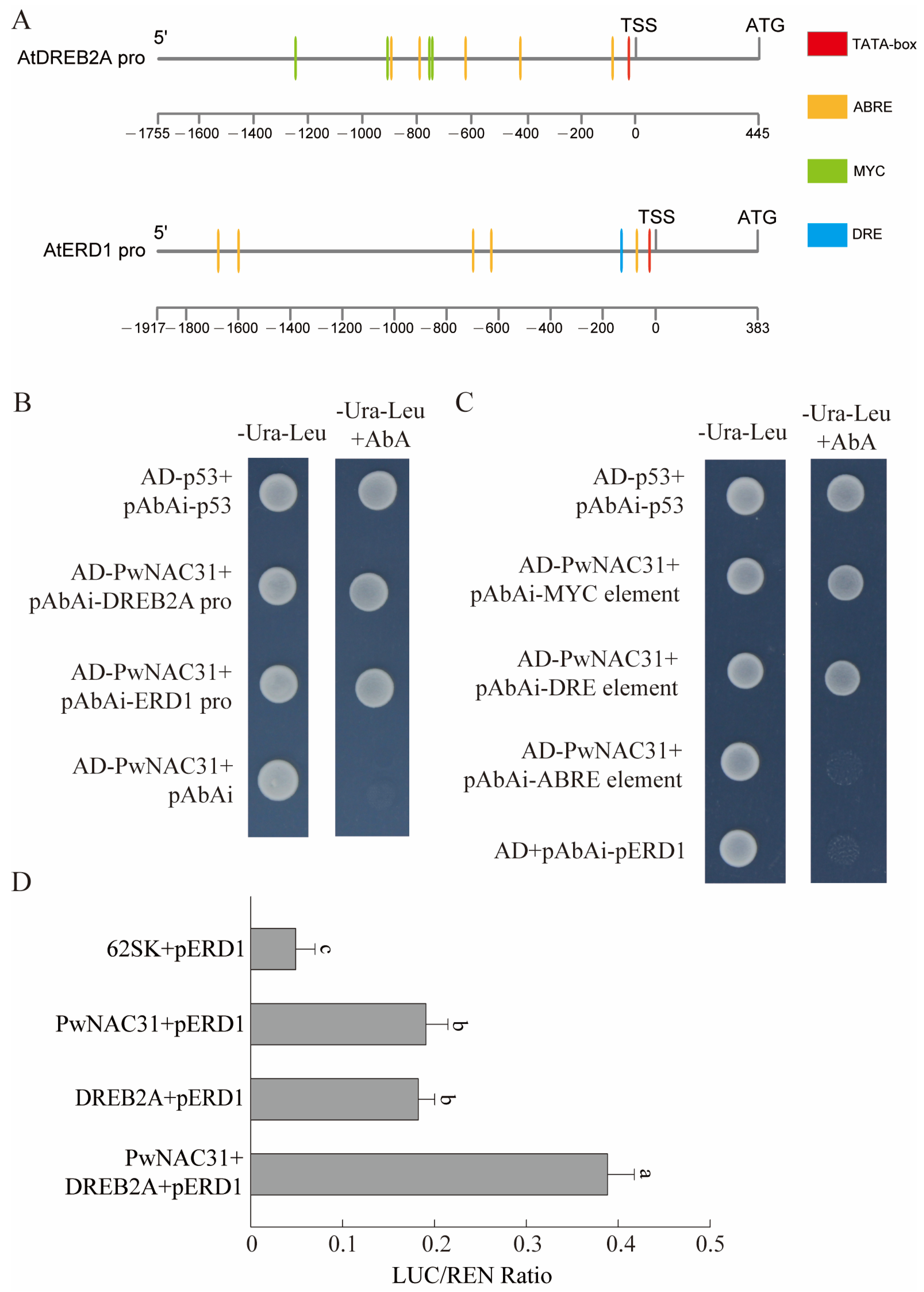
2.8. PwNAC31 Can Combine with the Promoter Region of DREB2A and ERD1
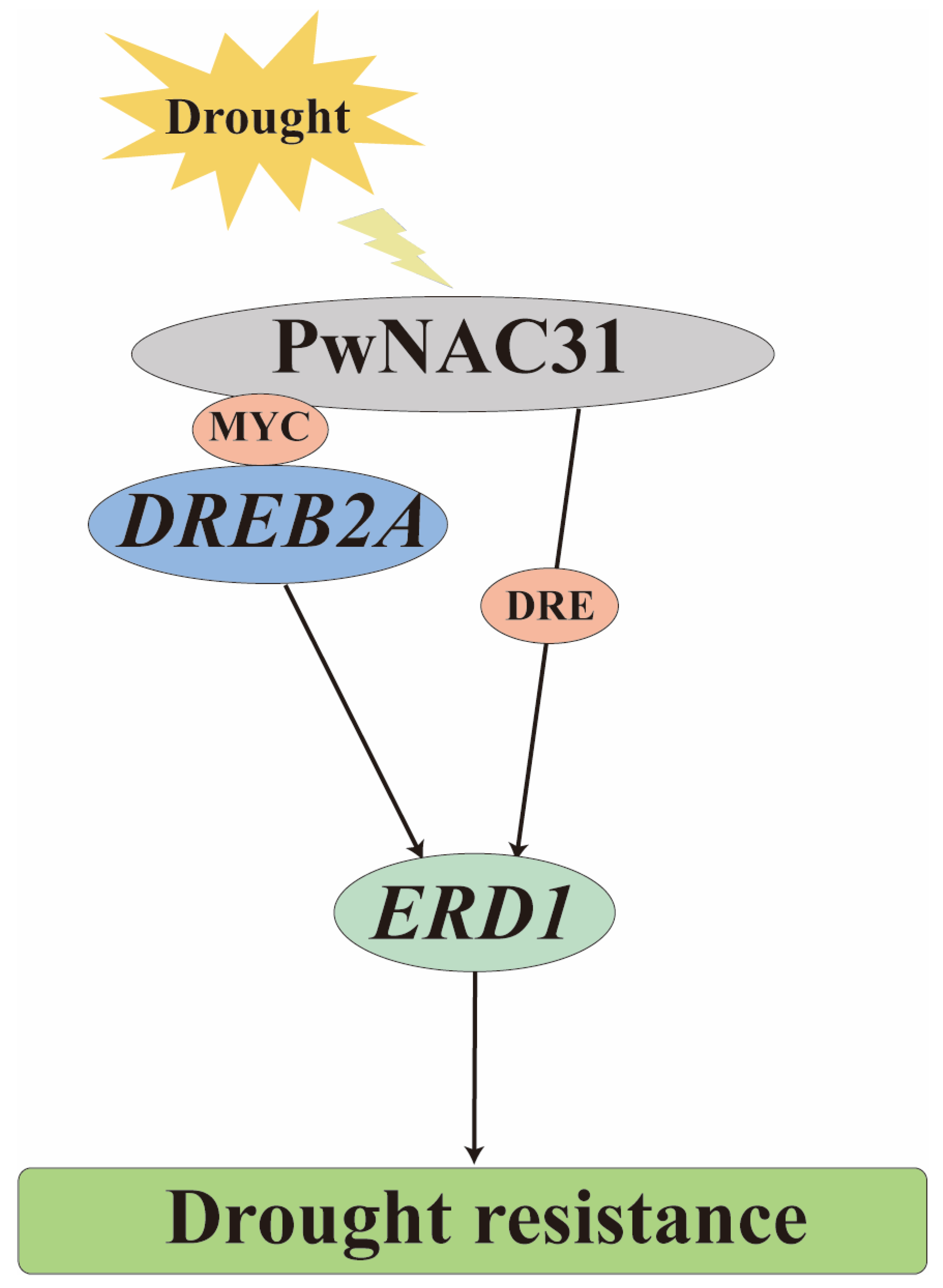
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Drought Treatments
4.2. Gene Isolation and Homologous Analysis
4.3. RNA Isolation and Quantitative Real-Time PCR
4.4. Subcellular Localization
4.5. Transcriptional Activity Analysis and the Verification of Interaction Proteins via Y2H Assay
4.6. The Verification of Interaction Proteins by BiFC (Bimolecular Fluorescence Complementation)
4.7. Yeast One-Hybrid (Y1H) Assays
4.8. Dual-Luciferase Assay
4.9. Physiological Index Measurement
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rezende, M.; Sanchez-Paus Diaz, A.; Bastin, J.-F.; Mollicone, D. Trees, Forests and Land Use in Drylands: The First Global Assessment; FAO: Roma, Italy, 2016. [Google Scholar]
- Pangle, R.E.; Limousin, J.M.; Plaut, J.A.; Yepez, E.A.; Hudson, P.J.; Boutz, A.L.; Gehres, N.; Pockman, W.T.; McDowell, N.G. Prolonged experimental drought reduces plant hydraulic conductance and transpiration and increases mortality in a piñon-juniper woodland. Ecol. Evol. 2015, 5, 1618–1638. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, X.; Guo, Y.; Luo, C.; Zhang, L. Picea wilsonii transcription factor NAC2 enhanced plant tolerance to abiotic stress and participated in RFCP1-regulated flowering time. Plant Mol. Biol. 2018, 98, 471–493. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Yadav, D.; Khan, A.; Hashem, A.; Tabassum, B.; Khan, A.L.; Abd Allah, E.F.; Al-Harrasi, A. Genomics, molecular and evolutionary perspective of NAC transcription factors. PLoS ONE 2020, 15, e0231425. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.J.; Li, C.Z.; Lin, S.Y.; Wang, J.P.; Zhou, B.R.; Jiang, T.B. Transcriptome analysis of salt-responsive and wood-associated NACs in Populus simonii × Populus nigra. BMC Plant Biol. 2020, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wu, T.; Guo, S.; Hu, J.; Zhan, Y. Ectopic Expression of AeNAC83, a NAC Transcription Factor from Abelmoschus esculentus, Inhibits Growth and Confers Tolerance to Salt Stress in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 10182. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.Q.; Ye, Z.H. Secondary Cell Walls: Biosynthesis, Patterned Deposition and Transcriptional Regulation. Plant Cell Physiol. 2015, 56, 195–214. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liu, S.; Zhang, C.; He, J.; Ma, D.; Wang, X.; Dong, T.; Guo, F.; Cai, J.; Long, T.; et al. The unique sweet potato NAC transcription factor IbNAC3 modulates combined salt and drought stresses. Plant Physiol. 