


Myogenetic Oligodeoxynucleotide Induces Myocardial Differentiation of Murine Pluripotent Stem Cells
 , , ,
, , , 
Abstract
:1. Introduction
2. Results
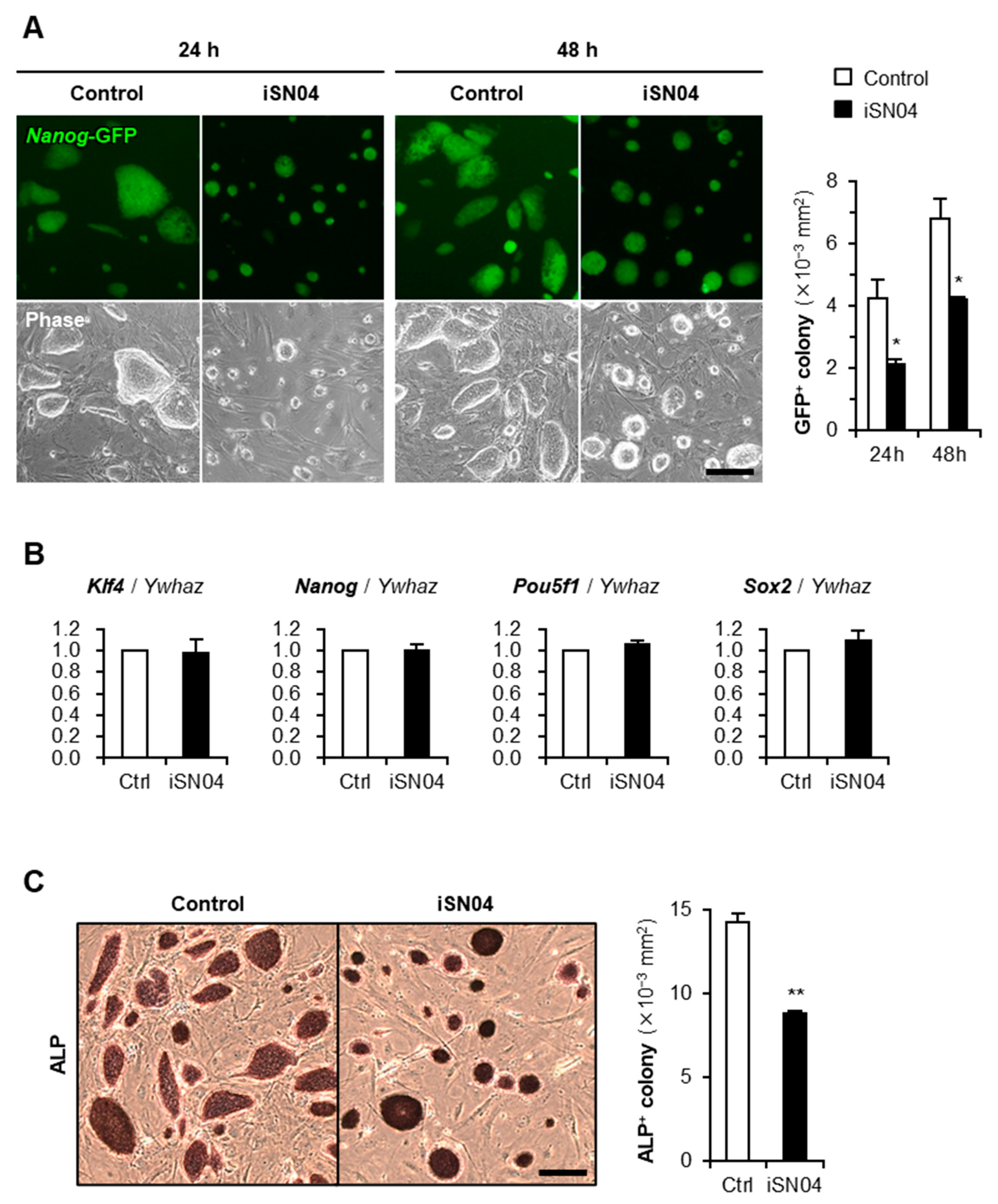
2.1. iSN04 Inhibits Proliferation of Undifferentiated PSCs
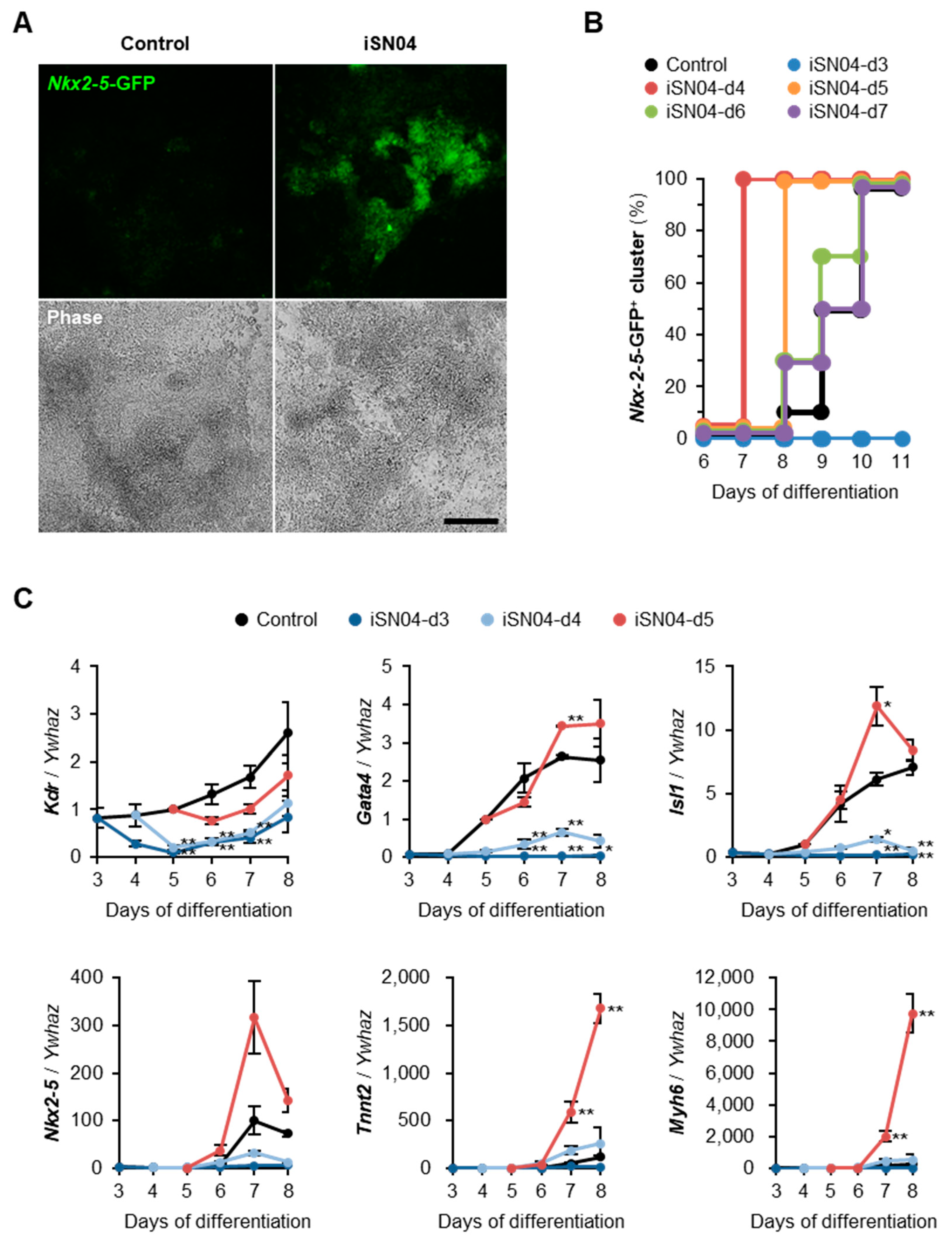
2.2. iSN04 Induces Myocardial Differentiation of PSCs
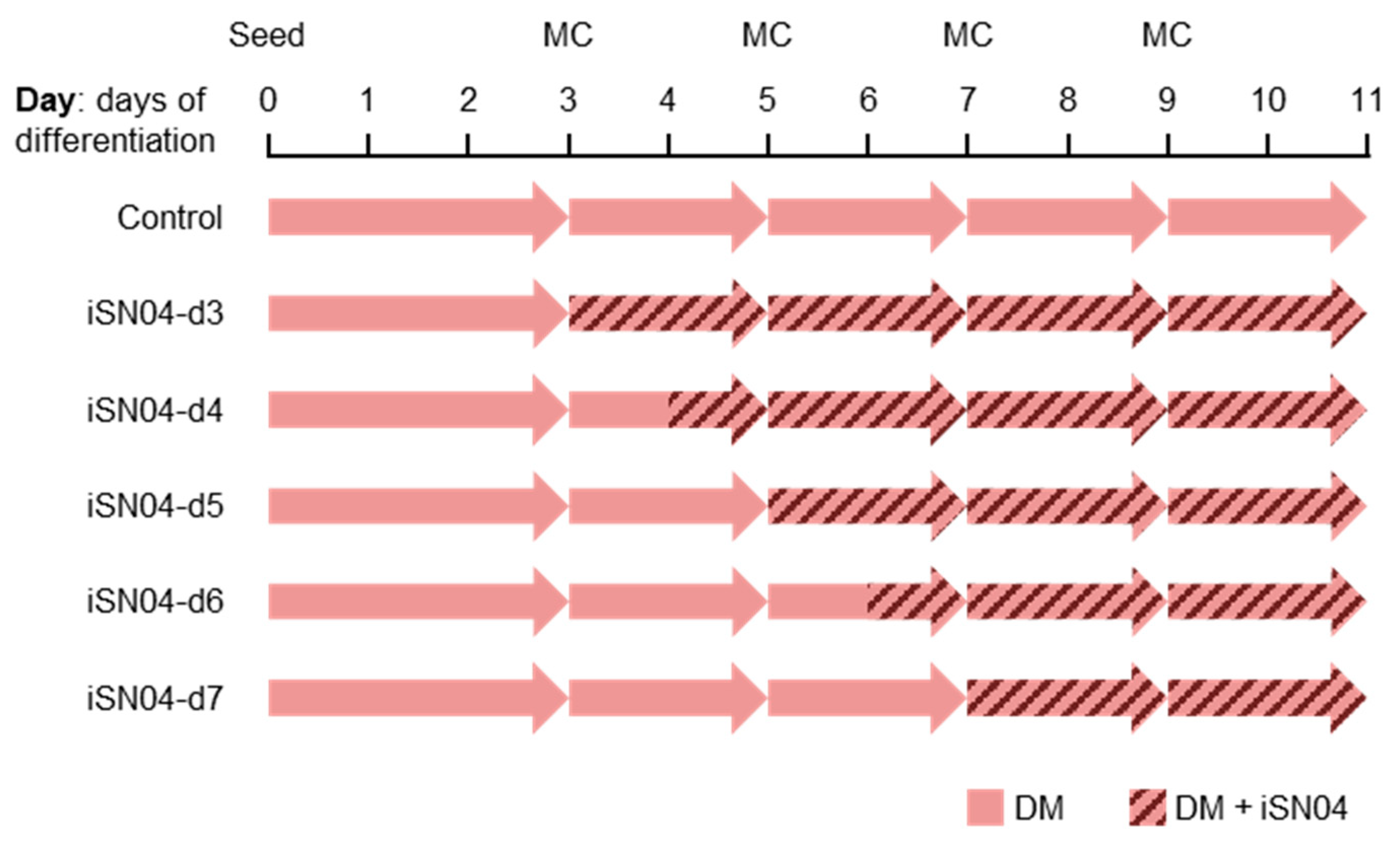
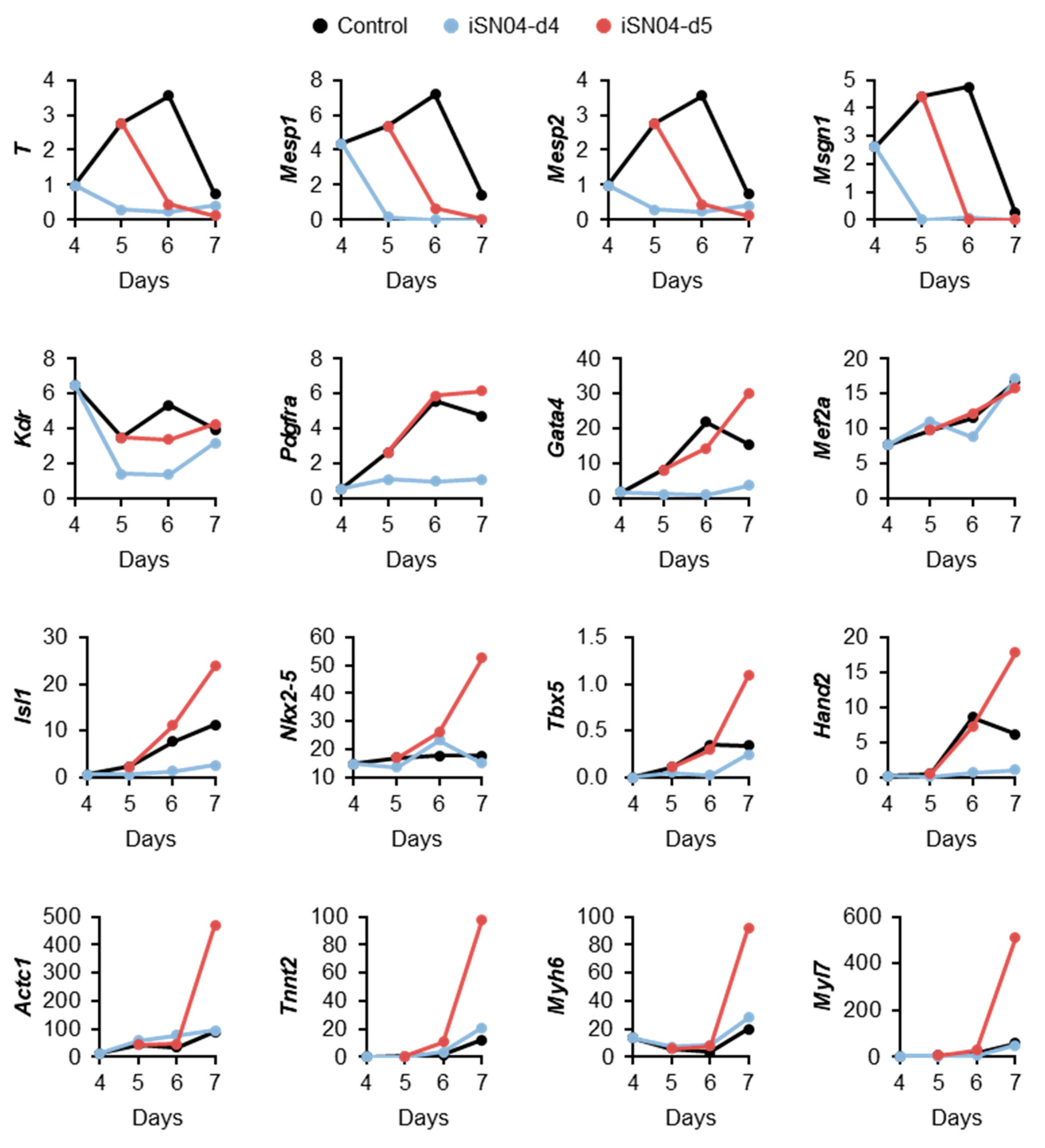
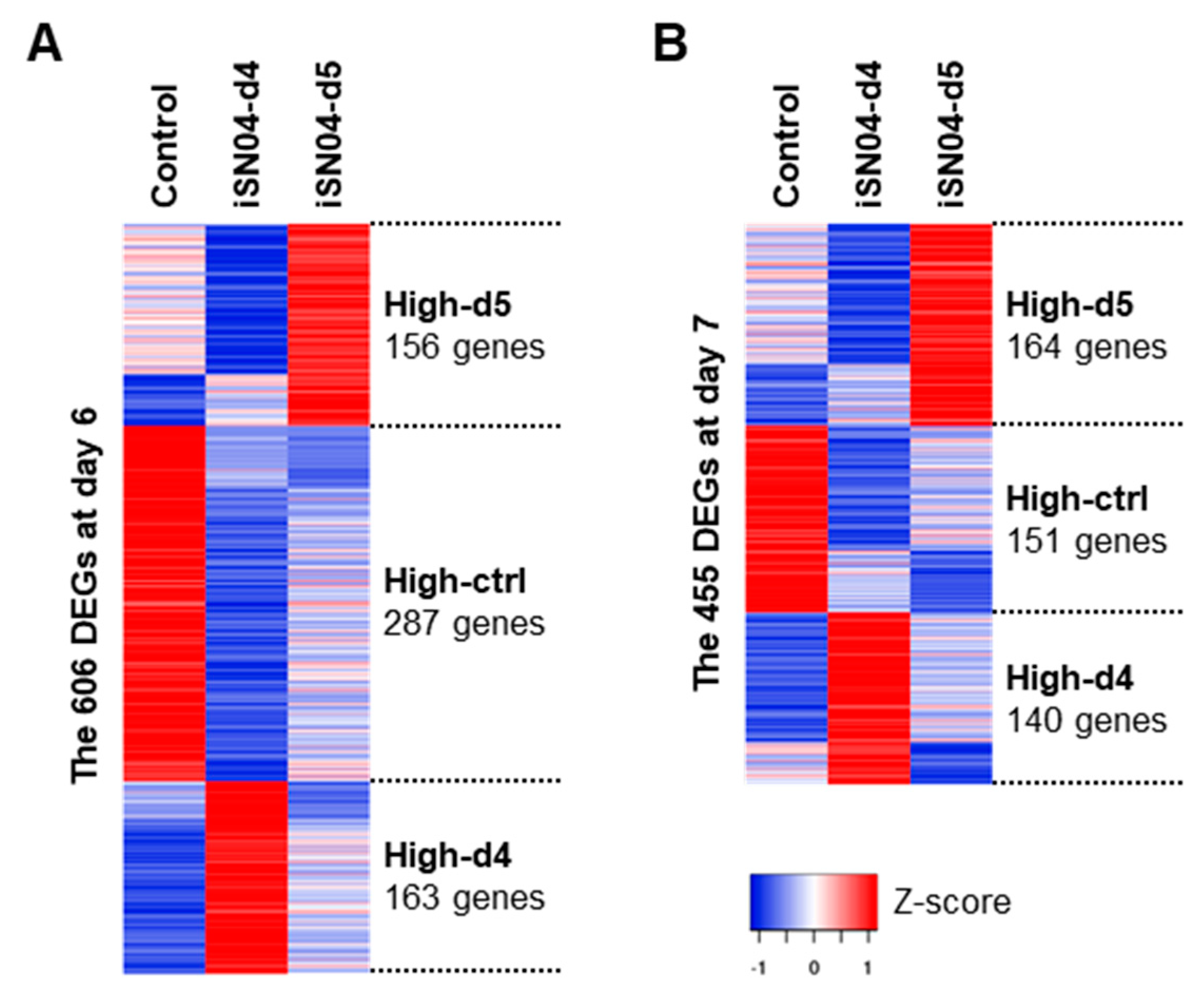
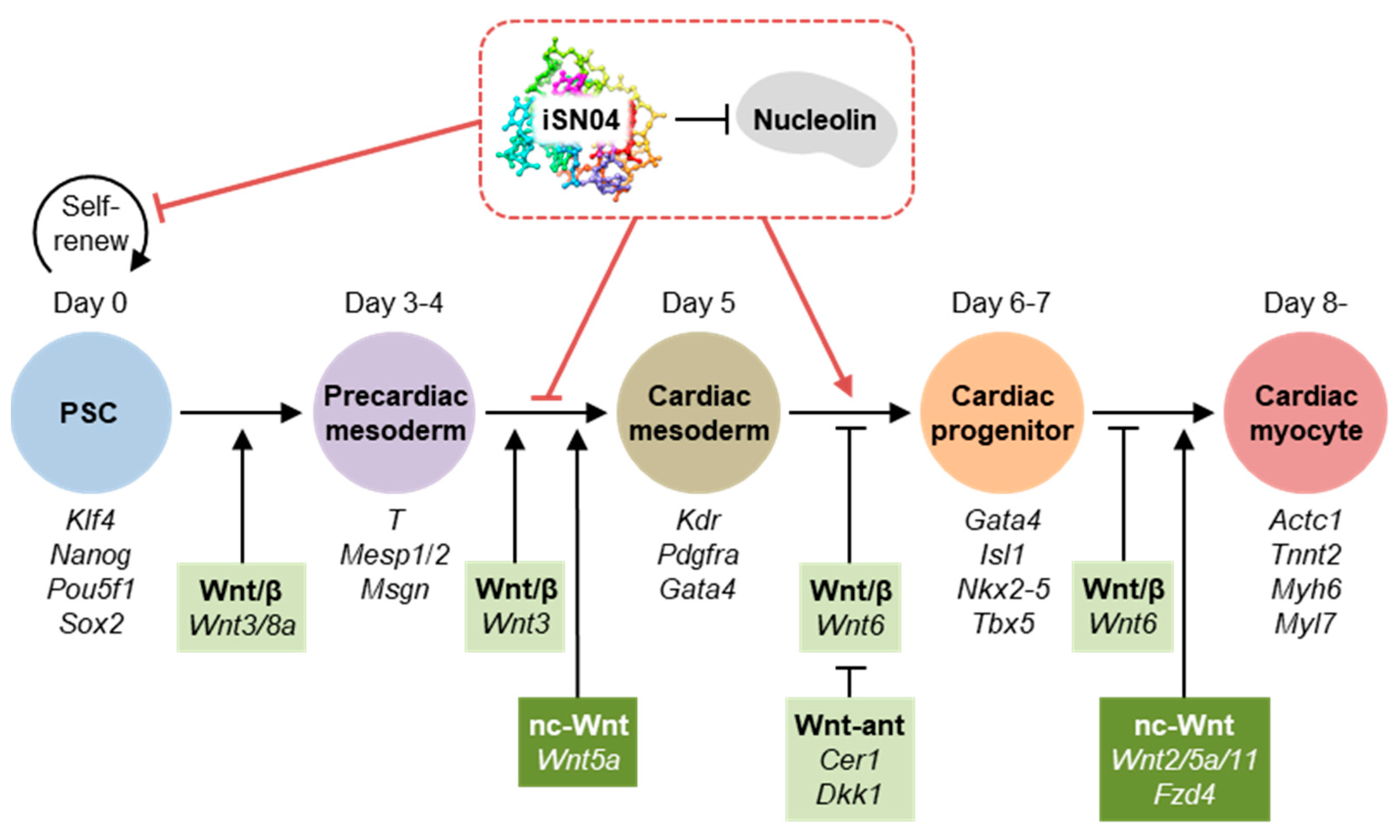
2.3. iSN04 Influences Mesoderm Differentiation into Cardiac Progenitors
2.4. iSN04 Affects the Wnt Signaling Pathway in PSCs
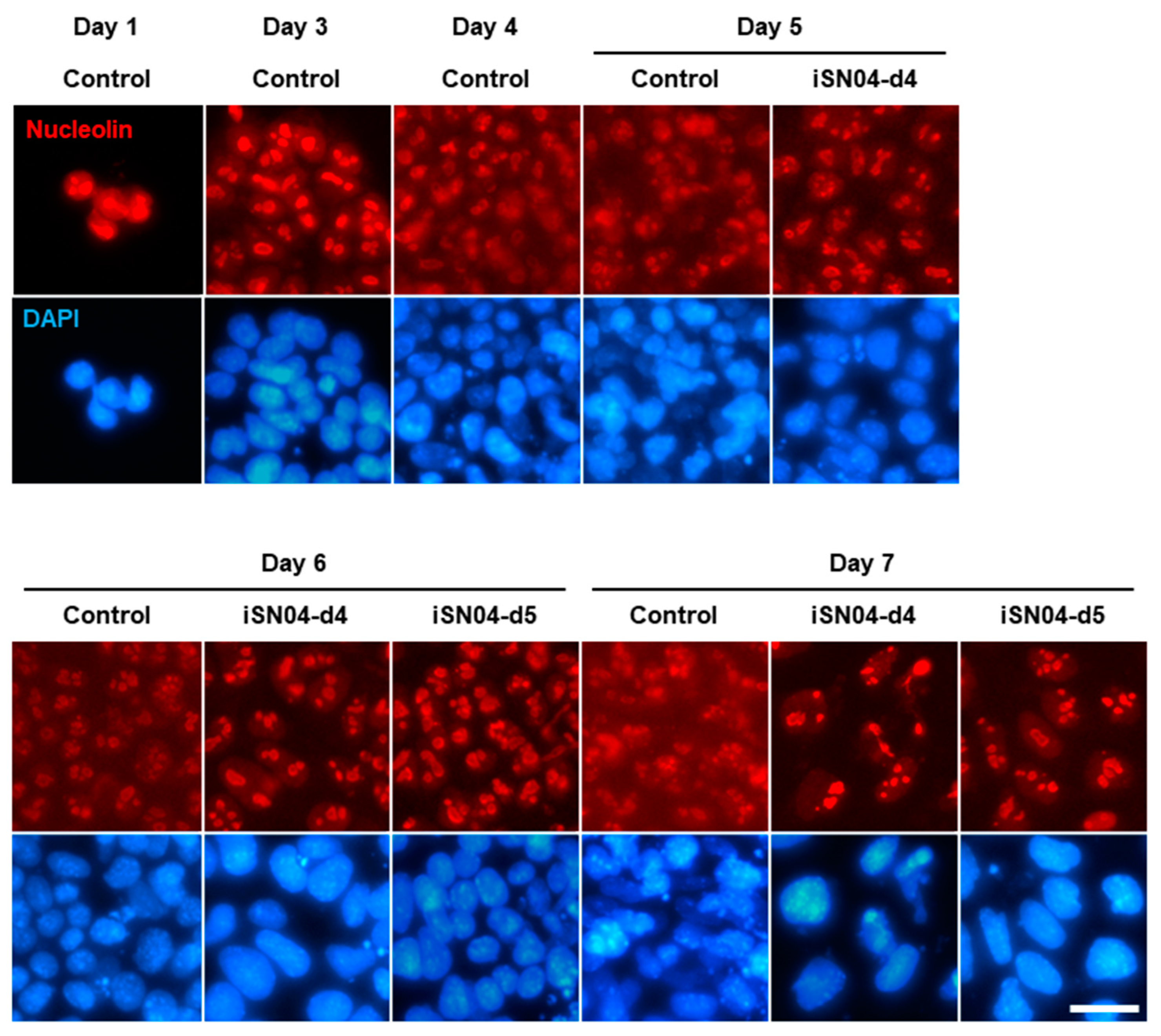
2.5. iSN04 Restricts Nucleolin Translocation in PSCs
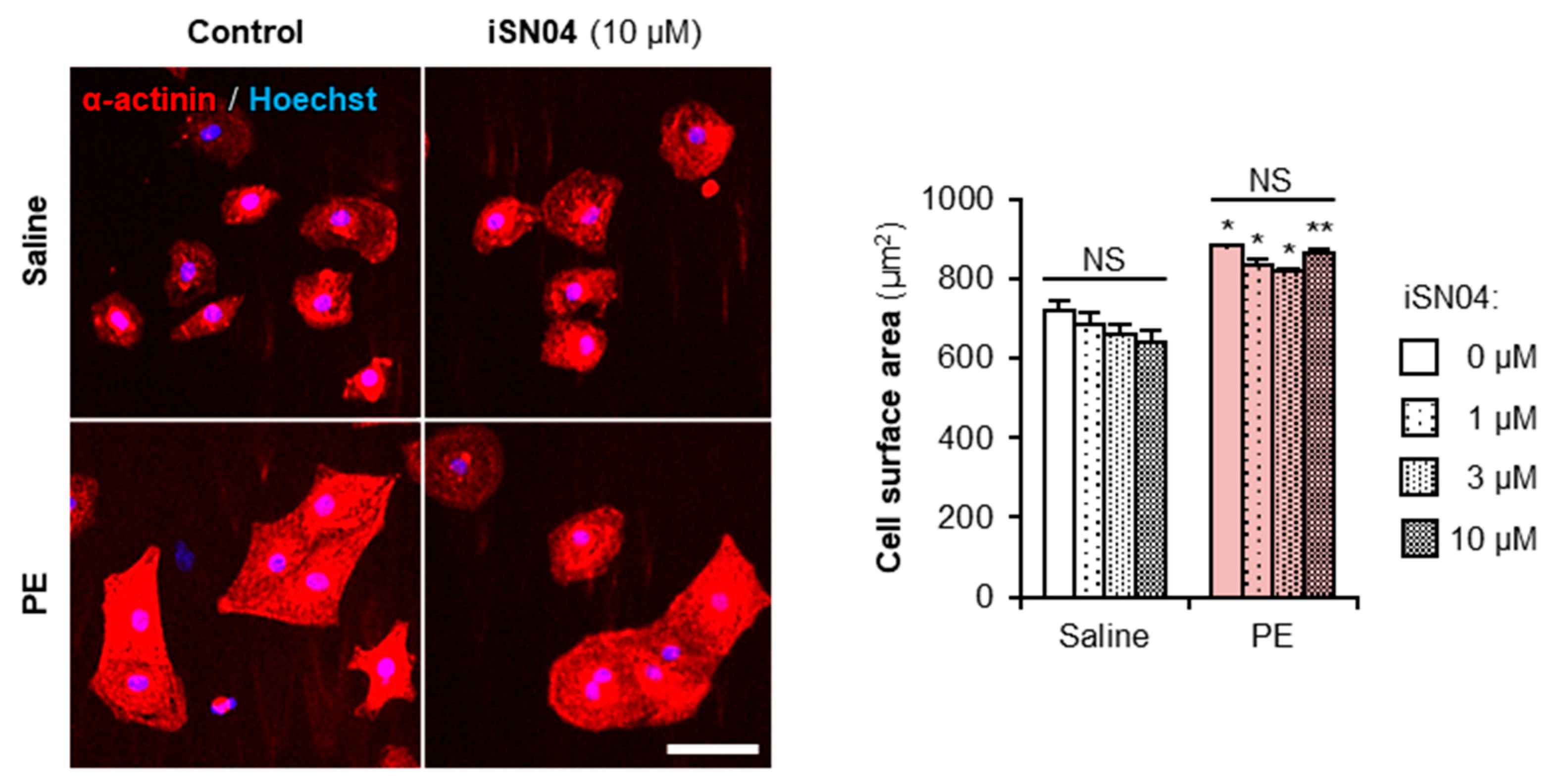
2.6. iSN04 Does Not Affect Myocardial Cell Hypertrophy
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. mESCs amd miPSCs
4.3. Cell Imaging
4.4. ALP Staining
4.5. Nucleolin Staining
4.6. qPCR
4.7. RNA-Seq
4.8. PCA
4.9. Heatmap and KEGG Pathway Analysis
4.10. Cardiomyocytes
4.11. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| DEG | Differentially expressed gene |
