
Retroviral Replicating Vectors Mediated Prodrug Activator Gene Therapy in a Gastric Cancer Model

 , and
, and 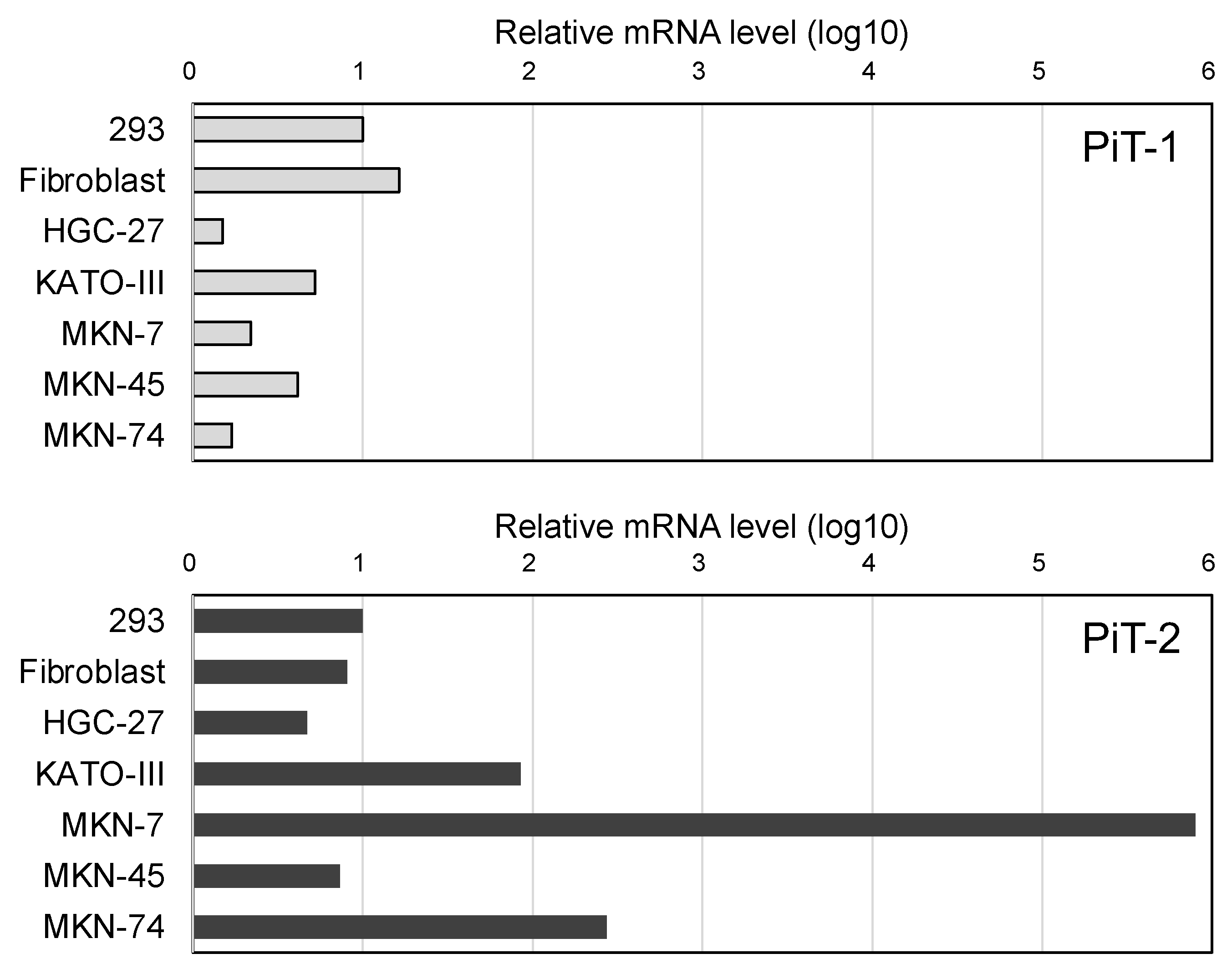{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Relative Evaluation of PiT-2 or PiT-1 Expression Levels
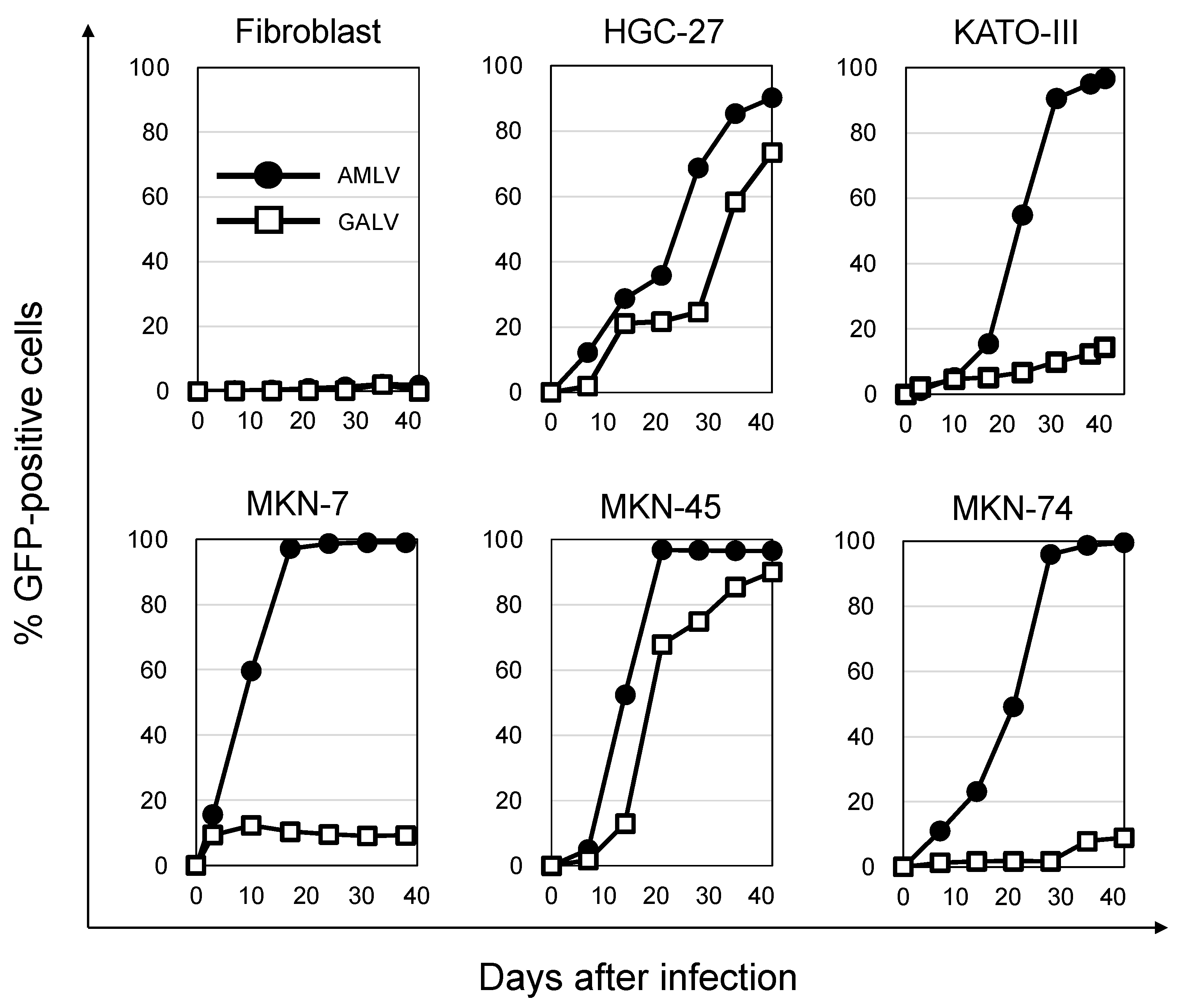
2.2. AMLV Vectors Replicate Efficiently in Human GC Cell Lines In Vitro
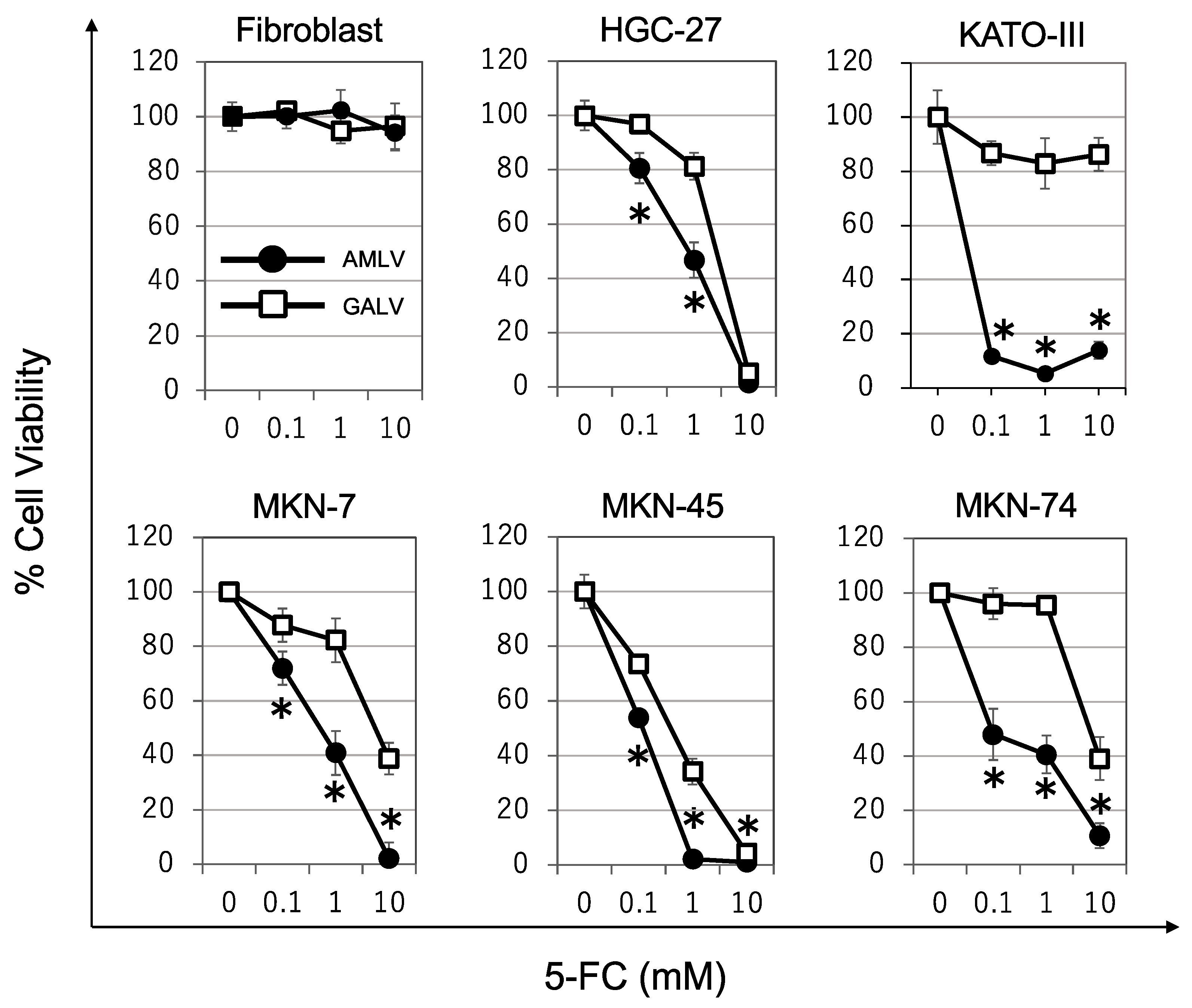
2.3. Prodrug Activator Gene-Mediated Cell-Killing Effect of RRVs on Human GC Cells
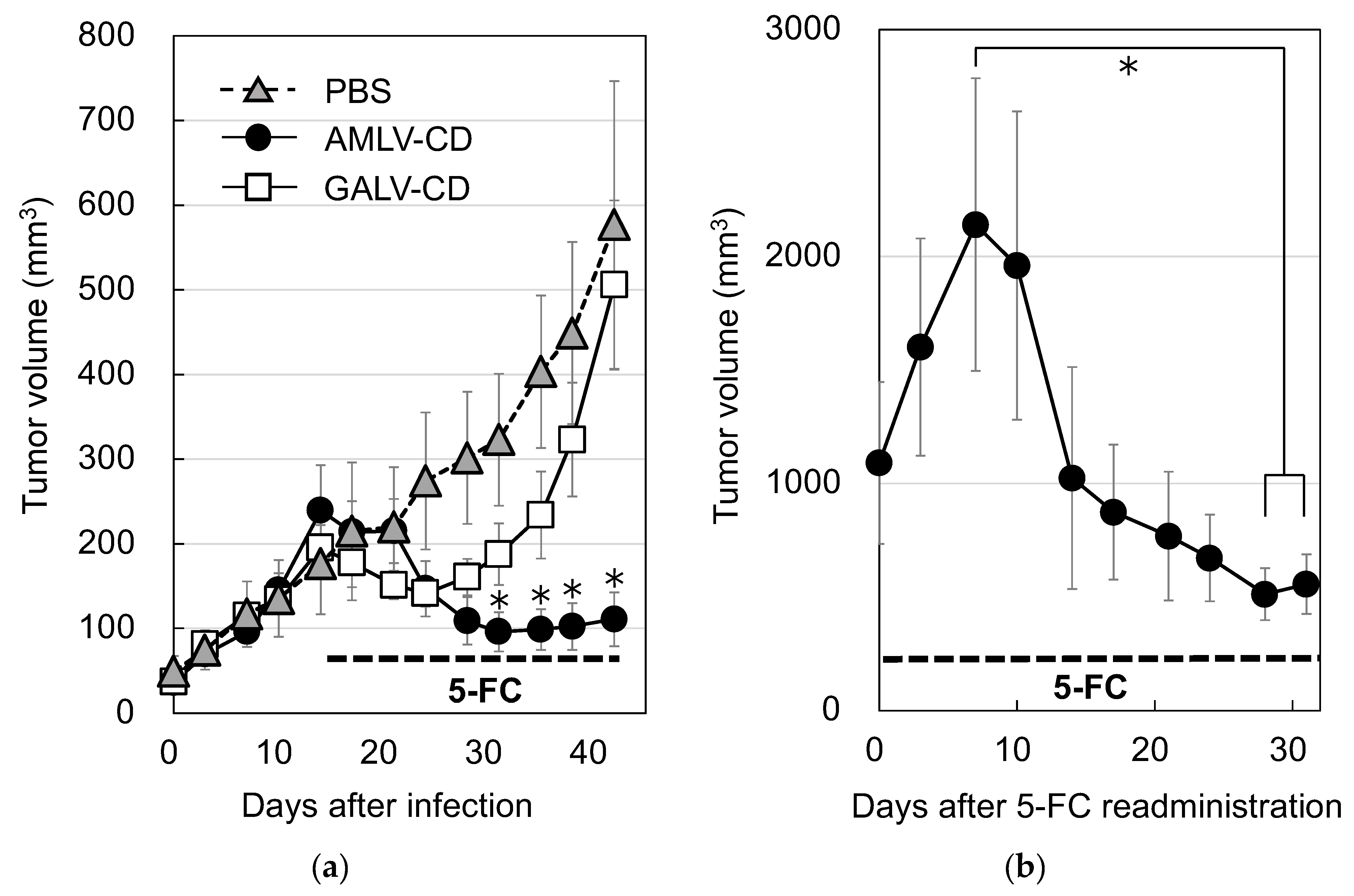
2.4. AMLV-Mediated CD/5-FC Prodrug Activator Gene Therapy Has Potent In Vivo Antitumor Effects in Primary and Recurrent Subcutaneous MKN-74 Human GC Xenograft Models
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmid and RRV Production
4.3. Quantitative Real-Time PCR Analysis
4.4. Flow Cytometry of RRV Replication Kinetics
4.5. AlamarBlue Assay
4.6. Subcutaneous Xenograft Model of Human GC
4.7. Statistical Analysis
4.8. Abbreviations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef]
- Wang, J.; Du, L.; Chen, X. Oncolytic virus: A catalyst for the treatment of gastric cancer. Front. Oncol. 2022, 12, 1017692. [Google Scholar] [CrossRef]
- Biagioni, A.; Skalamera, I.; Peri, S.; Schiavone, N.; Cianchi, F.; Giommoni, E.; Magnelli, L.; Papucci, L. Update on gastric cancer treatments and gene therapies. Cancer Metastasis Rev. 2019, 38, 537–548. [Google Scholar] [CrossRef]
- Tsuji, T.; Nakamori, M.; Iwahashi, M.; Nakamura, M.; Ojima, T.; Iida, T.; Katsuda, M.; Hayata, K.; Ino, Y.; Todo, T.; et al. An armed oncolytic herpes simplex virus expressing thrombospondin-1 has an enhanced in vivo antitumor effect against human gastric