2023, 191, 747–771. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-L.; Wu, Q.; Wu, H.-L.; Wang, A.-H.; Wang, X.-L.; Li, C.-L.; Zhao, H.-X.; Wu, Q. FtNAC31, a Tartary buckwheat NAC transcription factor, enhances salt and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2022, 191, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Zeng, Z.; Lyu, Y.; Zhao, S. Drought-Responsive NAC Transcription Factor RcNAC72 Is Recognized by RcABF4, Interacts with RcDREB2A to Enhance Drought Tolerance in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1755. [Google Scholar] [CrossRef] [PubMed]
- Mijiti, M.; Wang, Y.; Wang, L.; Habuding, X. Tamarix hispida NAC Transcription Factor ThNAC4 Confers Salt and Drought Stress Tolerance to Transgenic Tamarix and Arabidopsis. Plants 2022, 11, 2647. [Google Scholar] [CrossRef]
- Li, J.; Guo, W.; Zhao, J.; Meng, H.; Yang, Y.; Zheng, G.; Yuan, W. Transcriptional Regulation of the Acer truncatum B. Response to Drought and the Contribution of AtruNAC36 to Drought Tolerance. Antioxidants 2023, 12, 1339. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Huang, Y.; Lv, P.; Antwi-Boasiako, A.; Begum, N.; Zhao, T.; Zhao, J. NAC Transcription Factor GmNAC12 Improved Drought Stress Tolerance in Soybean. Int. J. Mol. Sci. 2022, 23, 12029. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.-S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and Functional Analysis of Arabidopsis Stress-Inducible NAC Transcription Factors That Bind to a Drought-Responsive cis-Element in the early responsive to dehydration stress 1 Promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef]
- Zheng, Y.C.; Jin, X.Y.; Wang, J.Y.; Chen, W.; Yang, Z.; Chen, Y.X.; Yang, Y.H.; Lu, G.H.; Sun, B. SbNAC9 Improves Drought Tolerance by Enhancing Scavenging Ability of Reactive Oxygen Species and Activating Stress-Responsive Genes of Sorghum. Int. J. Mol. Sci. 2023, 24, 2401. [Google Scholar] [CrossRef]
- Xia, L.; Sun, S.; Han, B.; Yang, X. NAC domain transcription factor gene GhNAC3 confers drought tolerance in plants. Plant Physiol. Biochem. 2023, 195, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, Y.; Yu, W.; Wang, L.; Lan, Q.; Wang, Y.; Chen, C.; Zhang, Y. Knocking Out the Transcription Factor OsNAC092 Promoted Rice Drought Tolerance. Biology 2022, 11, 1830. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Jiang, Q.; van Nocker, S.; Gong, X.; Ma, F. An apple (Malus domestica) NAC transcription factor enhances drought tolerance in transgenic apple plants. Plant Physiol. Biochem. 2019, 139, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wang, P.; Chen, X.; Zhao, H.; Zhu, Q.; Song, Y.; Nie, Y.; Li, Y.; Guo, M.; Niu, S. An ethylene-induced NAC transcription factor acts as a multiple abiotic stress responsor in conifer. Hortic. Res. 2023, 10, uhad130. [Google Scholar] [CrossRef]
