
Improved 2,3-Butanediol Production Rate of Metabolically Engineered Saccharomyces cerevisiae by Deletion of RIM15 and Activation of Pyruvate Consumption Pathway

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
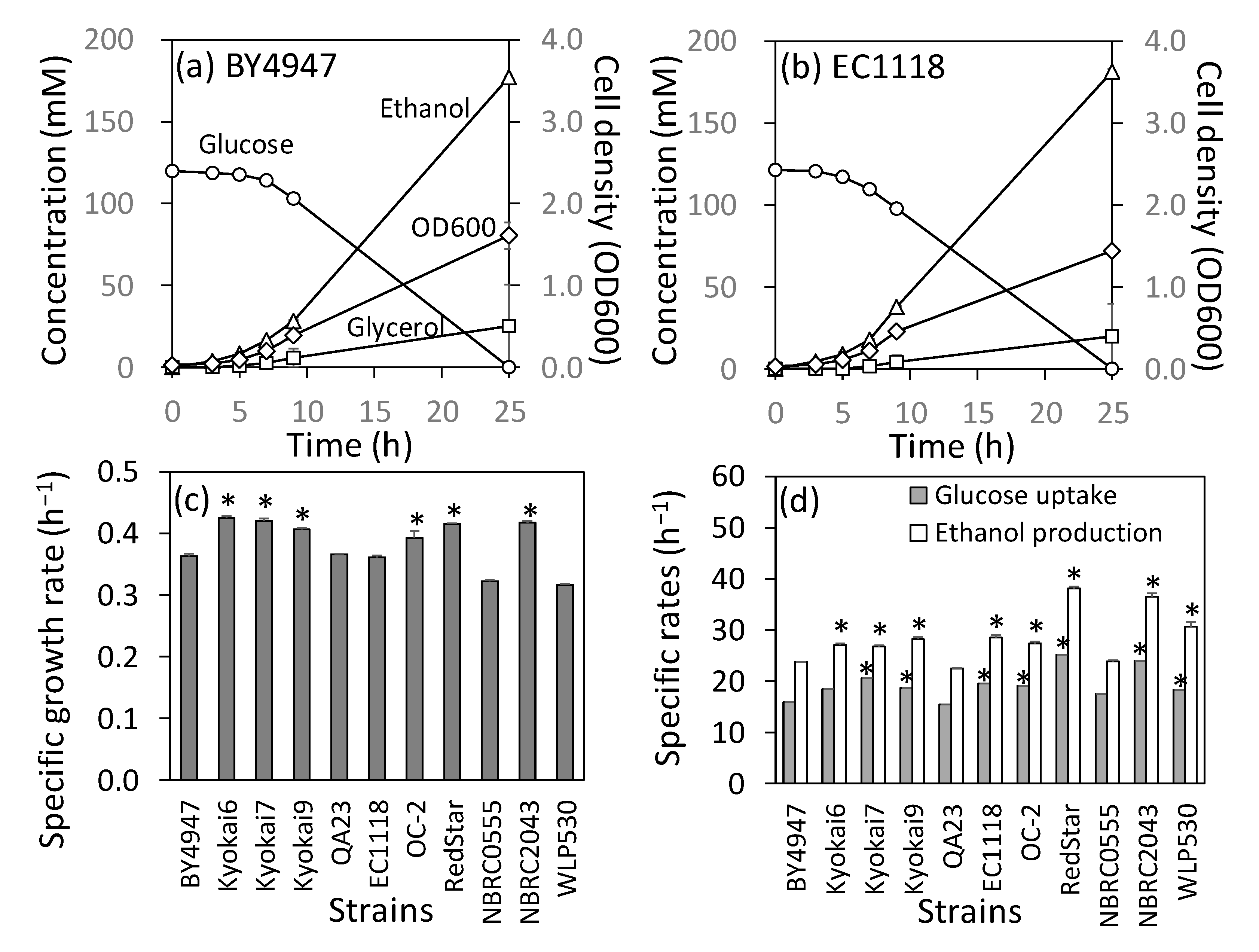
2.1. Culture Profiles and RIM15 Mutations in 11 Industrial Diploid Strains
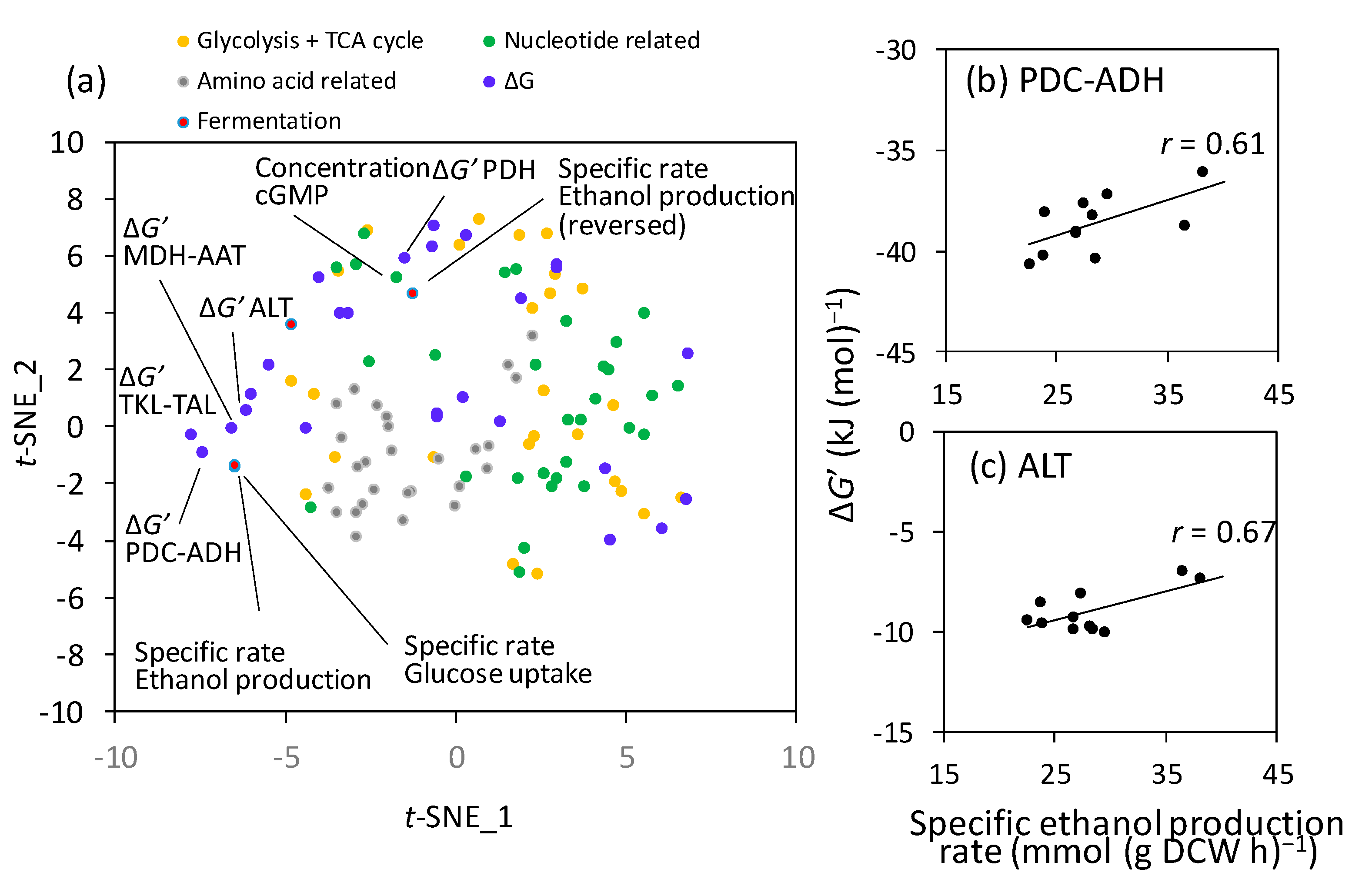
2.2. Correlation between the Intracellular Metabolite State and Ethanol Production Rate
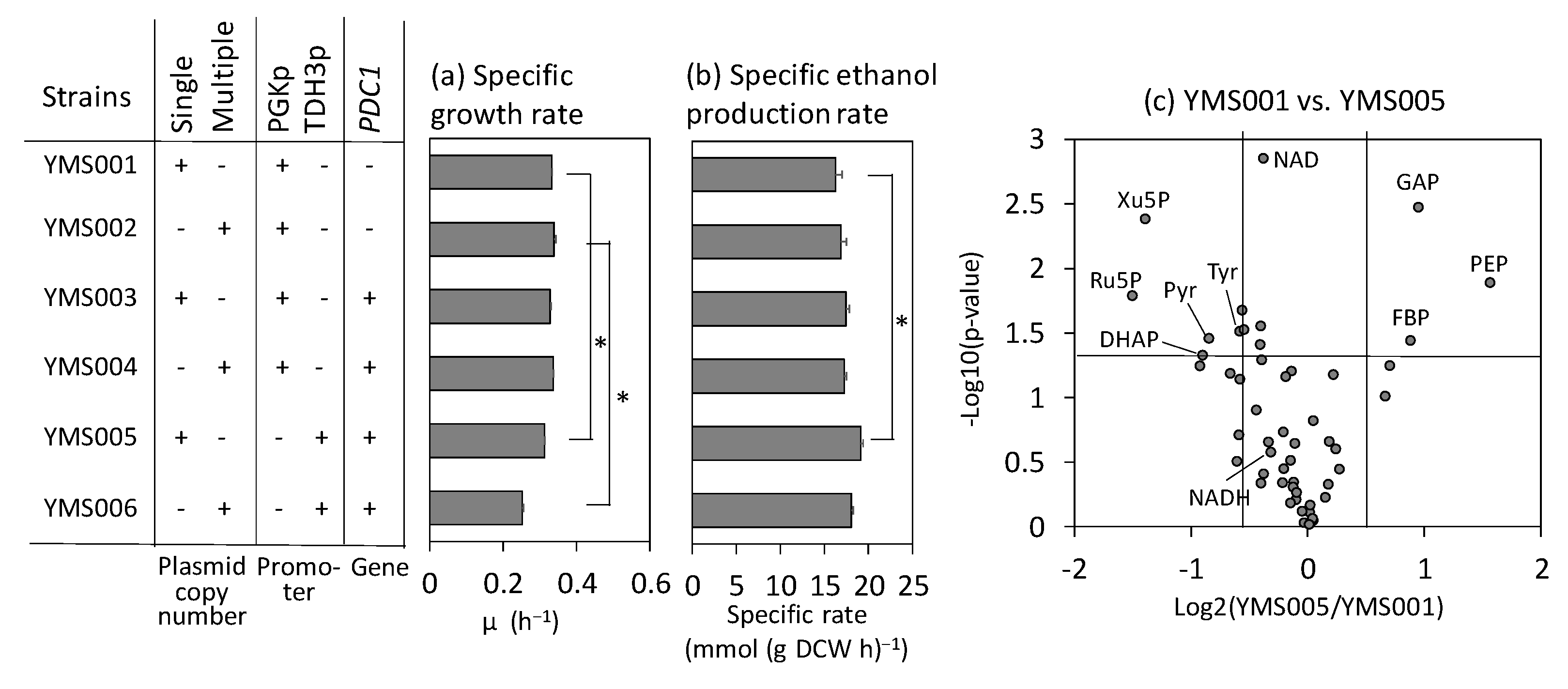
2.3. Elevation of Ethanol Production in the Laboratory Strain by Activating the Pyruvate-Consuming Reaction
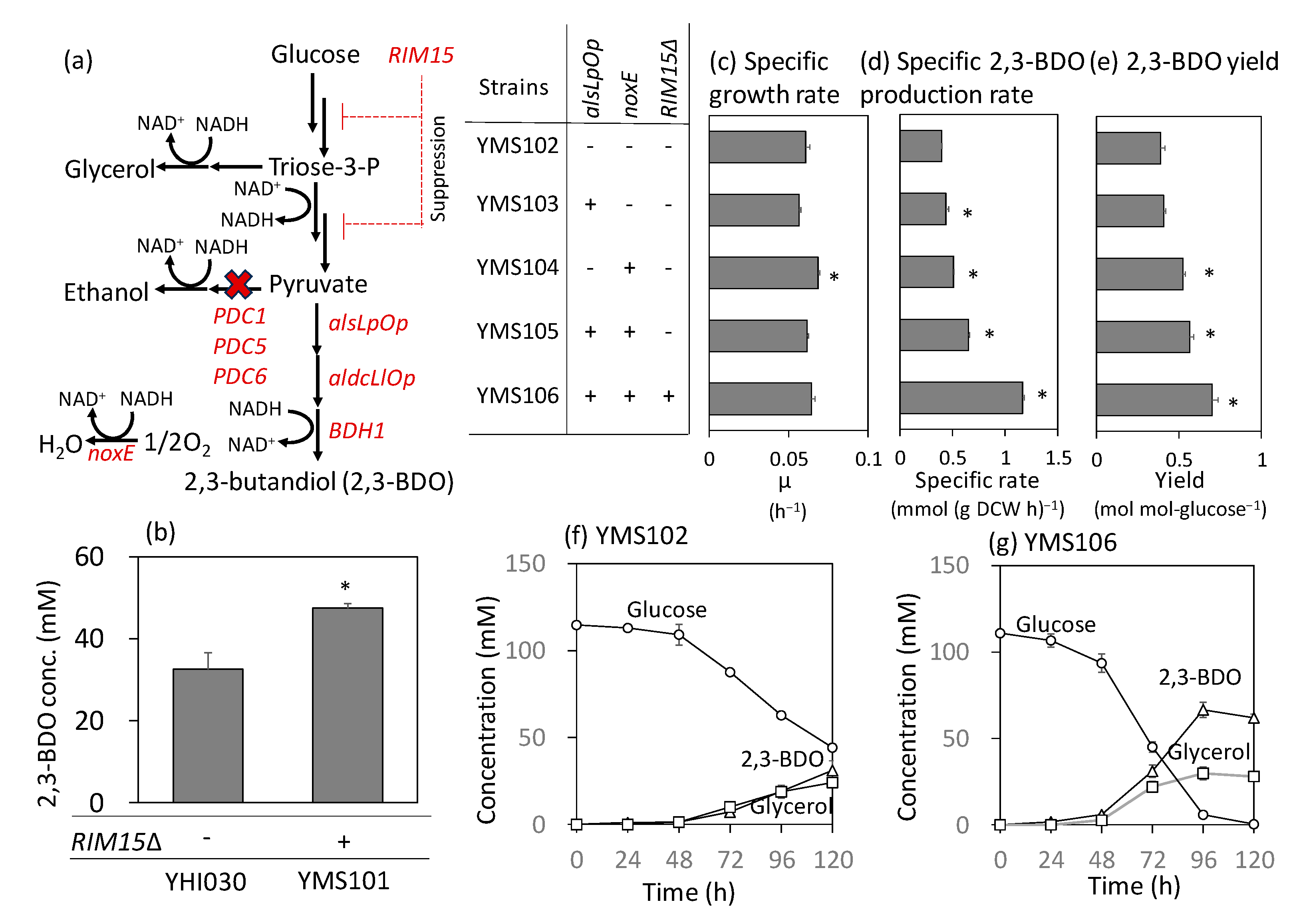
2.4. Implementation of RIM15 Deletion and Enhanced Pyruvate Consumption in a Metabolically Engineered S. cerevisiae Strain Producing 2,3-Butanediol
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Condition
4.2. Measurement of Extracellular Metabolites
4.3. Metabolome Analysis
4.4. Calculation of ΔG′ and Multivariate Analysis
4.5. Construction of Metabolically Engineered Strains
4.6. RIM15 Deletion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nielsen, J.; Larsson, C.; van Maris, A.; Pronk, J. Metabolic engineering of yeast for production of fuels and chemicals. Curr. Opin. Biotechnol. 2013, 24, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, R.; Yamada, R.; Matsumoto, T.; Ogino, H. Bioengineering for the industrial production of 2,3-butanediol by the yeast, Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2022, 38, 38. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, J.W.; Lee, Y.G.; Park, Y.C.; Seo, J.H. Metabolic engineering of Saccharomyces cerevisiae for 2,3-butanediol production. Appl. Microbiol. Biotechnol. 2017, 101, 2241–2250. [Google Scholar] [CrossRef]