| DM | Differentiation medium |
| ESC | Embryonic stem cell |
| FPKM | Fragments per kilobase per million reads |
| GFP | Green fluorescent protein |
| GM | Growth medium |
| iPSC | Induced pluripotent stem cell |
| MEF | Murine embryonic fibroblast |
| MMC | Mitomycin C |
| myoDN | Myogenetic oligodeoxynucleotide |
| PCA | Principal component analysis |
| PE | Phenylephrine |
| PSC | Pluripotent stem cell |
| qPCR | Quantitative real-time RT-PCR |
| RNA-seq | RNA sequencing |
References
- Dogan, A. Embryonic stem cells in development and regenerative medicine. Adv. Exp. Med. Biol. 2018, 1079, 1–15. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, C.; Larcher, L.M.; Barrero, R.A.; Veedu, R.N. Three decades of nucleic acid aptamer technologies: Lessons learned, progress and opportunities on aptamer development. Biotechnol. Adv. 2019, 37, 28–50. [Google Scholar] [CrossRef] [PubMed]
- Ueki, R.; Atsuta, S.; Ueki, A.; Hoshiyama, J.; Li, J.; Hayashi, Y.; Sando, S. DNA aptamer assemblies as fibroblast growth factor mimics and their application in stem cell culture. Chem. Commun. 2019, 55, 2672–2675. [Google Scholar] [CrossRef]
