
Expression Pattern of Trace Amine-Associated Receptors during Differentiation of Human Pluripotent Stem Cells to Dopaminergic Neurons
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
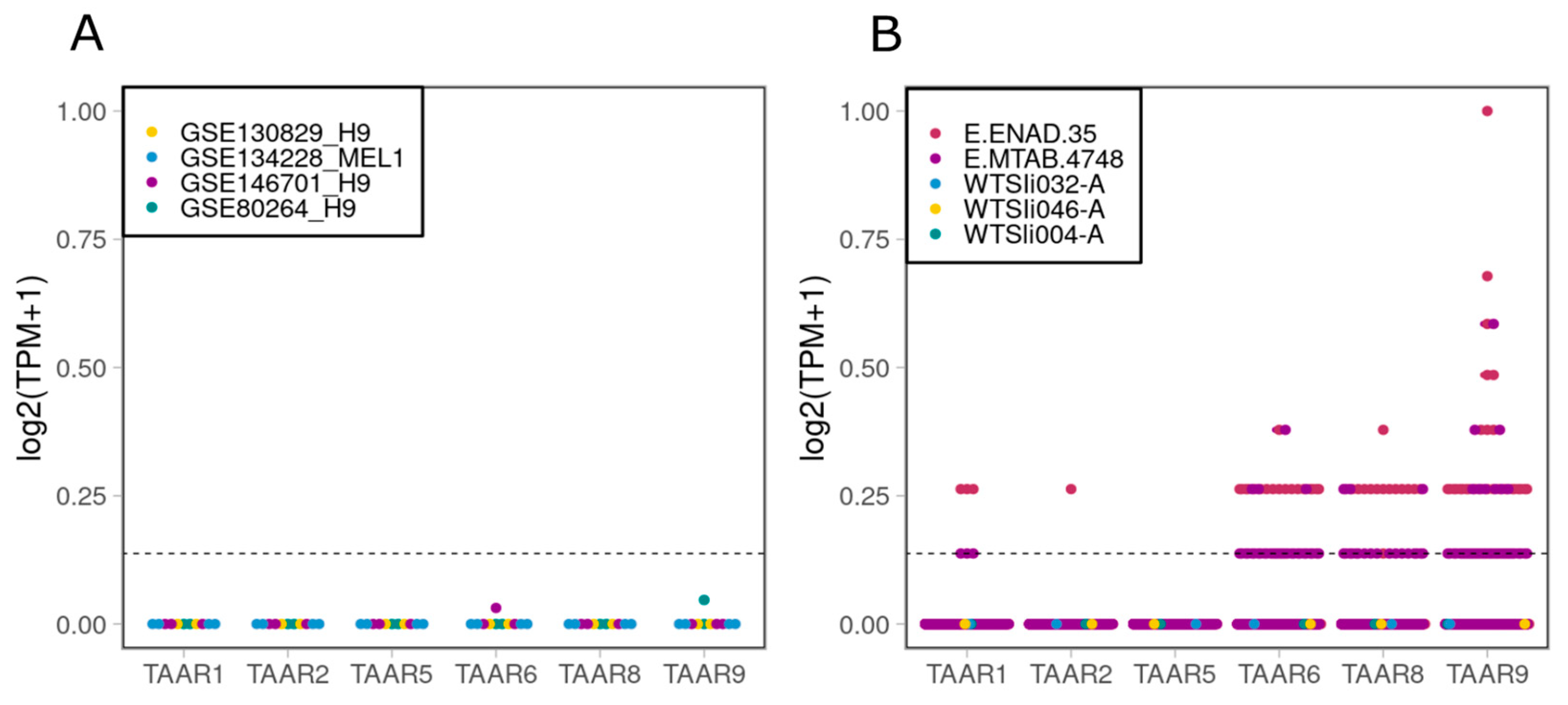
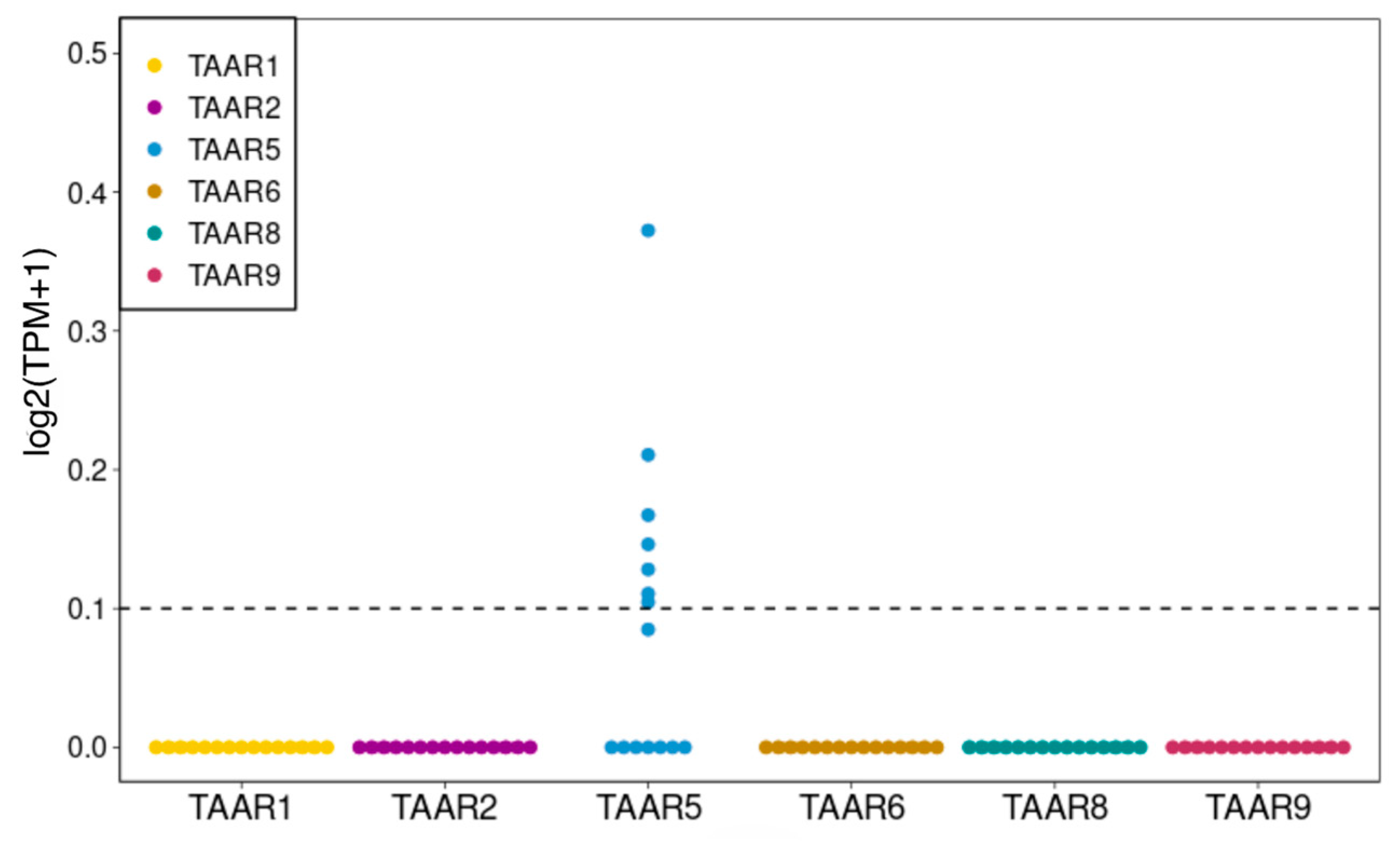
2.1. TAARs Are Expressed at Low Levels in a Number of Human IPSC Lines, but Not in H9 or MEL1 Embryonic Stem Cell (ESC) Lines
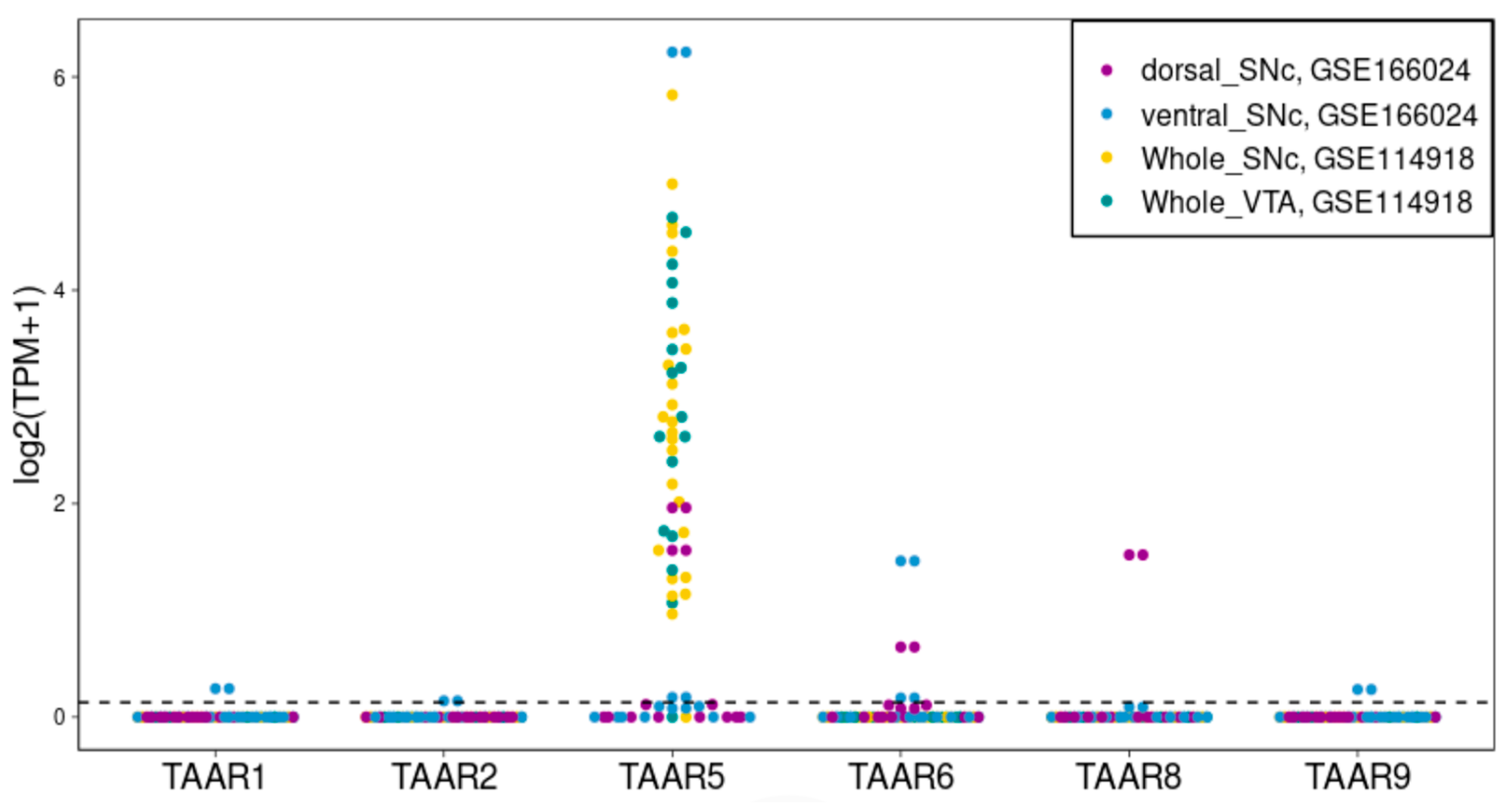
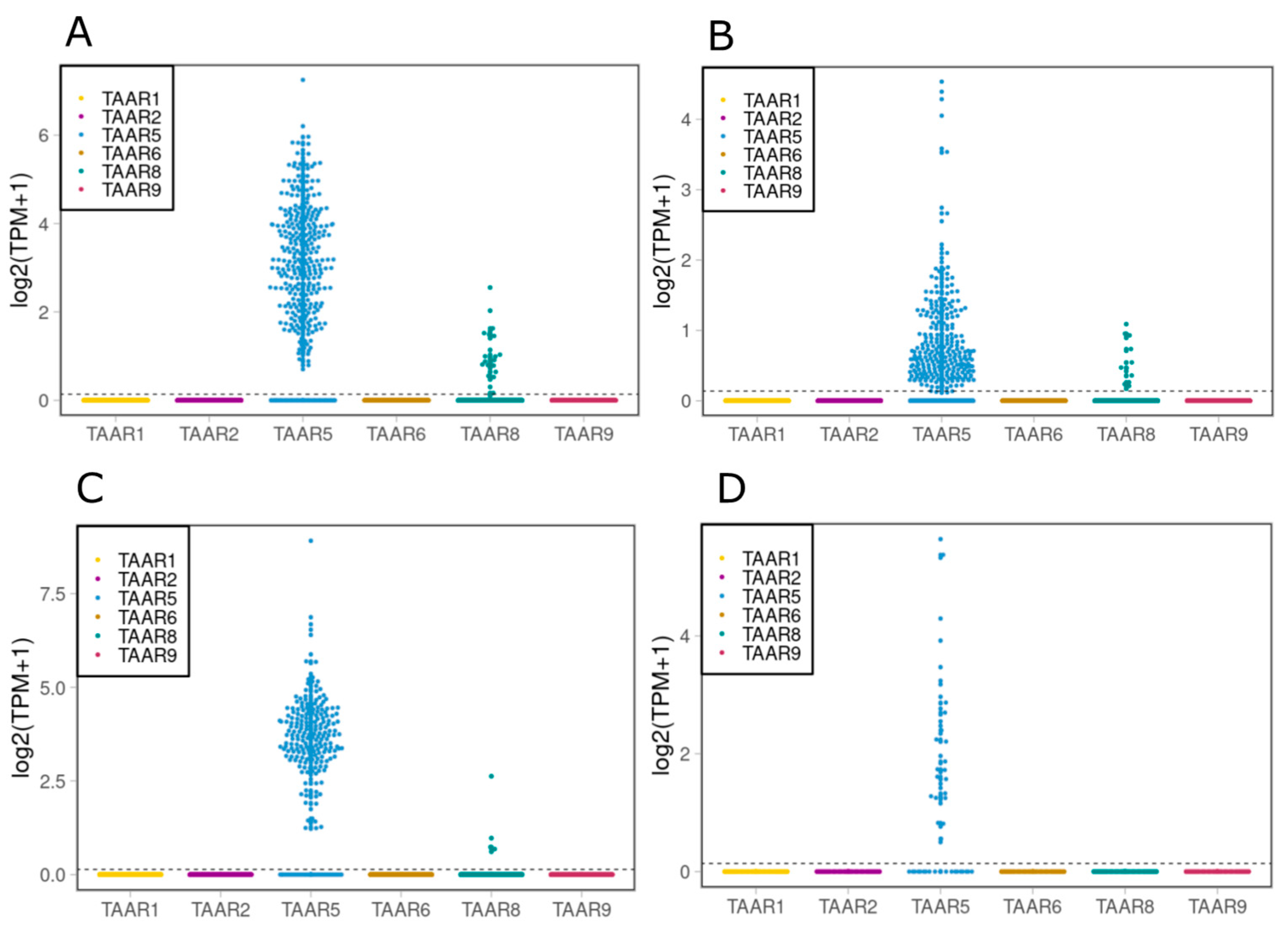
2.2. TAAR5, TAAR6 and TAAR8 Are Expressed in Cells, Isolated from Human SN and VTA
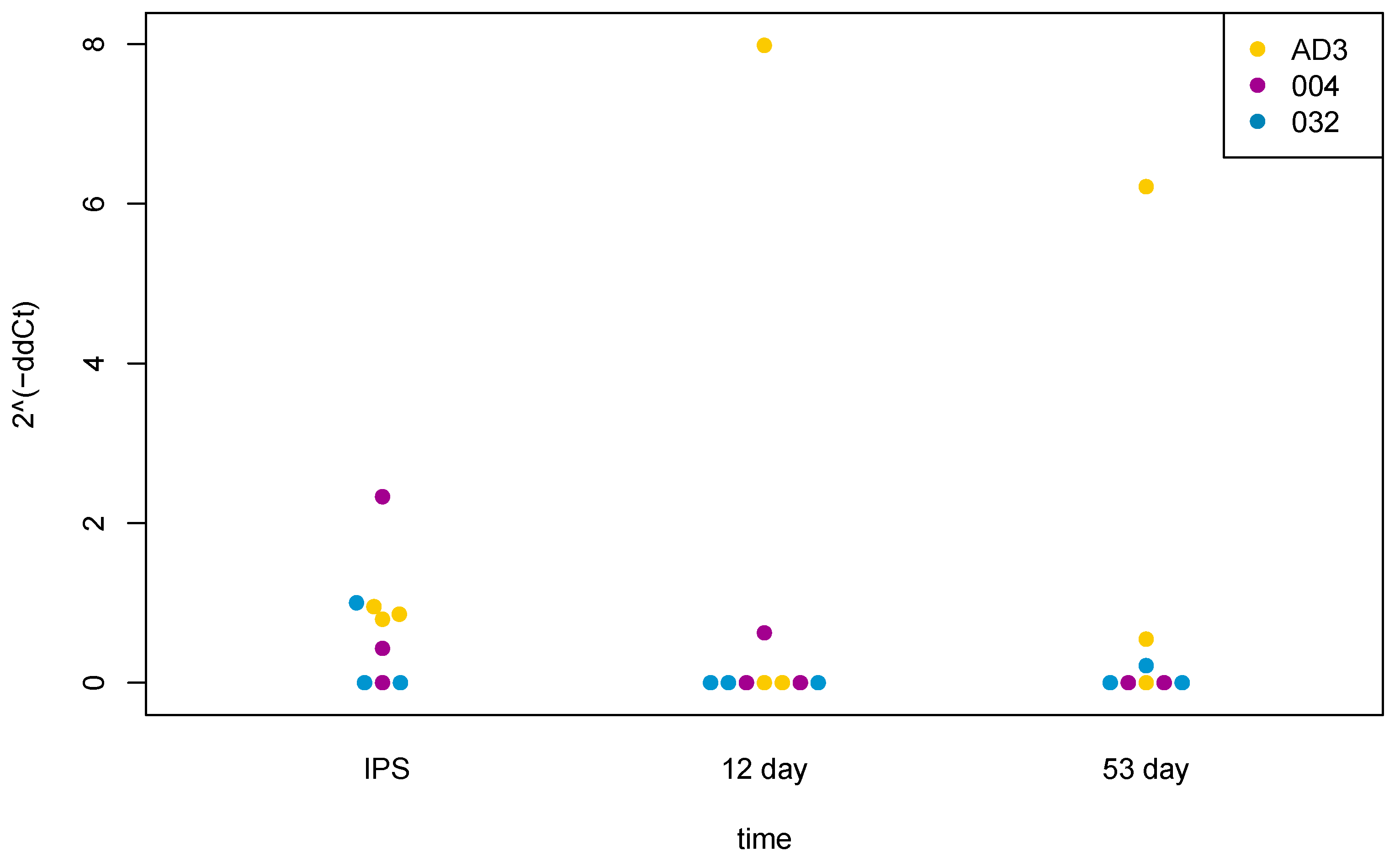
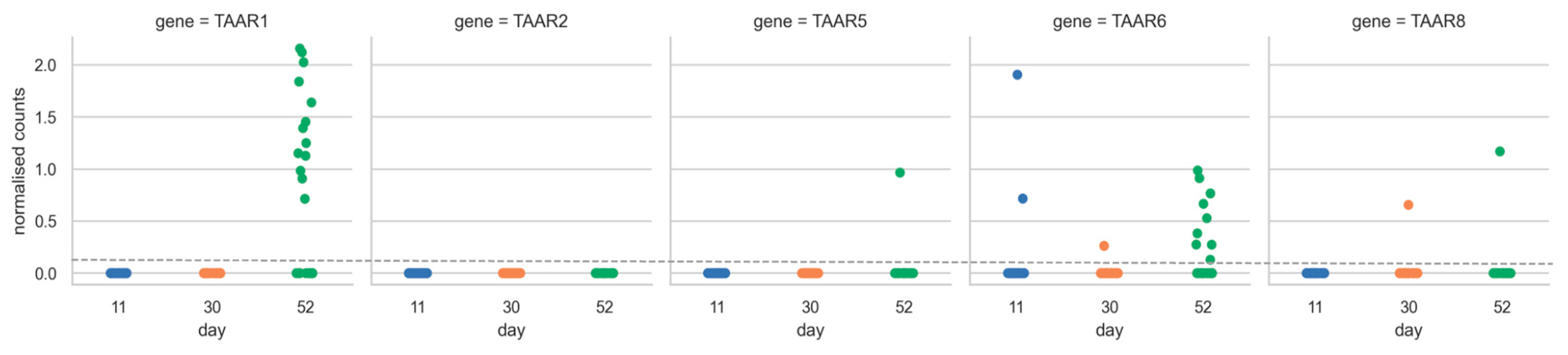
2.3. TAARs’ Profiles of Expression Are Changed during Differentiation of Human Pluripotent Stem Cells to Midbrain Neurons
3. Discussion
4. Materials and Methods
4.1. Public Transcriptomic Data Analysis
4.2. Cell Lines
Human IPSC Lines
- The human IPSC line (AD3) was generated from human newborn fibroblasts (HNFs) using the lentiviral nonintegrating Sendai reprogramming kit (CytoTune-iPS 2.0 Sendai Reprogramming kit (Invitrogen, Paisley, UK) according to the manufacturer’s instructions. HNFs were purchased from Lonza and were cultured as described [47]. The generated human IPSCs were cultured under feeder-free conditions and maintained on plates coated with Matrigel (growth factor reduced; BD, USA) with mTeSR1 (STEMCELL) at 37 °C, 5% CO2 and 21% O2 according to WiCell Inc. protocols. Cells were passaged every 4–5 days at ∼80% confluence by using 0.02% EDTA (Versene). A generated AD3 Sendai-derived human IPSC line was characterized according to the protocol published before [48] and fulfilled all pluripotency criteria [47];
- WTSIi004-A (HPSI1113i-qolg_3) from the European Bank for Induced pluripotent Stem Cells (EBiSC), Biosample ID SAMEA2464810
- WTSIi032-A (HPSI1113i-bima_1) from EBiSC, Biosamples ID SAMEA2399246
4.3. Differentiation of Human IPSCs
4.4. RNA Isolation and Reverse Transcription
4.5. TaqMan Real-Time PCR
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TA | Trace Amines |
| TAARs | Trace Amine-Associated Receptors |
| SN | Substantia Nigra |
| PCNA | Proliferating Cell Nuclear Antigen |
| SVZ | Subventricular Zone |
| SGZ | Subgranular Zone |
| DG | Dentate Gyrus |
| BDNF | Brain-Derived Neurotrophic Factor |
| MAO-B | Monoamine Oxidase B |
| DOPAC | 3,4-Dihydoxyphenylacetic Acid |
| HVA | Homovanillic Acid |
| GDNF | Glial Cell Line-Derived Neurotrophic Factor |
| DA | Dopamine |