- Xu, Y.; Chu, H.; Gao, C.; Tao, F.; Zhou, Z.; Li, K.; Li, L.; Ma, C.; Xu, P. Systematic metabolic engineering of Escherichia coli for high-yield production of fuel bio-chemical 2,3-butanediol. Metab. Eng. 2014, 23, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Jung, M.Y.; Lee, J.; Oh, M.K. Production of 2,3-butanediol in Saccharomyces cerevisiae by in silico aided metabolic engineering. Microb. Cell Fact. 2012, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Sim, H.J.; Kim, J.W.; Lee, Y.G.; Park, Y.C.; Seo, J.H. Enhanced production of 2,3-butanediol from xylose by combinatorial engineering of xylose metabolic pathway and cofactor regeneration in pyruvate decarboxylase-deficient Saccharomyces cerevisiae. Bioresour. Technol. 2017, 245, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Huo, G.; Foulquie-Moreno, M.R.; Thevelein, J.M. Development of an industrial yeast strain for efficient production of 2,3-butanediol. Microb. Cell Fact. 2022, 21, 199. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Geng, A. High-copy genome integration of 2,3-butanediol biosynthesis pathway in Saccharomyces cerevisiae via in vivo DNA assembly and replicative CRISPR-Cas9 mediated delta integration. J. Biotechnol. 2020, 310, 13–20. [Google Scholar] [CrossRef]
- Yamada, R.; Wakita, K.; Mitsui, R.; Nishikawa, R.; Ogino, H. Efficient production of 2,3-butanediol by recombinant Saccharomyces cerevisiae through modulation of gene expression by cocktail delta-integration. Bioresour. Technol. 2017, 245, 1558–1566. [Google Scholar] [CrossRef]
- Kim, J.W.; Seo, S.O.; Zhang, G.C.; Jin, Y.S.; Seo, J.H. Expression of Lactococcus lactis NADH oxidase increases 2,3-butanediol production in Pdc-deficient Saccharomyces cerevisiae. Bioresour. Technol. 2015, 191, 512–519. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, J.; Seo, S.O.; Kim, K.H.; Jin, Y.S.; Seo, J.H. Enhanced production of 2,3-butanediol by engineered Saccharomyces cerevisiae through fine-tuning of pyruvate decarboxylase and NADH oxidase activities. Biotechnol. Biofuels 2016, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hahn, J.S. Efficient production of 2,3-butanediol in Saccharomyces cerevisiae by eliminating ethanol and glycerol production and redox rebalancing. Metab. Eng. 2015, 31, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Ishii, J.; Morita, K.; Ida, K.; Kato, H.; Kinoshita, S.; Hataya, S.; Shimizu, H.; Kondo, A.; Matsuda, F. A pyruvate carbon flux tugging strategy for increasing 2,3-butanediol production and reducing ethanol subgeneration in the yeast Saccharomyces cerevisiae. Biotechnol. Biofuels 2018, 11, 180. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Seo, J.H. Production of 2,3-butanediol from glucose and cassava hydrolysates by metabolically engineered industrial polyploid Saccharomyces cerevisiae. Biotechnol. Biofuels 2019, 12, 204. [Google Scholar] [CrossRef] [PubMed]
- Ohnuki, S.; Okada, H.; Friedrich, A.; Kanno, Y.; Goshima, T.; Hasuda, H.; Inahashi, M.; Okazaki, N.; Tamura, H.; Nakamura, R.; et al. Phenotypic diagnosis of lineage and differentiation during sake yeast breeding. G3 2017, 7, 2807–2820. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Kaneko, A.; Sugimoto, Y.; Ohnuki, S.; Takagi, H.; Ohya, Y. Promoter engineering of the Saccharomyces cerevisiae RIM15 gene for improvement of alcoholic fermentation rates under stress conditions. J. Biosci. Bioeng. 2017, 123, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Takagi, H. Pleiotropic functions of the yeast Greatwall-family protein kinase Rim15p: A novel target for the control of alcoholic fermentation. Biosci. Biotechnol. Biochem. 2017, 81, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Uebayashi, K.; Shimizu, H.; Matsuda, F. Comparative analysis of fermentation and enzyme expression profiles among industrial Saccharomyces cerevisiae strains. Appl. Microbiol. Biotechnol. 2018, 102, 7071–7081. [Google Scholar] [CrossRef]
- Matsuda, F.; Toya, Y.; Shimizu, H. Learning from quantitative data to understand central carbon metabolism. Biotechnol. Adv. 2017, 35, 971–980. [Google Scholar] [CrossRef]
- Nagai, H.; Masuda, A.; Toya, Y.; Matsuda, F.; Shimizu, H. Metabolic engineering of mevalonate-producing Escherichia coli strains based on thermodynamic analysis. Metab. Eng. 2018, 47, 1–9. [Google Scholar] [CrossRef]