cancer. Int. J. Cancer 2013, 132, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Dai, S.; Zhu, H.; Song, Z.; Cai, Y.; Lee, J.B.; Li, Z.; Hu, X.; Fang, B.; He, C.; et al. Telomerase-specific oncolytic adenovirus expressing TRAIL suppresses peritoneal dissemination of gastric cancer. Gene Ther. 2017, 24, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Haley, E.S.; Au, G.G.; Carlton, B.R.; Barry, R.D.; Shafren, D.R. Regional administration of oncolytic Echovirus 1 as a novel therapy for the peritoneal dissemination of gastric cancer. J. Mol. Med. 2009, 87, 385–399. [Google Scholar] [CrossRef]
- Song, K.Y.; Wong, J.; Gonzalez, L.; Sheng, G.; Zamarin, D.; Fong, Y. Antitumor efficacy of viral therapy using genetically engineered Newcastle disease virus [NDV(F3aa)-GFP] for peritoneally disseminated gastric cancer. J. Mol. Med. 2010, 88, 589–596. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Etoh, T.; Suzuki, K.; Mitui, M.T.; Nishizono, A.; Shiraishi, N.; Kitano, S. Efficacy of oncolytic reovirus against human gastric cancer with peritoneal metastasis in experimental animal model. Int. J. Oncol. 2010, 37, 1433–1438. [Google Scholar] [CrossRef]
- Jun, K.-H.; Gholami, S.; Song, T.-J.; Au, J.; Haddad, D.; Carson, J.; Chen, C.-H.; Mojica, K.; Zanzonico, P.; Chen, N.G.; et al. A novel oncolytic viral therapy and imaging technique for gastric cancer using a genetically engineered vaccinia virus carrying the human sodium iodide symporter. J. Exp. Clin. Cancer Res. 2014, 33, 2. [Google Scholar] [CrossRef]
- Collins, S.A.; Shah, A.H.; Ostertag, D.; Kasahara, N.; Jolly, D.J. Clinical development of retroviral replicating vector Toca 511 for gene therapy of cancer. Expert Opin. Biol. Ther. 2021, 21, 1199–1214. [Google Scholar] [CrossRef] [PubMed]