- Shinji, S.; Umezawa, K.; Nihashi, Y.; Nakamura, S.; Shimosato, T.; Takaya, T. Identification of the myogenetic oligodeoxynucleotides (myoDNs) that promote differentiation of skeletal muscle myoblasts by targeting nucleolin. Front. Cell Dev. Biol. 2021, 8, 616706. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Yonekura, S.; Shimosato, T.; Takaya, T. Myogenetic oligodeoxynucleotide (myoDN) recovers the differentiation of skeletal muscle myoblasts deteriorated by diabetes mellitus. Front. Physiol. 2021, 12, 679152. [Google Scholar] [CrossRef] [PubMed]
- Nihashi, Y.; Shinji, S.; Umezawa, K.; Shimosato, T.; Ono, T.; Kagami, H.; Takaya, T. Myogenetic oligodeoxynucleotide complexed with berberine promotes differentiation of chicken myoblasts. Anim. Sci. J. 2021, 92, e13597. [Google Scholar] [CrossRef]
- Nihashi, Y.; Yamamoto, M.; Shimosato, T.; Takaya, T. Myogenetic oligodeoxynucleotide restores differentiation and reverses inflammation of myoblasts aggravated by cancer-conditioned medium. Muscles 2022, 1, 111–120. [Google Scholar] [CrossRef]