| IPSC | Induced Pluripotent Stem Cell |
| fMB | Fetal Ventral Midbrain |
| ESC | Embryonic Stem Cell |
References
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef]
- Berry, M.D. Mammalian Central Nervous System Trace Amines. Pharmacologic Amphetamines, Physiologic Neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace Amines: Identification of a Family of Mammalian G Protein-Coupled Receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef]
- Bunzow, J.R.; Sonders, M.S.; Arttamangkul, S.; Harrison, L.M.; Zhang, G.; Quigley, D.I.; Darland, T.; Suchland, K.L.; Pasumamula, S.; Kennedy, J.L.; et al. Amphetamine, 3,4-Methylenedioxymethamphetamine, Lysergic Acid Diethylamide, and Metabolites of the Catecholamine Neurotransmitters Are Agonists of a Rat Trace Amine Receptor. Mol. Pharmacol. 2001, 60, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, L.; Hoener, M.C. A Renaissance in Trace Amines Inspired by a Novel GPCR Family. Trends Pharmacol. Sci. 2005, 26, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, L.; Meyer, C.A.; Jeanneau, K.; Bradaia, A.; Ozmen, L.; Bluethmann, H.; Bettler, B.; Wettstein, J.G.; Borroni, E.; Moreau, J.-L.; et al. Trace Amine-Associated Receptor 1 Modulates Dopaminergic Activity. J. Pharmacol. Exp. Ther. 2008, 324, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Liberles, S.D.; Buck, L.B. A Second Class of Chemosensory Receptors in the Olfactory Epithelium. Nature 2006, 442, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Wolinsky, T.D.; Swanson, C.J.; Smith, K.E.; Zhong, H.; Borowsky, B.; Seeman, P.; Branchek, T.; Gerald, C.P. The Trace Amine 1 Receptor Knockout Mouse: An Animal Model with Relevance to Schizophrenia. Genes Brain Behav. 2007, 6, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Bradaia, A.; Trube, G.; Stalder, H.; Norcross, R.D.; Ozmen, L.; Wettstein, J.G.; Pinard, A.; Buchy, D.; Gassmann, M.; Hoener, M.C.; et al. The Selective Antagonist EPPTB Reveals TAAR1-Mediated Regulatory Mechanisms in Dopaminergic Neurons of the Mesolimbic System. Proc. Natl. Acad. Sci. USA 2009, 106, 20081–20086. [Google Scholar] [CrossRef]
- Rutigliano, G.; Zucchi, R. Molecular Variants in Human Trace Amine-Associated Receptors and Their Implications in Mental and Metabolic Disorders. Cell. Mol. Neurobiol. 2020, 40, 239–255. [Google Scholar] [CrossRef]