- Douville, R.N.; Hiscott, J. The interface between the innate interferon response and expression of host retroviral restriction factors. Cytokine 2010, 52, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C. Cell-to-cell transmission of retroviruses: Innate immunity and interferon-induced restriction factors. Virology 2011, 411, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Stavrou, S.; Aguilera, A.N.; Blouch, K.; Ross, S.R. DDX41 Recognizes RNA/DNA Retroviral Reverse Transcripts and Is Critical for In Vivo Control of Murine Leukemia Virus Infection. mBio 2018, 9, 00923-18. [Google Scholar] [CrossRef]
- Takeuchi, H.; Matano, T. Host factors involved in resistance to retroviral infection. Microbiol. Immunol. 2008, 52, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Biasi, G.; Facchinetti, A.; Cappellari, R.; Rossi, E.; Zanovello, P. Immune response to Moloney-murine leukemia virus-induced antigens in bone marrow. Immunol. Lett. 2011, 138, 79–85. [Google Scholar] [CrossRef]
- Hein, A.; Czub, S.; Xiao, L.X.; Schwender, S.; Dörries, R.; Czub, M. Effects of Adoptive Immune Transfers on Murine Leukemia Virus-Infection of Rats. Virology 1995, 211, 408–417. [Google Scholar] [CrossRef]
- Littwitz-Salomon, E.; Schimmer, S.; Dittmer, U. Natural killer T cells contribute to the control of acute retroviral infection. Retrovirology 2017, 14, 5. [Google Scholar] [CrossRef]
- Kievit, E.; Bershad, E.; Ng, E.; Sethna, P.; Dev, I.; Lawrence, T.S.; Rehemtulla, A. Superiority of yeast over bacterial cytosine deaminase for enzyme/prodrug gene therapy in colon cancer xenografts. Cancer Res. 1999, 59, 1417–1421. [Google Scholar]
