

LcSAO1, an Unconventional DOXB Clade 2OGD Enzyme from Ligusticum chuanxiong Catalyzes the Biosynthesis of Plant-Derived Natural Medicine Butylphthalide
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
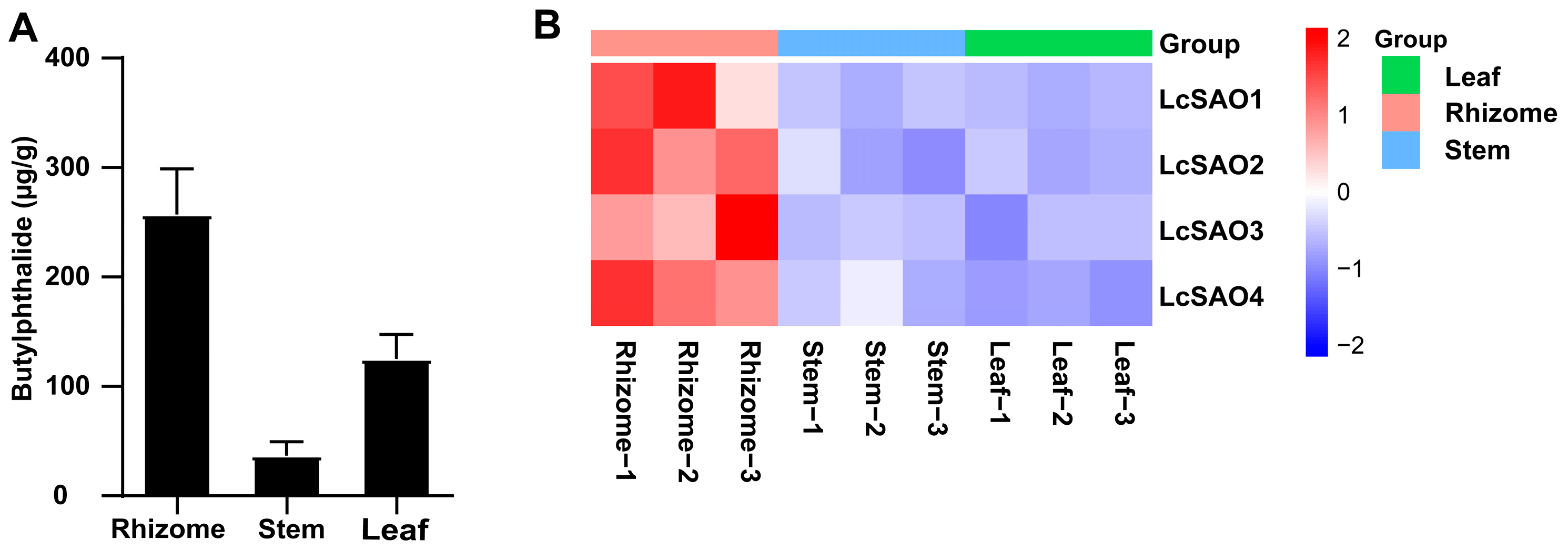
2.1. Butylphthalide Measurement in Different Tissues of L. chuanxiong
2.2. Transcriptome Analysis of L. chuanxiong
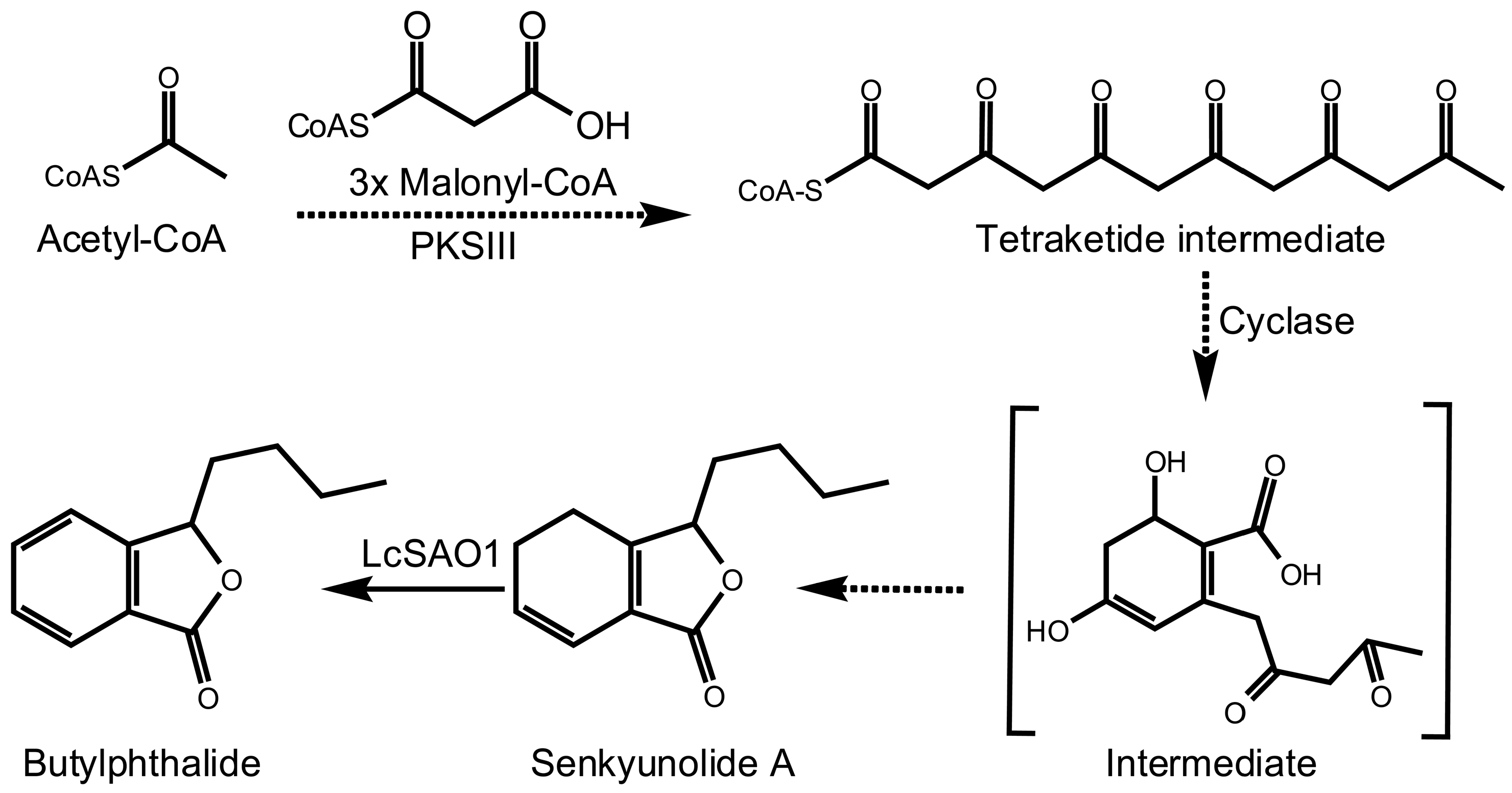
2.3. Screening of Candidate Genes for Butylphthalide
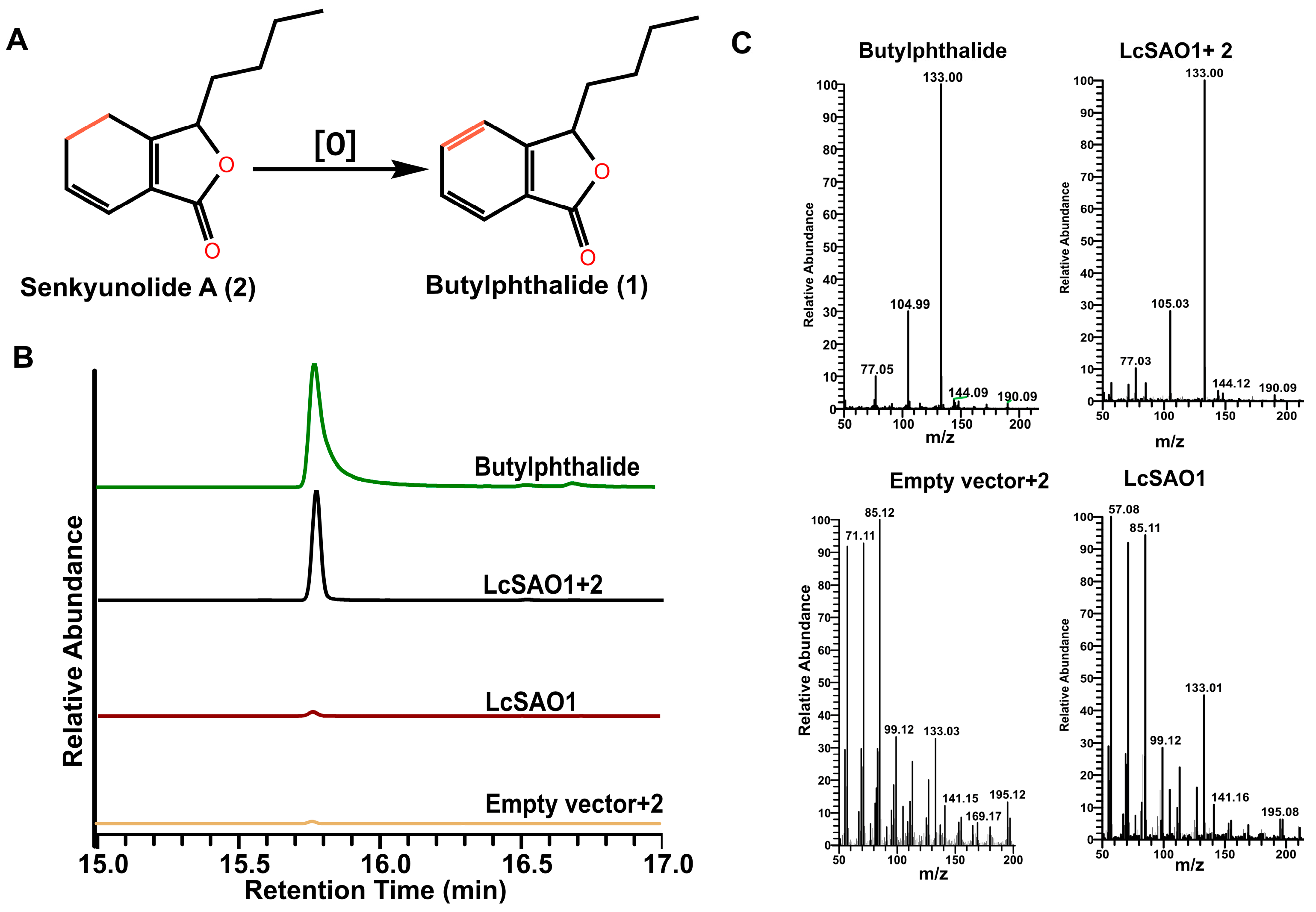
2.4. LcSAO1 Converts Senkyunolide A to Butylphthalide
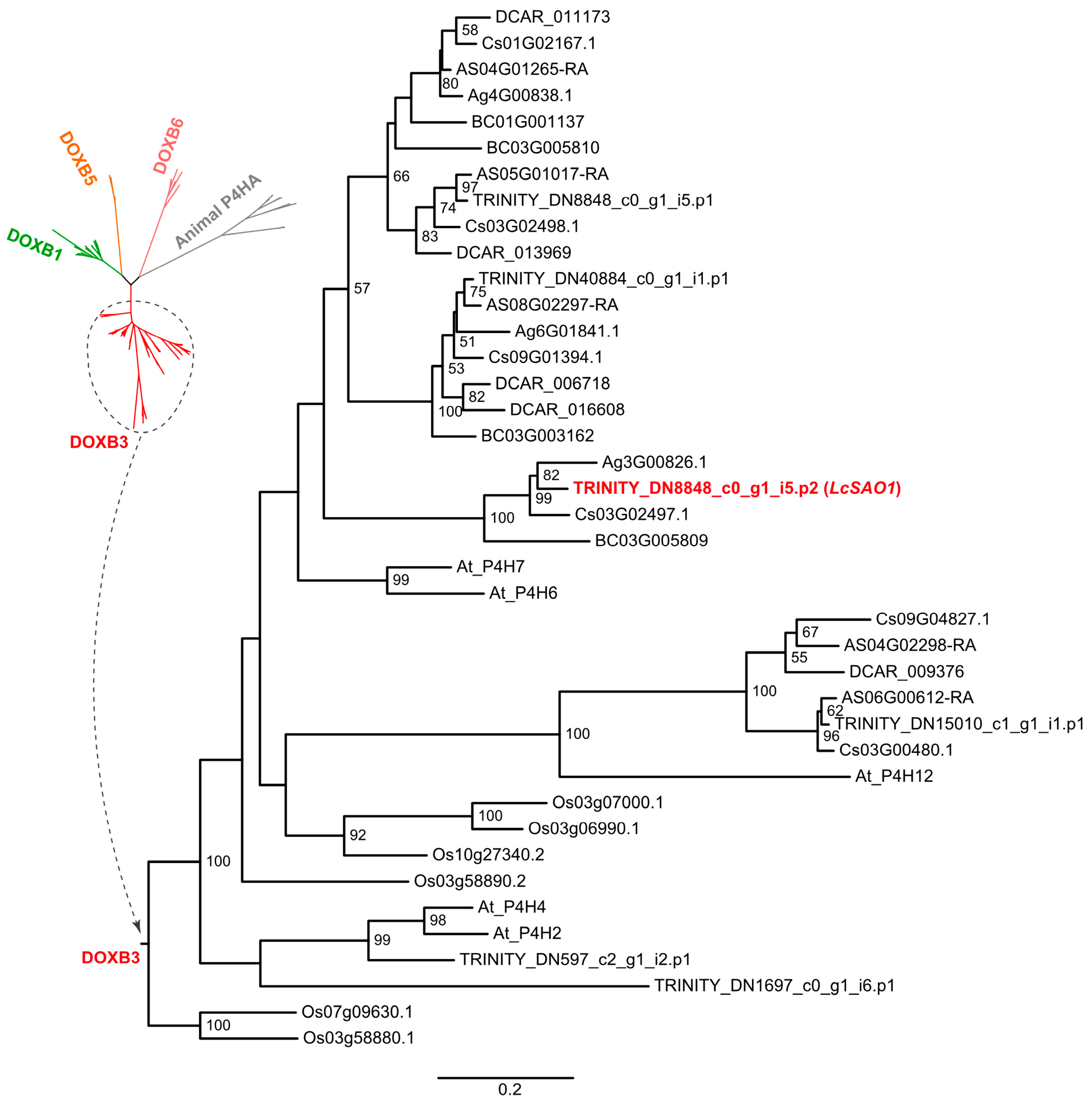
2.5. LcSAO1 Is a Desaturase That Unconventionally Belongs to the DOXB Family
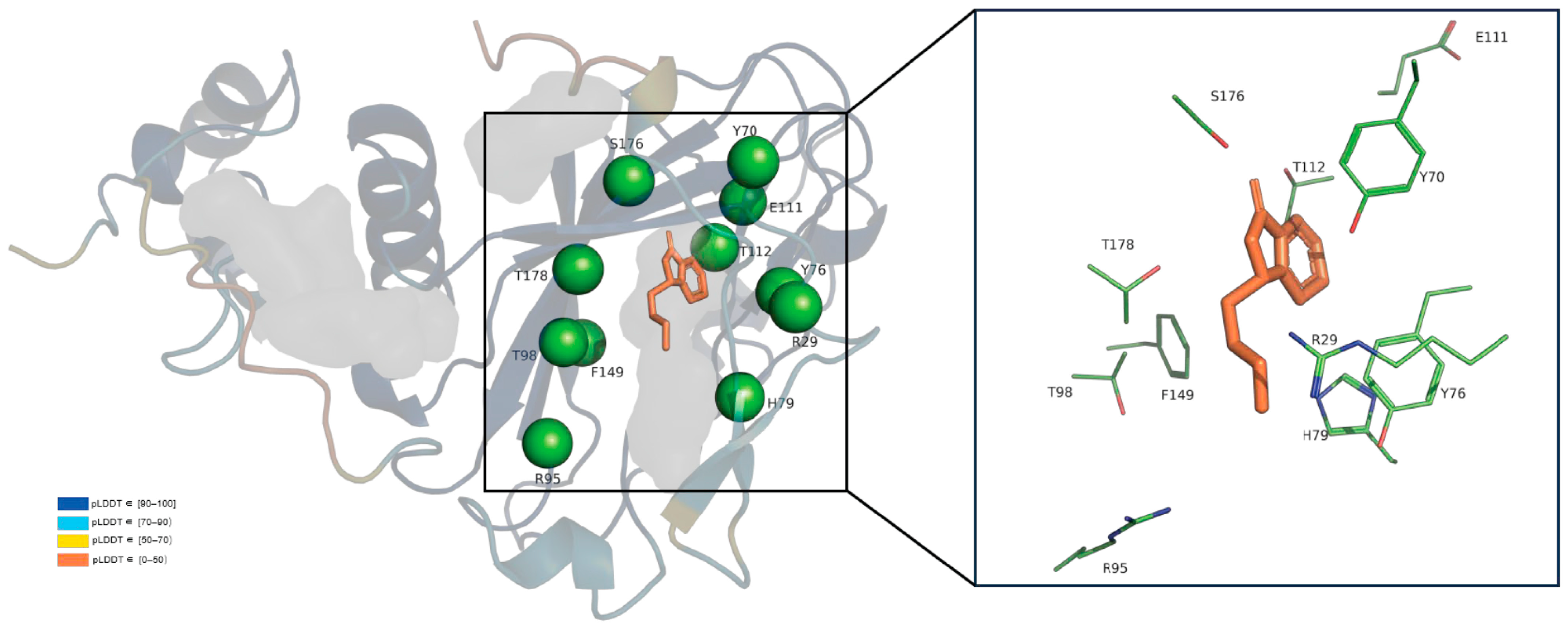
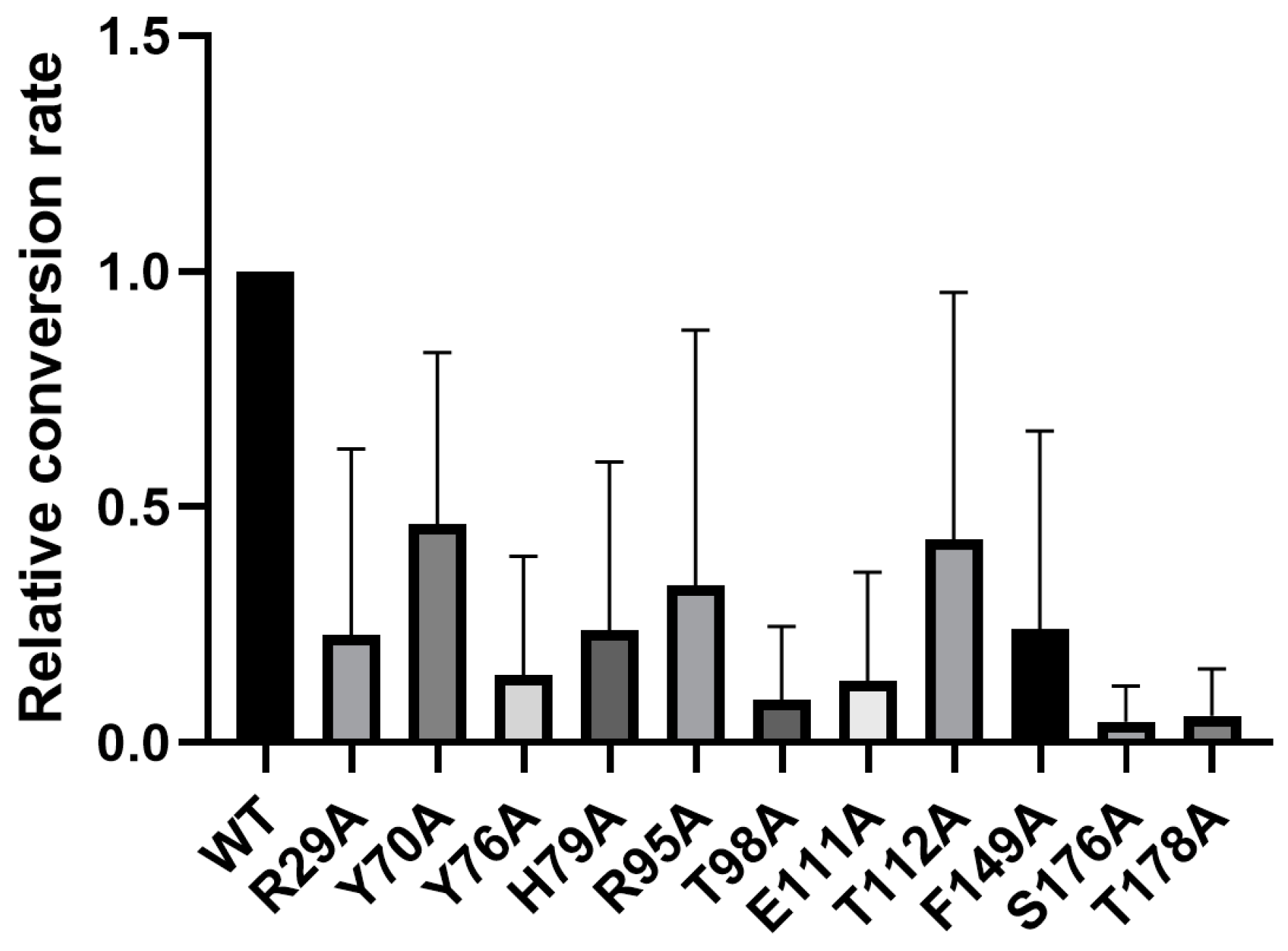
2.6. Key Catalytic Sites Identification Based on Structure Modelling and Activity Assay
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Chemicals, and Reagents
4.2. RNA Extraction and Transcriptome Sequencing
4.3. De Novo Assembly and Functional Annotation of the Transcriptome of L. chuanxiong
4.4. Determination of Butylphthalide Content in L. chuanxiong Tissues
4.5. GC-MS Analysis of Metabolites
4.6. Heterologous Expression of Butylphthalide Precursor Genes in N. benthamiana
4.7. Site-Directed Mutagenesis and Functional Characterization of Mutants
4.8. Heterologous Production, Purification, and In Vitro Reactions with LcSAO1
4.9. Phylogenetic Analysis
4.10. Homology Modelling, Ligand Docking, and Conservation Analysis
4.11. Statistics and Reproducibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, H.; Zhou, Y.; Tang, F.; Wang, C.; Wu, J.; Hu, C.; Xie, X.; Peng, C.; Tan, Y. A Comprehensive Investigation on the Chemical Diversity and Efficacy of Different Parts of Ligusticum chuanxiong. Food Funct. 2022, 13, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.S.