- Revel, F.G.; Moreau, J.-L.; Pouzet, B.; Mory, R.; Bradaia, A.; Buchy, D.; Metzler, V.; Chaboz, S.; Groebke Zbinden, K.; Galley, G.; et al. A New Perspective for Schizophrenia: TAAR1 Agonists Reveal Antipsychotic- and Antidepressant-like Activity, Improve Cognition and Control Body Weight. Mol. Psychiatry 2013, 18, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Martinez, M.; Zhang, J.; Sanders, A.R.; Badner, J.A.; Cravchik, A.; Markey, C.J.; Beshah, E.; Guroff, J.J.; Maxwell, M.E.; et al. Suggestive Evidence for a Schizophrenia Susceptibility Locus on Chromosome 6q and a Confirmation in an Independent Series of Pedigrees. Genomics 1997, 43, 1–8. [Google Scholar] [CrossRef]
- Pei, Y.; Asif-Malik, A.; Canales, J.J. Trace Amines and the Trace Amine-Associated Receptor 1: Pharmacology, Neurochemistry, and Clinical Implications. Front. Neurosci. 2016, 10, 148. [Google Scholar] [CrossRef]
- Ferragud, A.; Howell, A.D.; Moore, C.F.; Ta, T.L.; Hoener, M.C.; Sabino, V.; Cottone, P. The Trace Amine-Associated Receptor 1 Agonist RO5256390 Blocks Compulsive, Binge-like Eating in Rats. Neuropsychopharmacology 2017, 42, 1458–1470. [Google Scholar] [CrossRef]
- Liu, J.-F.; Seaman, R.; Siemian, J.N.; Bhimani, R.; Johnson, B.; Zhang, Y.; Zhu, Q.; Hoener, M.C.; Park, J.; Dietz, D.M.; et al. Role of Trace Amine-Associated Receptor 1 in Nicotine’s Behavioral and Neurochemical Effects. Neuropsychopharmacology 2018, 43, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Sukhanov, I.; Efimova, E.V.; Kozlova, A.; Antonova, K.A.; Illiano, P.; Leo, D.; Merkulyeva, N.; Kalinina, D.; Musienko, P.; et al. Trace Amine-Associated Receptor 5 Provides Olfactory Input Into Limbic Brain Areas and Modulates Emotional Behaviors and Serotonin Transmission. Front. Mol. Neurosci. 2020, 13, 18. [Google Scholar] [CrossRef]
- Gozal, E.A.; O’Neill, B.E.; Sawchuk, M.A.; Zhu, H.; Halder, M.; Chou, C.-C.; Hochman, S. Anatomical and Functional Evidence for Trace Amines as Unique Modulators of Locomotor Function in the Mammalian Spinal Cord. Front. Neural Circuits 2014, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Dedic, N.; Dworak, H.; Zeni, C.; Rutigliano, G.; Howes, O.D. Therapeutic Potential of TAAR1 Agonists in Schizophrenia: Evidence from Preclinical Models and Clinical Studies. Int. J. Mol. Sci. 2021, 22, 13185. [Google Scholar] [CrossRef]
- Cripps, M.J.; Bagnati, M.; Jones, T.A.; Ogunkolade, B.W.; Sayers, S.R.; Caton, P.W.; Hanna, K.; Billacura, M.P.; Fair, K.; Nelson, C.; et al. Identification of a Subset of Trace Amine-Associated Receptors and Ligands as Potential Modulators of Insulin Secretion. Biochem. Pharmacol. 2020, 171, 113685. [Google Scholar] [CrossRef] [PubMed]