- Nyati, M.K.; Symon, Z.; Kievit, E.; Dornfeld, K.J.; Rynkiewicz, S.D.; Ross, B.D.; Rehemtulla, A.; Lawrence, T.S. The potential of 5-fluorocytosine/cytosine deaminase enzyme prodrug gene therapy in an intrahepatic colon cancer model. Gene Ther. 2002, 9, 844–849. [Google Scholar] [CrossRef]
- Tai, C.-K.; Wang, W.J.; Chen, T.C.; Kasahara, N. Single-Shot, Multicycle Suicide Gene Therapy by Replication-Competent Retrovirus Vectors Achieves Long-Term Survival Benefit in Experimental Glioma. Mol. Ther. 2005, 12, 842–851. [Google Scholar] [CrossRef]
- Kubo, S.; Takagi-Kimura, M.; Logg, C.R.; Kasahara, N. Highly efficient tumor transduction and antitumor efficacy in experimental human malignant mesothelioma using replicating gibbon ape leukemia virus. Cancer Gene Ther. 2013, 20, 671–677. [Google Scholar] [CrossRef]
- Kushiya, H.; Hiraoka, K.; Suzuki, T.; Inoko, K.; Inagaki, A.; Niwa, H.; Sasaki, K.; Nakamura, T.; Tsuchikawa, T.; Shichinohe, T.; et al. Retroviral Replicating Vector Toca 511 (Vocimagene Amiretrorepvec) for Prodrug Activator Gene Therapy of Lung Cancer. Cancers 2022, 14, 5820. [Google Scholar] [CrossRef]
- Inoko, K.; Hiraoka, K.; Inagaki, A.; Takahashi, M.; Kushibiki, T.; Hontani, K.; Takano, H.; Sato, S.; Takeuchi, S.; Nakamura, T.; et al. Therapeutic activity of retroviral replicating vector-mediated prodrug activator gene therapy for pancreatic cancer. Cancer Gene Ther. 2018, 25, 184–195. [Google Scholar] [CrossRef]