-K.; Cheng, T.-Y.; Lin, G. Relaxation Effects of Ligustilide and Senkyunolide A, Two Main Constituents of Ligusticum chuanxiong, in Rat Isolated Aorta. J. Ethnopharmacol. 2007, 111, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Yan, Y.-M.; Yan, H.-L.; Wang, L.-X.; Hu, C.-J.; Wang, H.-L.; Ao, H.; Peng, C.; Tan, Y.-Z. Chuanxiongdiolides R4 and R5, Phthalide Dimers with a Complex Polycyclic Skeleton from the Aerial Parts of Ligusticum chuanxiong and Their Vasodilator Activity. Bioorg. Chem. 2021, 107, 104523. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Liang, S.; Xu, D.-S.; Lin, X.; He, C.-Y.; Feng, Y.; Hong, Y.-L. Effect and Mechanism of Senkyunolide I as an Anti-Migraine Compound from Ligusticum chuanxiong. J. Pharm. Pharmacol. 2011, 63, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yao, M.; Liu, J.; Takagi, N.; Yang, B.; Zhang, M.; Xu, L.; Ren, J.; Fan, X.; Tian, F. Ligusticum chuanxiong Exerts Neuroprotection by Promoting Adult Neurogenesis and Inhibiting Inflammation in the Hippocampus of ME Cerebral Ischemia Rats. J. Ethnopharmacol. 2020, 249, 112385. [Google Scholar] [CrossRef]
- Li, M.; Zhao, C.; Wong, R.N.; Goto, S.; Wang, Z.; Liao, F. Inhibition of Shear-induced Platelet Aggregation in Rat by Tetramethylpyrazine and Salvianolic Acid B. Clin. Hemorheol. Microcirc. 2004, 31, 97–103. [Google Scholar] [PubMed]
- Li, C.-M.; Guo, Y.-Q.; Dong, X.-L.; Li, H.; Wang, B.; Wu, J.-H.; Wong, M.-S.; Chan, S.-W. Ethanolic Extract of Rhizome of Ligusticum chuanxiong Hort.(Chuanxiong) Enhances Endothelium-Dependent Vascular Reactivity in Ovariectomized Rats Fed with High-Fat Diet. Food Funct. 2014, 5, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-M.; Wu, J.-H.; Yang, R.-F.; Dong, X.-L.; He, Z.-Y.; Tian, X.-L.; Guo, D.-J.; Wong, M.-S.; Qiu, T.-Q.; Chan, S.-W. Ligusticum chuanxiong Prevents Ovariectomy-Induced Liver and Vascular Damage in Rats. Am. J. Chin. Med. 2013, 41, 831–848. [Google Scholar] [CrossRef]