- Raab, S.; Wang, H.; Uhles, S.; Cole, N.; Alvarez-Sanchez, R.; Künnecke, B.; Ullmer, C.; Matile, H.; Bedoucha, M.; Norcross, R.D.; et al. Incretin-like Effects of Small Molecule Trace Amine-Associated Receptor 1 Agonists. Mol. Metab. 2016, 5, 47–56. [Google Scholar] [CrossRef]
- Regard, J.B.; Kataoka, H.; Cano, D.A.; Camerer, E.; Yin, L.; Zheng, Y.-W.; Scanlan, T.S.; Hebrok, M.; Coughlin, S.R. Probing Cell Type–Specific Functions of Gi in Vivo Identifies GPCR Regulators of Insulin Secretion. J. Clin. Investig. 2007, 117, JCI32994. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Kuvarzin, S.R.; Mor, M.S.; Katolikova, N.V.; Shemiakova, T.S.; Razenkova, V.; Ptukha, M.; Kozlova, A.A.; Murtazina, R.Z.; Smirnova, D.; et al. Trace Amine-Associated Receptor 2 Is Expressed in the Limbic Brain Areas and Is Involved in Dopamine Regulation and Adult Neurogenesis. Front. Behav. Neurosci. 2022, 16, 847410. [Google Scholar] [CrossRef]
- Efimova, E.V.; Kozlova, A.A.; Razenkova, V.; Katolikova, N.V.; Antonova, K.A.; Sotnikova, T.D.; Merkulyeva, N.S.; Veshchitskii, A.S.; Kalinina, D.S.; Korzhevskii, D.E.; et al. Increased Dopamine Transmission and Adult Neurogenesis in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. Neuropharmacology 2021, 182, 108373. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Katolikova, N.V.; Kanov, E.V.; Gainetdinov, R.R. Trace Amine-Associated Receptors at the Cross-Road between Innate Olfaction of Amines, Emotions, and Adult Neurogenesis. Neural Regen. Res. 2022, 17, 1257–1258. [Google Scholar]
- Katolikova, N.V.; Vaganova, A.N.; Efimova, E.V.; Gainetdinov, R.R. Expression of Trace Amine-Associated Receptors in the Murine and Human Hippocampus Based on Public Transcriptomic Data. Cells 2022, 11, 1813. [Google Scholar] [CrossRef]
- Dang, J.; Tiwari, S.K.; Lichinchi, G.; Qin, Y.; Patil, V.S.; Eroshkin, A.M.; Rana, T.M. Zika Virus Depletes Neural Progenitors in Human Cerebral Organoids through Activation of the Innate Immune Receptor TLR3. Cell Stem Cell 2016, 19, 258–265. [Google Scholar] [CrossRef]
- Fatima, M.; Ahmad, M.H.; Srivastav, S.; Rizvi, M.A.; Mondal, A.C. A Selective D2 Dopamine Receptor Agonist Alleviates Depression through Up-Regulation of Tyrosine Hydroxylase and Increased Neurogenesis in Hippocampus of the Prenatally Stressed Rats. Neurochem. Int. 2020, 136, 104730. [Google Scholar] [CrossRef]