- Nohira, S.; Shinji, S.; Nakamura, Y.; Nihashi, Y.; Shimosato, T.; Takaya, T. Myogenetic oligodeoxynucleotides as anti-nucleolin aptamers inhibit the growth of embryonal rhabdomyosarcoma cells. Biomedicines 2022, 10, 2691. [Google Scholar] [CrossRef]
- Jia, W.; Yao, Z.; Zhao, J.; Guan, Q.; Gao, L. New perspectives of physiological and pathological functions of nucleolin (NCL). Life Sci. 2017, 186, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Laurincik, J.; Bjerregaard, B.; Strejcek, F.; Rath, D.; Niemann, H.; Rosenkranz, C.; Ochs, R.L.; Maddox-Hyttel, P. Nucleolar ultrastructure and protein allocation in in vitro produced porcine embryos. Mol. Reprod. Dev. 2004, 68, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, B.; Yang, A.; Lu, R.; Wang, W.; Zhou, Y.; Shi, G.; Kwon, S.W.; Zhao, Y.; Jin, Y. Ly-1 antibody reactive clone is an important nucleolar protein for control of self-renewal and differentiation in embryonic stem cells. Stem Cells 2009, 27, 1244–1254. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.; Svensson, F.; Runnberg, R.; Simonsson, T.; Simonsson, S. Phosphorylated nucleolin interacts with translationally controlled tumor protein during mitosis and with Oct4 during interphase in ES cells. PLoS ONE 2010, 5, e13678. [Google Scholar] [CrossRef]