- Yuan, J.-F.; Zhang, Z.-Q.; Fan, Z.-C.; Yang, J.-X. Antioxidant Effects and Cytotoxicity of Three Purified Polysaccharides from Ligusticum chuanxiong Hort. Carbohydr. Polym. 2008, 74, 822–827. [Google Scholar] [CrossRef]
- Jeong, J.B.; Ju, S.Y.; Park, J.H.; Lee, J.R.; Yun, K.W.; Kwon, S.T.; Lim, J.-H.; Chung, G.Y.; Jeong, H.J. Antioxidant Activity in Essential Oils of Cnidium Officinale Makino and Ligusticum chuanxiong Hort and Their Inhibitory Effects on DNA Damage and Apoptosis Induced by Ultraviolet B in Mammalian Cell. Cancer Epidemiol. 2009, 33, 41–46. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, C.; Gao, F.; Fu, Q.; Fu, C.; He, Y.; Zhang, J. A Systematic Review on the Rhizome of Ligusticum chuanxiong Hort. (Chuanxiong). Food Chem. Toxicol. 2018, 119, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Long, Y.; Yu, S.; Shi, A.; Wan, J.; Wen, J.; Li, X.; Liu, S.; Zhang, Y.; Li, N. Research Advances in Cardio-Cerebrovascular Diseases of Ligusticum chuanxiong Hort. Front. Pharmacol. 2022, 12, 4116. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.J.; Chou, S.-C. The Structural Diversity of Phthalides from the Apiaceae. J. Nat. Prod. 2007, 70, 891–900. [Google Scholar] [CrossRef]
- Del-Ángel, M.; Nieto, A.; Ramírez-Apan, T.; Delgado, G. Anti-Inflammatory Effect of Natural and Semi-Synthetic Phthalides. Eur. J. Pharmacol. 2015, 752, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Peng, C.; Guo, L.; Feng, R.; Shu, H.-Z.; Tian, Y.-C.; Zhou, Q.-M.; Xiong, L. Six Pairs of Enantiomeric Phthalide Dimers from the Rhizomes of Ligusticum chuanxiong and Their Absolute Configurations and Anti-Inflammatory Activities. Bioorg. Chem. 2022, 127, 105970. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Chan, S.S.-K.; Chung, H.-S.; Li, S.-L. Chemistry and Biological Activities of Naturally Occurring Phthalides. Stud. Nat. Prod. Chem. 2005, 32, 611–669. [Google Scholar] [CrossRef]