- Philippi, A.; Heller, S.; Costa, I.G.; Senée, V.; Breunig, M.; Li, Z.; Kwon, G.; Russell, R.; Illing, A.; Lin, Q.; et al. Mutations and Variants of ONECUT1 in Diabetes. Nat. Med. 2021, 27, 1928–1940. [Google Scholar] [CrossRef] [PubMed]
- Kilpinen, H.; Goncalves, A.; Leha, A.; Afzal, V.; Alasoo, K.; Ashford, S.; Bala, S.; Bensaddek, D.; Casale, F.P.; Culley, O.J.; et al. Common Genetic Variation Drives Molecular Heterogeneity in Human IPSCs. Nature 2017, 546, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Katolikova, N.V.; Khudiakov, A.A.; Shafranskaya, D.D.; Prjibelski, A.D.; Masharskiy, A.E.; Mor, M.S.; Golovkin, A.S.; Zaytseva, A.K.; Neganova, I.E.; Efimova, E.V.; et al. Modulation of Notch Signaling at Early Stages of Differentiation of Human Induced Pluripotent Stem Cells to Dopaminergic Neurons. Int. J. Mol. Sci. 2023, 24, 1429. [Google Scholar] [CrossRef]
- Monzón-Sandoval, J.; Poggiolini, I.; Ilmer, T.; Wade-Martins, R.; Webber, C.; Parkkinen, L. Human-Specific Transcriptome of Ventral and Dorsal Midbrain Dopamine Neurons. Ann. Neurol. 2020, 87, 853–868. [Google Scholar] [CrossRef]
- Aguila, J.; Cheng, S.; Kee, N.; Cao, M.; Wang, M.; Deng, Q.; Hedlund, E. Spatial RNA Sequencing Identifies Robust Markers of Vulnerable and Resistant Human Midbrain Dopamine Neurons and Their Expression in Parkinson’s Disease. Front. Mol. Neurosci. 2021, 14, 699562. [Google Scholar] [CrossRef]
- Jerber, J.; Seaton, D.D.; Cuomo, A.S.E.; Kumasaka, N.; Haldane, J.; Steer, J.; Patel, M.; Pearce, D.; Andersson, M.; Bonder, M.J.; et al. Population-Scale Single-Cell RNA-Seq Profiling across Dopaminergic Neuron Differentiation. Nat. Genet. 2021, 53, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Kriks, S.; Shim, J.W.; Piao, J.; Ganat, Y.M.; Wakeman, D.R.; Xie, Z.; Carrillo-Reid, L.; Auyeung, G.; Antonacci, C.; Buch, A.; et al. Dopamine Neurons Derived from Human ES Cells Efficiently Engraft in Animal Models of Parkinson’s Disease. Nature 2011, 480, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Kirkeby, A.; Nolbrant, S.; Tiklova, K.; Heuer, A.; Kee, N.; Cardoso, T.; Ottosson, D.R.; Lelos, M.J.; Rifes, P.; Dunnett, S.B.; et al. Predictive Markers Guide Differentiation to Improve Graft Outcome in Clinical Translation of HESC-Based Therapy for Parkinson’s Disease. Cell Stem Cell 2017, 20, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Nolbrant, S.; Heuer, A.; Parmar, M.; Kirkeby, A. Generation of High-Purity Human Ventral Midbrain Dopaminergic Progenitors for in Vitro Maturation and Intracerebral Transplantation. Nat. Protoc. 2017, 12, 1962–1979. [Google Scholar] [CrossRef]