- Yang, A.; Shi, G.; Zhou, C.; Lu, R.; Li, H.; Sun, L.; Jin, Y. Nucleolin maintains embryonic stem cell self-renewal by suppression of p53 protein-dependent pathway. J. Biol. Chem. 2011, 286, 43370–43382. [Google Scholar] [CrossRef]
- Percharde, M.; Lin, C.J.; Yin, Y.; Guan, J.; Peixoto, G.A.; Bulut-Karslioglu, A.; Biechele, S.; Huang, B.; Shen, X.; Ramalho-Santos, M. A LINE1-nucleolin partnership regulates early development and ESC identity. Cell 2018, 174, 391–405. [Google Scholar] [CrossRef]
- Okita, K.; Ichisaka, T.; Yamanaka, S. Generation of germline-competent induced pluripotent stem cells. Nature 2007, 488, 313–317. [Google Scholar] [CrossRef]
- Hidaka, K.; Lee, J.K.; Kim, H.S.; Ihm, C.H.; Iio, A.; Ogawa, M.; Nishikawa, S.; Kodama, I.; Morisaki, T. Chamber-specific differentiation of Nkx2.5-positive cardiac precursor cells from murine embryonic stem cells. FASEB J. 2003, 17, 740–742. [Google Scholar] [CrossRef]