- Huang, S.; Chen, J.; Li, W.; Song, S.; Li, X.; Yu, H.; Long, F.; Chen, R.; Bao, X.; Chan, K. Comparison of Volatile Compositions among Four Related Ligusticum chuanxiong Herbs by HS-SPME-GC-MS. Processes 2023, 11, 196. [Google Scholar] [CrossRef]
- Huang, H.; Qiu, M.; Lin, J.; Li, M.; Ma, X.; Ran, F.; Luo, C.; Wei, X.; Xu, R.; Tan, P.; et al. Potential Effect of Tropical Fruits Phyllanthus emblica L. for the Prevention and Management of Type 2 Diabetic Complications: A Systematic Review of Recent Advances. Eur. J. Nutr. 2021, 60, 3525–3542. [Google Scholar] [CrossRef]
- Kobayashi, S.; Nagasawa, S.; Yamamoto, Y.; Donghyo, K.; Bamba, T.; Fukusaki, E. Metabolic Profiling and Identification of the Genetic Varieties and Agricultural Origin of Cnidium Officinale and Ligusticum chuanxiong. J. Biosci. Bioeng. 2012, 114, 86–91. [Google Scholar] [CrossRef]
- Liu, L.; Shi, B.; Li, J.; Wen, J.; Zhou, L.; He, Y. Assessing Environmental Suitability of Ligusticum chuanxiong Based on Ecological Analyses with Chemical and Molecular Verification. Heliyon 2023, 9, e14629. [Google Scholar] [CrossRef]
- Peng, Y.; Zeng, X.; Feng, Y.; Wang, X. Antiplatelet and Antithrombotic Activity of L-3-n-Butylphthalide in Rats. J. Cardiovasc. Pharmacol. 2004, 43, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, H.; Nomura, M. Studies on the Consitituents of Umbelliferae Plants. XII. Biogenesis of 3-Butylphthalide (1). Chem. Pharm. Bull. 1966, 14, 777–778. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Yang, J.; Ren, J.; Wang, A.; Ji, T.; Su, Y. Bioactive Phthalides from Ligusticum Sinense Oliv Cv. Chaxiong. Fitoterapia 2014, 93, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Zuo, A.H.; Wang, L.; Xiao, H.B. Study on Degradation Products of Senkyunolide A and Senkyunolide I. Chin. Tradit. Herbal Drugs 2012, 43, 2127–2131. [Google Scholar]