- Yu, M.; Liu, J.; Du, B.; Zhang, M.; Wang, A.; Zhang, L. NAC Transcription Factor PwNAC11 Activates ERD1 by Interaction with ABF3 and DREB2A to Enhance Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 6952. [Google Scholar] [CrossRef]
- He, L.; Shi, X.; Wang, Y.; Guo, Y.; Yang, K.; Wang, Y. Arabidopsis ANAC069 binds to C A/G CG T/G sequences to negatively regulate salt and osmotic stress tolerance. Plant Mol. Biol. 2017, 93, 369–387. [Google Scholar] [CrossRef]
- Liang, K.-H.; Wang, A.-B.; Yuan, Y.-H.; Miao, Y.-H.; Zhang, L.-Y. Picea wilsonii NAC Transcription Factor PwNAC30 Negatively Regulates Abiotic Stress Tolerance in Transgenic Arabidopsis. Plant Mol. Biol. Rep. 2020, 38, 554–571. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, Y.; Xie, Z.; Yu, B.; Sun, Y.; Huang, J. OsNAC016 regulates plant architecture and drought tolerance by interacting with the kinases GSK2 and SAPK8. Plant Physiol. 2022, 189, 1296–1313. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Kim, Y.-S.; Han, S.-H.; Lee, B.-D.; Paek, N.-C. The Arabidopsis Transcription Factor NAC016 Promotes Drought Stress Responses by Repressing AREB1 Transcription through a Trifurcate Feed-Forward Regulatory Loop Involving NAP. Plant Cell 2015, 27, 1771–1787. [Google Scholar] [CrossRef] [PubMed]
- Ma, H. Functional Traits of Picea wilsonii Based on Tree Age and Altitude. Master’s Thesis, Shanxi Agricultural University, Jinzhong, China, 2019. [Google Scholar]
- Zhang, T.; Li, Y.; Zhou, Y.; Zhang, L. Cloning and expression analysis of a homologous expansin gene EXP2 in Picea wilsonii. J. For. Res. 2016, 27, 247–255. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, H.; Yuan, Y.; Cui, X.; Zhang, L. Identification and characterization of NAC genes in response to abiotic stress conditions in Picea wilsonii using transcriptome sequencing. Biotechnol. Biotechnol. Equip. 2020, 34, 93–103. [Google Scholar] [CrossRef]
- Cui, X.Y.; Cao, Y.B.; Zhang, H.H.; Zhang, L.Y. A Picea wilsonii NAC transcription factor, PwNAC1, interacts with RNA-binding protein PwRBP1 and synergistically enhances drought and salt tolerance of transgenic Arabidopsis. Environ. Exp. Bot. 2023, 206, 105174. [Google Scholar] [CrossRef]