- Kaichi, S.; Takaya, T.; Morimoto, T.; Sunagawa, Y.; Kawamura, T.; Ono, K.; Shimatsu, A.; Baba, S.; Heike, T.; Nakahata, T.; et al. Cyclin-dependent kinase 9 forms a complex with GATA4 and is involved in the differentiation of mouse ES cells into cardiomyocytes. J. Cell. Physiol. 2011, 226, 248–254. [Google Scholar] [CrossRef]
- Narita, S.; Unno, K.; Kato, K.; Okuno, Y.; Sato, Y.; Tsumura, Y.; Fujikawa, Y.; Shimizu, Y.; Hayashida, R.; Kondo, K.; et al. Direct reprogramming of adult adipose-derived regenerative cells toward cardiomyocytes using six transcriptional factors. iScience 2022, 25, 104651. [Google Scholar] [CrossRef]
- Mercola, M.; Ruiz-Lozano, P.; Schneider, M.D. Cardiac muscle regeneration: Lessons from development. Genes. Dev. 2011, 25, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 989–999. [Google Scholar]
- Mazzotta, S.; Neves, C.; Bonner, R.J.; Bernardo, A.S.; Docherty, K.; Hoppler, S. Distinctive roles of canonical and noncanonical Wnt signaling in human embryonic cardiomyocyte development. Stem Cell Rep. 2016, 7, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Schmeckpeper, J.; Verma, A.; Yin, L.; Beigi, F.; Zhang, L.; Payne, A.; Zhang, Z.; Pratt, R.E.; Dzau, V.J.; Mirotsou, M. Inhibition of Wnt6 by Sfrp2 regulates adult cardiac progenitor cell differentiation by differential modulation of Wnt pathways. J. Mol. Cell. Cardiol. 