- Nie, B.; Chen, X.; Hou, Z.; Li, C.; Sun, W.; Ji, J.; Zang, L.; Yang, S.; Fan, P.; Zhang, W.; et al. Haplotype-Phased Genome Revealed the Butylphthalide Biosynthesis and Hybrid Origin of Ligusticum chuanxiong. bioRxiv 2023, 2023.06.13.544868. [Google Scholar] [CrossRef]
- Nadi, R.; Mateo-Bonmatí, E.; Juan-Vicente, L.; Micol, J.L. The 2OGD Superfamily: Emerging Functions in Plant Epigenetics and Hormone Metabolism. Mol. Plant 2018, 11, 1222–1224. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, Y.; Li, K.; Yang, D.; Liu, N.; Zhang, L.; Zhao, L.; Zhang, X.; Liu, Y.; Gao, L.; et al. Roles of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily in the Flavonoid Pathway: A Review of the Functional Diversity of F3H, FNS I, FLS, and LDOX/ANS. Molecules 2021, 26, 6745. [Google Scholar] [CrossRef]
- Song, J.-J.; Fang, X.; Li, C.-Y.; Jiang, Y.; Li, J.-X.; Wu, S.; Guo, J.; Liu, Y.; Fan, H.; Huang, Y.-B. A 2-Oxoglutarate-Dependent Dioxygenase Converts Dihydrofuran to Furan in Salvia Diterpenoids. Plant Physiol. 2022, 188, 1496–1506. [Google Scholar] [CrossRef]
- Parkinson, E.I.; Lakkis, H.G.; Alwali, A.A.; Metcalf, M.E.M.; Modi, R.; Metcalf, W.W. An Unusual Oxidative Rearrangement Catalyzed by a Divergent Member of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily during Biosynthesis of Dehydrofosmidomycin. Angew. Chem. Int. Ed. 2022, 61, e202206173. [Google Scholar] [CrossRef]
- Kim, C.Y.; Mitchell, A.J.; Glinkerman, C.M.; Li, F.-S.; Pluskal, T.; Weng, J.-K. The Chloroalkaloid (-)-Acutumine Is Biosynthesized via a Fe (II)-and 2-Oxoglutarate-Dependent Halogenase in Menispermaceae Plants. Nat. Commun. 2020, 11, 1867. [Google Scholar] [CrossRef]