- Liu, J.; Qiao, Y.; Li, C.; Hou, B. The NAC transcription factors play core roles in flowering and ripening fundamental to fruit yield and quality. Front. Plant Sci. 2023, 14, 1095967. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Li, C.; Wang, F.; Zhang, F.; Liu, J.; Wang, J.; Zhang, X.; Kong, X.; Ding, Z. NAC1 regulates root ground tissue maturation by coordinating with the SCR/SHR-CYCD6;1 module in Arabidopsis. Mol. Plant 2023, 16, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Yu, G.; Lei, S.; Wang, H.; Xu, B. STRONG STAYGREEN inhibits DNA binding of PvNAP transcription factors during leaf senescence in switchgrass. Plant Physiol. 2022, 190, 2045–2058. [Google Scholar] [CrossRef]
- Dong, Y.; Tang, M.; Huang, Z.; Song, J.; Xu, J.; Ahammed, G.J.; Yu, J.; Zhou, Y. The miR164a-NAM3 module confers cold tolerance by inducing ethylene production in tomato. Plant J. 2022, 111, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Yue, X.; Zeng, H.; Zhu, J. The protein phosphatase RCF2 and its interacting partner NAC019 are critical for heat stress-responsive gene regulation and thermotolerance in Arabidopsis. Plant Cell 2014, 26, 438–453. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Willems, L.A.J.; Jiao, A.; Zhao, T.; Eric Schranz, M.; Bentsink, L. The membrane associated NAC transcription factors ANAC060 and ANAC040 are functionally redundant in the inhibition of seed dormancy in Arabidopsis thaliana. J. Exp. Bot. 2022, 73, 5514–5528. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Zhao, Y.; Sun, Y.; Li, Y. NACs, generalist in plant life. Plant Biotechnol. J. 2023, 21, 2433–2457. [Google Scholar] [CrossRef] [PubMed]
- Yeung, E.; van Veen, H.; Vashisht, D.; Sobral Paiva, A.L.; Hummel, M.; Rankenberg, T.; Steffens, B.; Steffen-Heins, A.; Sauter, M.; de Vries, M.; et al. A stress recovery signaling network for enhanced flooding tolerance in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2018, 115, E6085–E6094. [Google Scholar] [CrossRef]
- Tak, H.; Negi, S.; Ganapathi, T.R. Banana NAC transcription factor MusaNAC042 is positively associated with drought and salinity tolerance. Protoplasma 2017, 254, 803–816. [Google Scholar] [CrossRef]
- Mei, F.; Chen, B.; Du, L.; Li, S.; Zhu, D.; Chen, N.; Zhang, Y.; Li, F.; Wang, Z.; Cheng, X.; et al. A gain-of-function allele of a DREB transcription factor gene ameliorates drought tolerance in wheat. Plant Cell 2022, 34, 4472–4494. [Google Scholar] [CrossRef]
- Salladini, E.; Jørgensen, M.L.M.; Theisen, F.F.; Skriver, K. Intrinsic Disorder in Plant Transcription Factor Systems: Functional Implications. Int. J. Mol. Sci. 2020, 21, 9755. [Google Scholar] [CrossRef]