- Noor, E.; Bar-Even, A.; Flamholz, A.; Reznik, E.; Liebermeister, W.; Milo, R. Pathway thermodynamics highlights kinetic obstacles in central metabolism. PLoS Comput. Biol. 2014, 10, e1003483. [Google Scholar] [CrossRef]
- Park, J.O.; Rubin, S.A.; Xu, Y.F.; Amador-Noguez, D.; Fan, J.; Shlomi, T.; Rabinowitz, J.D. Metabolite concentrations, fluxes and free energies imply efficient enzyme usage. Nat. Chem. Biol. 2016, 12, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Araki, Y.; Zhou, Y.; Maeya, N.; Akao, T.; Shimoi, H. A loss-of-function mutation in the PAS kinase Rim15p is related to defective quiescence entry and high fermentation rates of Saccharomyces cerevisiae sake yeast strains. Appl. Environ. Microbiol. 2012, 78, 4008–4016. [Google Scholar] [CrossRef] [PubMed]
- Su, S.S.; Mitchell, A.P. Identification of functionally related genes that stimulate early meiotic gene expression in yeast. Genetics 1993, 133, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S. Disruption of RIM15 confers an increased tolerance to heavy metals in Saccharomyces cerevisiae. Biotechnol. Lett. 2020, 42, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Kanesaki, Y.; Yoshikawa, H.; Hirasawa, T. Identification of metabolic engineering targets for improving glycerol assimilation ability of Saccharomyces cerevisiae based on adaptive laboratory evolution and transcriptome analysis. J. Biosci. Bioeng. 2019, 128, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Kajihara, T.; Sugimoto, Y.; Takagi, K.; Mizuno, M.; Zhou, Y.; Chen, J.; Takeda, K.; Tatebe, H.; Shiozaki, K.; et al. Nutrient signaling via the TORC1-Greatwall-PP2A(B55delta) pathway is responsible for the high initial rates of alcoholic fermentation in sake yeast strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2019, 85, e02083-18. [Google Scholar] [CrossRef] [PubMed]
- Pronk, J.T.; Yde Steensma, H.; Van Dijken, J.P. Pyruvate metabolism in Saccharomyces cerevisiae. Yeast 1996, 12, 1607–1633. [Google Scholar] [CrossRef]