2015, 85, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Luedeke, D.M.; McCoy, L.; Iwafuchi, M.; Zorn, A.M. SOX transcription factors direct TCF-independent WNT/beta-catenin responsive transcription to govern cell fate in human pluripotent stem cells. Cell Rep. 2022, 40, 111247. [Google Scholar] [CrossRef] [PubMed]
- Naito, T.A.; Shiojima, I.; Akazawa, H.; Hidaka, K.; Morisaki, T.; Kikuchi, A.; Komuro, I. Developmental stage-specific biphasic roles of Wnt/β-catenin signaling in cardiomyogenesis and hematopoiesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19812–19817. [Google Scholar] [CrossRef]
- Frey, N.; Katus, H.A.; Olson, E.N.; Hill, J.A. Hypertrophy of the heart: A new therapeutic target? Circulation 2004, 109, 1580–1589. [Google Scholar] [CrossRef]
- Lei, H.; Hu, J.; Sun, K.; Xu, D. The role and molecular mechanism of epigenetics in cardiac hypertrophy. Heart Fail. Rev. 2020, 26, 1505–1514. [Google Scholar] [CrossRef]
- D’Amato, G.; Luxan, G.; de la Pmpa, J.L. Notch signalling in ventricular chamber development and cardiomyopathy. FEBS J. 2016, 283, 4223–4237. [Google Scholar] [CrossRef]