- Kubo, S.; Takagi-Kimura, M.; Kasahara, N. Efficient tumor transduction and antitumor efficacy in experimental human osteosarcoma using retroviral replicating vectors. Cancer Gene Ther. 2019, 26, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Perez, O.D.; Logg, C.R.; Hiraoka, K.; Diago, O.; Burnett, R.; Inagaki, A.; Jolson, D.; Amundson, K.; Buckley, T.; Lohse, D.; et al. Design and Selection of Toca 511 for Clinical Use: Modified Retroviral Replicating Vector with Improved Stability and Gene Expression. Mol. Ther. 2012, 20, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Cloughesy, T.F.; Landolfi, J.; Vogelbaum, M.A.; Ostertag, D.; Elder, J.B.; Bloomfield, S.; Carter, B.; Chen, C.C.; Kalkanis, S.N.; Kesari, S.; et al. Durable complete responses in some recurrent high-grade glioma patients treated with Toca 511 + Toca FC. Neuro-Oncology 2018, 20, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.J.; Zhu, J.-J.; Diago, O.R.; Gammon, D.K.; Haghighi, A.; Lu, G.; Das, A.; Gruber, H.E.; Jolly, D.J.; Ostertag, D. Molecular Analyses Support the Safety and Activity of Retroviral Replicating Vector Toca 511 in Patients. Clin. Cancer Res. 2018, 24, 4680–4693. [Google Scholar] [CrossRef]
- Cloughesy, T.F.; Petrecca, K.; Walbert, T.; Butowski, N.; Salacz, M.; Perry, J.; Damek, D.; Bota, D.; Bettegowda, C.; Zhu, J.-J.; et al. Effect of Vocimagene Amiretrorepvec in Combination with Flucytosine vs Standard of Care on Survival Following Tumor Resection in Patients with Recurrent High-Grade Glioma: A Randomized Clinical Trial. JAMA Oncol. 2020, 6, 1936–1946. [Google Scholar] [CrossRef]
- Liu, M.; Eiden, M.V. The receptors for gibbon ape leukemia virus and amphotropic murine leukemia virus are not downregulated in productively infected cells. Retrovirology 2011, 8, 53. [Google Scholar] [CrossRef]