- Wu, L.; van Dam, J.; Schipper, D.; Kresnowati, M.T.; Proell, A.M.; Ras, C.; van Winden, W.A.; van Gulik, W.M.; Heijnen, J.J. Short-term metabolome dynamics and carbon, electron, and ATP balances in chemostat-grown Saccharomyces cerevisiae CEN.PK 113-7D following a glucose pulse. Appl. Environ. Microbiol. 2006, 72, 3566–3577. [Google Scholar] [CrossRef]
- van Heerden, J.H.; Wortel, M.T.; Bruggeman, F.J.; Heijnen, J.J.; Bollen, Y.J.; Planque, R.; Hulshof, J.; O’Toole, T.G.; Wahl, S.A.; Teusink, B. Lost in transition: Start-up of glycolysis yields subpopulations of nongrowing cells. Science 2014, 343, 1245114. [Google Scholar] [CrossRef]
- van Heerden, J.H.; Bruggeman, F.J.; Teusink, B. Multi-tasking of biosynthetic and energetic functions of glycolysis explained by supply and demand logic. Bioessays 2014, 37, 34–45. [Google Scholar] [CrossRef] [PubMed]
- van Hoek, P.; Flikweert, M.T.; van der Aart, Q.J.; Steensma, H.Y.; van Dijken, J.P.; Pronk, J.T. Effects of pyruvate decarboxylase overproduction on flux distribution at the pyruvate branch point in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1998, 64, 2133–2140. [Google Scholar] [CrossRef]
- de Assis, L.J.; Zingali, R.B.; Masuda, C.A.; Rodrigues, S.P.; Montero-Lomeli, M. Pyruvate decarboxylase activity is regulated by the Ser/Thr protein phosphatase Sit4p in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Yamada, R.; Nishikawa, R.; Wakita, K.; Ogino, H. Rapid and stable production of 2,3-butanediol by an engineered Saccharomyces cerevisiae strain in a continuous airlift bioreactor. J. Ind. Microbiol. Biotechnol. 2018, 45, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Yatabe, F.; Seike, T.; Okahashi, N.; Ishii, J.; Matsuda, F. Improvement of ethanol and 2,3-butanediol production in Saccharomyces cerevisiae by ATP wasting. Microb. Cell Fact. 2023, 22, 204. [Google Scholar] [CrossRef] [PubMed]
- Borodina, I.; Nielsen, J. Advances in metabolic engineering of yeast Saccharomyces cerevisiae for production of chemicals. Biotechnol. J. 2014, 9, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Krivoruchko, A.; Siewers, V.; Nielsen, J. Opportunities for yeast metabolic engineering: Lessons from synthetic biology. Biotechnol. J. 2011, 6, 262–276. [Google Scholar] [CrossRef]