- Tripathi, P.; Rabara, R.C.; Reese, R.N.; Miller, M.A.; Rohila, J.S.; Subramanian, S.; Shen, Q.J.; Morandi, D.; Bücking, H.; Shulaev, V.; et al. A toolbox of genes, proteins, metabolites and promoters for improving drought tolerance in soybean includes the metabolite coumestrol and stomatal development genes. BMC Genom. 2016, 17, 102. [Google Scholar] [CrossRef]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Osakabe, Y.; Qin, F.; Simpson, S.D.; Maruyama, K.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Co-expression of the stress-inducible zinc finger homeodomain ZFHD1 and NAC transcription factors enhances expression of the ERD1 gene in Arabidopsis. Plant J. 2007, 49, 46–63. [Google Scholar] [CrossRef]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K.; et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Chen, L.; Thao, N.P.; Fujiwara, M.; Wong, H.L.; Kuwano, M.; Umemura, K.; Shirasu, K.; Kawasaki, T.; Shimamoto, K. RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 2008, 20, 2265–2279. [Google Scholar] [CrossRef]
- Shyu, C.; Figueroa, P.; Depew, C.L.; Cooke, T.F.; Sheard, L.B.; Moreno, J.E.; Katsir, L.; Zheng, N.; Browse, J.; Howe, G.A. JAZ8 lacks a canonical degron and has an EAR motif that mediates transcriptional repression of jasmonate responses in Arabidopsis. Plant Cell 2012, 24, 536–550. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, J.; Hu, J.; Zhou, J.; Zhang, M.; Cao, Y.; Zhang, L. Picea wilsonii Transcription Factor PwNAC38 Promotes Drought and Salt Tolerance in Transgenic Arabidopsis and Apple Calli. Forests 2022, 13, 1304. [Google Scholar] [CrossRef]
- Shah, S.T.; Pang, C.; Fan, S.; Song, M.; Arain, S.; Yu, S. Isolation and expression profiling of GhNAC transcription factor genes in cotton (Gossypium hirsutum L.) during leaf senescence and in response to stresses. Gene 2013, 531, 220–234. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Du, B.; Yu, M.; Cao, Y.; Liang, K.; Zhang, L. Picea wilsonii NAC31 and DREB2A Cooperatively Activate ERD1 to Modulate Drought Resistance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2024, 25, 2037. https://doi.org/10.3390/ijms25042037
Huang Y, Du B, Yu M, Cao Y, Liang K, Zhang L. Picea wilsonii NAC31 and DREB2A Cooperatively Activate ERD1 to Modulate Drought Resistance in Transgenic Arabidopsis. International Journal of Molecular Sciences. 2024; 25(4):2037. https://doi.org/10.3390/ijms25042037
Chicago/Turabian StyleHuang, Yiming, Bingshuai Du, Mingxin Yu, Yibo Cao, Kehao Liang, and Lingyun Zhang. 2024. "Picea wilsonii NAC31 and DREB2A Cooperatively Activate ERD1 to Modulate Drought Resistance in Transgenic Arabidopsis" International Journal of Molecular Sciences 25, no. 4: 2037. https://doi.org/10.3390/ijms25042037