- Tiklová, K.; Nolbrant, S.; Fiorenzano, A.; Björklund, Å.K.; Sharma, Y.; Heuer, A.; Gillberg, L.; Hoban, D.B.; Cardoso, T.; Adler, A.F.; et al. Single Cell Transcriptomics Identifies Stem Cell-Derived Graft Composition in a Model of Parkinson’s Disease. Nat. Commun. 2020, 11, 2434. [Google Scholar] [CrossRef]
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of Human Trace Amine-Associated Receptors: Therapeutic Opportunities and Challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets--Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Seltmann, S.; Lekschas, F.; Müller, R.; Stachelscheid, H.; Bittner, M.-S.; Zhang, W.; Kidane, L.; Seriola, A.; Veiga, A.; Stacey, G.; et al. HPSCreg—The Human Pluripotent Stem Cell Registry. Nucleic Acids Res. 2016, 44, D757–D763. [Google Scholar] [CrossRef]
- Moreno, P.; Fexova, S.; George, N.; Manning, J.R.; Miao, Z.; Mohammed, S.; Muñoz-Pomer, A.; Fullgrabe, A.; Bi, Y.; Bush, N.; et al. Expression Atlas Update: Gene and Protein Expression in Multiple Species. Nucleic Acids Res. 2022, 50, D129–D140. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: New York, NY, USA, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Everaert, C.; Luypaert, M.; Maag, J.L.V.; Cheng, Q.X.; Dinger, M.E.; Hellemans, J.; Mestdagh, P. Benchmarking of RNA-Sequencing Analysis Workflows Using Whole-Transcriptome RT-QPCR Expression Data. Sci. Rep. 2017, 7, 1559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, B.; Lin, L.-L.; Zhao, S. Evaluation and Comparison of Computational Tools for RNA-Seq Isoform Quantification. BMC Genom. 2017, 18, 583. [Google Scholar] [CrossRef]
- Gu, J.; Dai, J.; Lu, H.; Zhao, H. Comprehensive Analysis of Ubiquitously Expressed Genes in Humans from A Data-Driven Perspective. Genom. Proteom. Bioinform. 2023, 21, 164–176. [Google Scholar] [CrossRef]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) Project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Neganova, I.; Chichagova, V.; Armstrong, L.; Lako, M. A Critical Role for P38MAPK Signalling Pathway during Reprogramming of Human Fibroblasts to IPSCs. Sci. Rep. 2017, 7, 41693. [Google Scholar] [CrossRef] [PubMed]