- Watanabe, D.; Zhou, Y.; Hirata, A.; Sugimoto, Y.; Takagi, K.; Akao, T.; Ohya, Y.; Takagi, H.; Shimoi, H. Inhibitory role of Greatwall-like protein kinase Rim15p in alcoholic fermentation via upregulating the UDP-glucose synthesis pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2016, 82, 340–351. [Google Scholar] [CrossRef]
- Yang, G.; Liu, G.L.; Wang, S.J.; Chi, Z.M.; Chi, Z. Pullulan biosynthesis in yeast-like fungal cells is regulated by the transcriptional activator Msn2 and cAMP-PKA signaling pathway. Int. J. Biol. Macromol. 2020, 157, 591–603. [Google Scholar] [CrossRef]
- Ishii, J.; Izawa, K.; Matsumura, S.; Wakamura, K.; Tanino, T.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. A simple and immediate method for simultaneously evaluating expression level and plasmid maintenance in yeast. J. Biochem. 2009, 145, 701–708. [Google Scholar] [CrossRef]
- Okada, S.; Doi, G.; Nakagawa, S.; Kusumoto, E.; Ito, T. Simple-to-use CRISPR-SpCas9/SaCas9/AsCas12a vector series for genome editing in Saccharomyces cerevisiae. G3 2021, 11, jkab304. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, K.; Izumi, Y.; Hata, K.; Bamba, T. An analyticals system for single-cell metabolomics of typical mammalian cells based on highly sensitive nano-liquid chromatography tandem mass spectrometry. Mass Spectrom 2020, 9, A0080. [Google Scholar] [CrossRef] [PubMed]
- Day, A.; Schneider, C.; Schneider, B.L. Yeast cell synchronization. Methods Mol. Biol. 2004, 241, 55–76. [Google Scholar] [CrossRef]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Milo, R. eQuilibrator--the biochemical thermodynamics calculator. Nucleic Acids Res. 2012, 40, D770–D775. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Fukuda, Y.; Murata, K.; Kimura, A. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 1983, 153, 163–168. [Google Scholar] [CrossRef]