- Nett, R.S.; Dho, Y.; Low, Y.-Y.; Sattely, E.S. A Metabolic Regulon Reveals Early and Late Acting Enzymes in Neuroactive Lycopodium Alkaloid Biosynthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2102949118. [Google Scholar] [CrossRef] [PubMed]
- García-Contreras, R.; Vos, P.; Westerhoff, H.V.; Boogerd, F.C. Why in Vivo May Not Equal in Vitro–New Effectors Revealed by Measurement of Enzymatic Activities under the Same in Vivo-like Assay Conditions. FEBS J. 2012, 279, 4145–4159. [Google Scholar] [CrossRef] [PubMed]
- Fleishman, S.J.; Leaver-Fay, A.; Corn, J.E.; Strauch, E.-M.; Khare, S.D.; Koga, N.; Ashworth, J.; Murphy, P.; Richter, F.; Lemmon, G. RosettaScripts: A Scripting Language Interface to the Rosetta Macromolecular Modeling Suite. PLoS ONE 2011, 6, e20161. [Google Scholar] [CrossRef] [PubMed]
- Myllykoski, M.; Sutinen, A.; Koski, M.K.; Kallio, J.P.; Raasakka, A.; Myllyharju, J.; Wierenga, R.K.; Koivunen, P. Structure of Transmembrane Prolyl 4-Hydroxylase Reveals Unique Organization of EF and Dioxygenase Domains. J. Biol. Chem. 2021, 296, 100197. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T. Protein Secondary Structure Prediction Based on Position-Specific Scoring Matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kamisetty, H.; Ovchinnikov, S.; Baker, D. Assessing the Utility of Coevolution-Based Residue–Residue Contact Predictions in a Sequence-and Structure-Rich Era. Proc. Natl. Acad. Sci. USA 2013, 110, 15674–15679. [Google Scholar] [CrossRef]
- Ge, C.; Tang, C.; Zhu, Y.-X.; Wang, G.-F. Genome-Wide Identification of the Maize 2OGD Superfamily Genes and Their Response to Fusarium Verticillioides and Fusarium Graminearum. Gene 2021, 764, 145078. [Google Scholar] [CrossRef]