- Miller, D.G.; Adam, A.M.; Miller, A.D. Gene transfer by retrovirus vectors occurs only in cells that are actively replicating at the time of infection. Mol. Cell. Biol. 1990, 10, 4239–4242. [Google Scholar] [CrossRef] [PubMed]
- Overbaugh, J.; Miller, A.D.; Eiden, M.V. Receptors and Entry Cofactors for Retroviruses Include Single and Multiple Transmembrane-Spanning Proteins as well as Newly Described Glycophosphatidylinositol-Anchored and Secreted Proteins. Microbiol. Mol. Biol. Rev. 2001, 65, 371–389. [Google Scholar] [CrossRef] [PubMed]
- Kubo, S.; Takagi-Kimura, M.; Tagawa, M.; Kasahara, N. Dual-vector prodrug activator gene therapy using retroviral replicating vectors. Cancer Gene Ther. 2019, 26, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Heiber, J.F.; Barber, G.N. Evaluation of Innate Immune Signaling Pathways in Transformed Cells. Methods Mol. Biol. 2012, 797, 217–238. [Google Scholar] [CrossRef]
- Naik, S.; Russell, S.J. Engineering oncolytic viruses to exploit tumor specific defects in innate immune signaling pathways. Expert Opin. Biol. Ther. 2009, 9, 1163–1176. [Google Scholar] [CrossRef]
- Arens, R. Rational Design of Vaccines: Learning from immune evasion mechanisms of persistent viruses and tumors. Adv. Immunol. 2012, 114, 217–243. [Google Scholar] [CrossRef]
- Butt, A.Q.; Mills, K.H.G. Immunosuppressive networks and checkpoints controlling antitumor immunity and their blockade in the development of cancer immunotherapeutics and vaccines. Oncogene 2014, 33, 4623–4631. [Google Scholar] [CrossRef]
- Tung, I.; Sahu, A. The treatment of resectable gastric cancer: A literature review of an evolving landscape. J. Gastrointest. Oncol. 2022, 13, 871–884. [Google Scholar] [CrossRef]