- Katahira, S.; Mizuike, A.; Fukuda, H.; Kondo, A. Ethanol fermentation from lignocellulosic hydrolysate by a recombinant xylose- and cellooligosaccharide-assimilating yeast strain. Appl. Microbiol. Biotechnol. 2006, 72, 1136–1143. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Description | Ref. |
|---|---|---|
| BY4947 | Diploid of S288C (X2180-1A × X2180-1B) | National Bio-Resource Project (NBRP), Japan |
| Kyokai6 | Sake strain | [18] |
| Kyokai7 | Sake strain (NBRC2347) | [18] |
| Kyokai9 | Sake strain (NBRC2377) | [18] |
| QA23™ | Wine strain | Gift from SCETI K.K. Tokyo, Japan |
| Lalvin EC1118™ | Wine strain | Gift from SCETI K.K. Tokyo, Japan |
| OC-2 | Wine strain (NBRC104078) | Purchased from NITE |
| RedStar | Bread strain (NBRC2375) | Purchased from NITE |
| NBRC0555 | Bread strain | Purchased from NITE |
| NBRC2043 | Bread strain | Purchased from NITE |
| WLP530 | Beer strain | Gift from Suntory Co. |
| S288C | Identical to BY27002, MATα mal SUC2 | NBRP Yeast, Japan |
| YPH499 | MATa ura3-52 lys2-801 ade2-101 trp1-Δ63 his3-Δ200 leu2-Δ1 | Stratagene/Agilent Technologies |
| YSM021 (PDCΔ) | YPH499 pdc1Δ pdc5Δ pdc6Δ MTH1-ΔT(L165F) | [13] |
| YSM046 (PDCΔ + evolved) | Laboratory-evolved yeast strain derived from the PDCΔ (YSM021) strain | [13] |
| YHI030 | YSM046 [pATP422-alsLpOp-aldcLlOp/pAT425-BDH1] | [13] |
| YMS001 | YPH499 [pGK414] | This study |
| YMS002 | YPH499 [pGK424] | This study |
| YMS003 | YPH499 [pGK414-PDC1] | This study |
| YMS004 | YPH499 [pGK424-PDC1] | This study |
| YMS005 | YPH499 [pGK414-TDH3p_PDC1] | This study |
| YMS006 | YPH499 [pGK424-TDH3p_PDC1] | This study |
| YMS101 | YHI030 rim15Δ | This study |
| YMS102 | YHI030 [pGK423/pGK424] | This study |
| YMS103 | YHI030 [pGK423-ALS/pGK424] | This study |
| YMS104 | YHI030 [pGK423-ALS/pGK424] | This study |
| YMS105 | YHI030 [pGK423-ALS/pGK424-noxE] | This study |
| YMS106 | YHI030_rim15Δ [pGK423-ALS/pGK424-noxE] | This study |
| Plasmids | Description | Ref. |
| pGK423 | Yeast multi-copy type single-gene expression vector containing PGK1 promoter, PGK1 terminator, 2μ origin, and HIS3 marker | [40] |
| pGK423-ALS | pGK423, expression of the ALS gene by the PGK1 promoter | This study |