- Hieta, R.; Myllyharju, J. Cloning and Characterization of a Low Molecular Weight Prolyl 4-Hydroxylase from Arabidopsis Thaliana: Effective Hydroxylation of Proline-Rich, Collagen-like, and Hypoxia-Inducible Transcription Factor α-like Peptides. J. Biol. Chem. 2002, 277, 23965–23971. [Google Scholar] [CrossRef]
- Velasquez, S.M.; Ricardi, M.M.; Poulsen, C.P.; Oikawa, A.; Dilokpimol, A.; Halim, A.; Mangano, S.; Juarez, S.P.D.; Marzol, E.; Salter, J.D.S. Complex Regulation of Prolyl-4-Hydroxylases Impacts Root Hair Expansion. Mol. Plant 2015, 8, 734–746. [Google Scholar] [CrossRef]
- Dunham, N.P.; Chang, W.; Mitchell, A.J.; Martinie, R.J.; Zhang, B.; Bergman, J.A.; Rajakovich, L.J.; Wang, B.; Silakov, A.; Krebs, C. Two Distinct Mechanisms for C–C Desaturation by Iron (II)-and 2-(Oxo) Glutarate-Dependent Oxygenases: Importance of α-Heteroatom Assistance. J. Am. Chem. Soc. 2018, 140, 7116–7126. [Google Scholar] [CrossRef]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and Diversity of the 2–Oxoglutarate-dependent Dioxygenase Superfamily in Plants. Plant J. 2014, 78, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Nakashima, Y.; Chen, H.; Hoshino, S.; Mitsuhashi, T.; Abe, I. Structure-Based Redesign of Fe(II)/2-Oxoglutarate-Dependent Oxygenase AndA to Catalyze Spiro-Ring Formation. Chem. Commun. 2022, 58, 5510–5513. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Kourmpetli, S.; Ward, D.A.; Thomas, S.G.; Gong, F.; Powers, S.J.; Carrera, E.; Taylor, B.; de Caceres Gonzalez, F.N.; Tudzynski, B. Characterization of the Fungal Gibberellin Desaturase as a 2-Oxoglutarate-Dependent Dioxygenase and Its Utilization for Enhancing Plant Growth. Plant Physiol. 2012, 160, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Koski, M.K.; Hieta, R.; Bollner, C.; Kivirikko, K.I.; Myllyharju, J.; Wierenga, R.K. The Active Site of an Algal Prolyl 4-Hydroxylase Has a Large Structural Plasticity. J. Biol. Chem. 2007, 282, 37112–37123. [Google Scholar] [CrossRef] [PubMed]