- Chichagova, V.; Sanchez-Vera, I.; Armstrong, L.; Steel, D.; Lako, M. Generation of Human Induced Pluripotent Stem Cells Using RNA-Based Sendai Virus System and Pluripotency Validation of the Resulting Cell Population. In Patient-Specific Induced Pluripotent Stem Cell Models; Nagy, A., Turksen, K., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; Volume 1353, pp. 285–307. ISBN 978-1-4939-3033-3. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset ID | Title | Group Description | n |
|---|---|---|---|
| GSE80264 [26] | Transcriptome analysis of H9 human ESC-derived cerebral organoids | H9 ESC line | 2 |
| GSE130829 | Effect of cell division on cell differentiation (RNA-Seq H9 cells UD E24 E24HU E24Noc) | H9 ESC line | 2 |
| GSE146701 [27] | Regulation of histone H3 by the ubiquitin-conjugating enzyme UBE2K determines neurogenesis of human ESCs [RNA-Seq] | H9 ESC line | 3 |
| GSE134228 [28] | In vitro pancreas differentiation of human ESCs line Mel1. | Mel1 ESC line | 4 |
| E-ENAD-35 | HipSci Project—RNA-seq of healthy volunteers | Human IPSCs derived from healthy individuals | 191 |
| E-EMTAB-4748 | RNA-seq of coding RNA in human fibroblasts, peripheral blood mononuclear cells (PBMCs) and IPSCs as part of the HipSci project | IPSCs derived from healthy individuals | 187 |
| GSE166024 [31] | Human-specific transcriptome of ventral and dorsal tiers of the SN pars compacta dopamine neurons | Dorsal midbrain dopamine neurons | 7 |
| Ventral midbrain dopamine neurons | 7 | ||
| GSE114918 [32] | RNA-seq of human SNc and VTA midbrain dopamine neurons isolated from post-mortem material of control subjects and Parkinson’s Disease patients using laser capture microdissection | Substantia nigra | 25 |
| Ventral tegmental area | 16 | ||
| GSE86654 [35] | Identifying markers predicting successful graft outcome for clinical translation of human ESC-based cell therapy for Parkinson’s disease | Human ESCs differentiated to ventral midbrain progenitors (high DA group) | 9 |
| Human ESCs differentiated to ventral midbrain progenitors (low DA group) | 6 | ||
| GSE118412 [37] | Single cell transcriptomics identifies stem cell-derived graft composition in a model of Parkinson’s disease | Dopaminergic progenitors at day 16 of differentiation, obtained from RC17human IPSC line | 404 |
| Dopaminergic progenitors at day 16 of differentiation, obtained from RC17 human IPSC line 6 months after transplantation to striatum of 6-OHDA rats | 683 | ||
| Cells from human fetal midbrain | 256 | ||
| Cells from human fetal midbrain 6 months after transplantation to striatum of 6-OHDA rats | 63 | ||
| EGAD00001006157 [33] | Single cell RNA sequencing data from differentiation of 215 cell lines of human IPSCs to midbrain fate, including dopaminergic neurons | Midbrain fate neurons | 42 |