| pGK414 | Yeast low-copy type single-gene expression vector containing PGK1 promoter, PGK1 terminator, CEN/ARS ori, and TRP1 marker | [40] |
| pGK424 | Yeast multi-copy type single-gene expression vector containing PGK1 promoter, PGK1 terminator, CEN/ARS ori, and TRP1 marker | [40] |
| pGK414-PDC | pGK414, expression of the PDC1 gene by the PDC1 promoter | This study |
| pGK424-PDC | pGK424, expression of the PDC1 gene by the PDC1 promoter | This study |
| pGK414-TDH3p-PDC1 | pGK414, expression of the PDC1 gene by the TDH3 promoter | This study |
| pGK424-TDHp3-PDC1 | pGK424, expression of the PDC1 gene by the TDH3 promoter | This study |
| pGK424-noxE | pGK424, expression of the noxE gene by the TDH3 promoter | This study |
| pGAL1-Cas9-tADH1-pGAL1-2BsaI-sgRNAFE(empty)-HDV-tCYC1-CU | YCp vector, and URA3 marker | [41] |
| pGAL1-RIM15Δ | pGAL1-Cas9-tADH1-pGAL1-2BsaI-sgRNAFE(RIM15D)-HDV-tCYC1-CU | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugimura, M.; Seike, T.; Okahashi, N.; Izumi, Y.; Bamba, T.; Ishii, J.; Matsuda, F. Improved 2,3-Butanediol Production Rate of Metabolically Engineered Saccharomyces cerevisiae by Deletion of RIM15 and Activation of Pyruvate Consumption Pathway. Int. J. Mol. Sci. 2023, 24, 16378. https://doi.org/10.3390/ijms242216378
Sugimura M, Seike T, Okahashi N, Izumi Y, Bamba T, Ishii J, Matsuda F. Improved 2,3-Butanediol Production Rate of Metabolically Engineered Saccharomyces cerevisiae by Deletion of RIM15 and Activation of Pyruvate Consumption Pathway. International Journal of Molecular Sciences. 2023; 24(22):16378. https://doi.org/10.3390/ijms242216378
Chicago/Turabian StyleSugimura, Masahiko, Taisuke Seike, Nobuyuki Okahashi, Yoshihiro Izumi, Takeshi Bamba, Jun Ishii, and Fumio Matsuda. 2023. "Improved 2,3-Butanediol Production Rate of Metabolically Engineered Saccharomyces cerevisiae by Deletion of RIM15 and Activation of Pyruvate Consumption Pathway" International Journal of Molecular Sciences 24, no. 22: 16378. https://doi.org/10.3390/ijms242216378