- Duric, K.; Liu, Y.; Chen, S.-N.; Lankin, D.C.; Nikolic, D.; McAlpine, J.B.; Friesen, J.B.; Pauli, G.F. Studying Mass Balance and the Stability of (Z)-Ligustilide from Angelica Sinensis Helps to Bridge a Botanical Instability–Bioactivity Chasm. J. Nat. Prod. 2019, 82, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Bartosińska, E.; Buszewska-Forajta, M.; Siluk, D. GC–MS and LC–MS Approaches for Determination of Tocopherols and Tocotrienols in Biological and Food Matrices. J. Pharm. Biomed. Anal. 2016, 127, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, J.; Lee, F.S.-C.; Wang, X. GC-MS Fingerprints for Discrimination of Ligusticum chuanxiong from Angelica. J. Sep. Sci. 2008, 31, 3231–3237. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Zhang, X.; Sun, W.; Hou, Z.; Nie, B.; Wang, F.; Yang, S.; Feng, S.; Li, W.; Wang, L. LcSAO1, an Unconventional DOXB Clade 2OGD Enzyme from Ligusticum chuanxiong Catalyzes the Biosynthesis of Plant-Derived Natural Medicine Butylphthalide. Int. J. Mol. Sci. 2023, 24, 17417. https://doi.org/10.3390/ijms242417417
Chen X, Zhang X, Sun W, Hou Z, Nie B, Wang F, Yang S, Feng S, Li W, Wang L. LcSAO1, an Unconventional DOXB Clade 2OGD Enzyme from Ligusticum chuanxiong Catalyzes the Biosynthesis of Plant-Derived Natural Medicine Butylphthalide. International Journal of Molecular Sciences. 2023; 24(24):17417. https://doi.org/10.3390/ijms242417417
Chicago/Turabian StyleChen, Xueqing, Xiaopeng Zhang, Wenkai Sun, Zhuangwei Hou, Bao Nie, Fengjiao Wang, Song Yang, Shourui Feng, Wei Li, and Li Wang. 2023. "LcSAO1, an Unconventional DOXB Clade 2OGD Enzyme from Ligusticum chuanxiong Catalyzes the Biosynthesis of Plant-Derived Natural Medicine Butylphthalide" International Journal of Molecular Sciences 24, no. 24: 17417. https://doi.org/10.3390/ijms242417417