- Mori, R.; Yoshida, K.; Futamura, M.; Suetsugu, T.; Shizu, K.; Tanahashi, T.; Tanaka, Y.; Matsuhashi, N.; Yamaguchi, K. The inhibition of thymidine phosphorylase can reverse acquired 5FU-resistance in gastric cancer cells. Gastric Cancer 2019, 22, 497–505. [Google Scholar] [CrossRef]
- Düzgüneş, N. Origins of Suicide Gene Therapy. Methods Mol. Biol. 2019, 1895, 1–9. [Google Scholar] [CrossRef]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a Human Cell Line Transformed by DNA from Human Adenovirus Type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef] [PubMed]
- DuBridge, R.B.; Tang, P.; Hsia, H.C.; Leong, P.-M.; Miller, J.H.; Calos, M.P. Analysis of Mutation in Human Cells by Using an Epstein-Barr Virus Shuttle System. Mol. Cell. Biol. 1987, 7, 379–387. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujino, H.; Sonoda-Fukuda, E.; Isoda, L.; Kawabe, A.; Takarada, T.; Kasahara, N.; Kubo, S. Retroviral Replicating Vectors Mediated Prodrug Activator Gene Therapy in a Gastric Cancer Model. Int. J. Mol. Sci. 2023, 24, 14823. https://doi.org/10.3390/ijms241914823
Fujino H, Sonoda-Fukuda E, Isoda L, Kawabe A, Takarada T, Kasahara N, Kubo S. Retroviral Replicating Vectors Mediated Prodrug Activator Gene Therapy in a Gastric Cancer Model. International Journal of Molecular Sciences. 2023; 24(19):14823. https://doi.org/10.3390/ijms241914823
Chicago/Turabian StyleFujino, Hiroaki, Emiko Sonoda-Fukuda, Lisa Isoda, Ayane Kawabe, Toru Takarada, Noriyuki Kasahara, and Shuji Kubo. 2023. "Retroviral Replicating Vectors Mediated Prodrug Activator Gene Therapy in a Gastric Cancer Model" International Journal of Molecular Sciences 24, no. 19: 14823. https://doi.org/10.3390/ijms241914823