| Gene | Type | Sequence | Location |
|---|---|---|---|
| TAAR1 | Forward | tgaccacactcgttggcaatctg | 283–305 |
| Reverse | acagtgctcagcagatctcacca | 421–443 | |
| Probe | FAM-ggccactgtggactttcttctggggt-BHQ1 | 374–399 | |
| TAAR2 | Forward | ggtccctggagcatttgccttc | 507–528 |
| Reverse | gccataaacaaggtggtccccc | 626–647 | |
| Probe | FAM-tggttgcttgttccagttcctgccca | 581–606 | |
| TAAR5 | Forward | agcaccattcgctcagtggaga | 323–341 |
| Reverse | gtgaggcagaagagggtgtcca | 369–388 | |
| Probe | FAM-ttcctctgccgcctgcacacct | 364–386 | |
| TAAR6 | Forward | atgtacagcggtgctgtgtt | 391–411 |
| Reverse | caacggtctgacaacctcct | 561–580 | |
| Probe | FAM-ggctggaggaattatctgatgcc-BHQ1 | 527–549 | |
| TAAR8 | Forward | acacaggtgtcaatgatgatggg | 512–534 |
| Reverse | atttgacagccacctacgca | 562–581 | |
| Probe | ctggaggaattagtaagtgctctc | 535–558 | |
| TAAR9 | Forward | gccaggctccactgaatcaa | 611–630 |
| Reverse | agcttggctggctgtacttt | 697–716 | |
| Probe | FAM-ggtggccaagcatcaggctaggaa | 666–689 | |
| HPRT | Forward | ggctccgttatggcgacc | 139–156 |
| Reverse | tcgagcaagacgttcagtcc | 281–300 | |
| Probe | FAM-cagccctggcgtcgtgattagtg | 159–181 | |
| RPS18 | Forward | tcaacaccaacatcgatgggcg | 92–113 |
| Reverse | gctttcctcaacaccacatgagca | 165–188 | |
| Probe | FAM-actgccattaagggtgtgggccga | 136–159 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katolikova, N.V.; Vaganova, A.N.; Shafranskaya, D.D.; Efimova, E.V.; Malashicheva, A.B.; Gainetdinov, R.R. Expression Pattern of Trace Amine-Associated Receptors during Differentiation of Human Pluripotent Stem Cells to Dopaminergic Neurons. Int. J. Mol. Sci. 2023, 24, 15313. https://doi.org/10.3390/ijms242015313
Katolikova NV, Vaganova AN, Shafranskaya DD, Efimova EV, Malashicheva AB, Gainetdinov RR. Expression Pattern of Trace Amine-Associated Receptors during Differentiation of Human Pluripotent Stem Cells to Dopaminergic Neurons. International Journal of Molecular Sciences. 2023; 24(20):15313. https://doi.org/10.3390/ijms242015313
Chicago/Turabian StyleKatolikova, Nataliia V., Anastasia N. Vaganova, Daria D. Shafranskaya, Evgeniya V. Efimova, Anna B. Malashicheva, and Raul R. Gainetdinov. 2023. "Expression Pattern of Trace Amine-Associated Receptors during Differentiation of Human Pluripotent Stem Cells to Dopaminergic Neurons" International Journal of Molecular Sciences 24, no. 20: 15313. https://doi.org/10.3390/ijms242015313