- Tajbakhsh, S. Skeletal muscle stem cells in developmental versus regenerative myogenesis. J. Intern. Med. 2009, 266, 372–389. [Google Scholar] [CrossRef]
- Mizuno, Y.; Chang, H.; Umeda, K.; Niwa, A.; Iwasa, T.; Awaya, T.; Fukada, S.; Yamamoto, H.; Yamanaka, S.; Nakahata, T.; et al. Generation of skeletal muscle stem/progenitor cells from murine induced pluripotent stem cells. FASEB J. 2010, 24, 2245–2253. [Google Scholar] [CrossRef]
- Mummery, C.L.; Zhang, J.; Ng, E.S.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: A methods overview. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Reister, S.; Mahotka, C.; van den Hofel, N.; Grinstein, E. Nucleolin promotes Wnt signaling in human hematopoietic stem/progenitor cells. Leukemia 2019, 33, 1052–1054. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Miyoshi, M.; Morioka, K.; Mitani, T.; Takaya, T. Anti-nucleolin aptamer, iSN04, inhibits the inflammatory responses in C2C12 myoblasts by modulating the β-catenin/NF-κB signaling pathway. Biochem. Biophys. Res. Commun. 2023, 664, 1–8. [Google Scholar] [CrossRef]
- Liu, J.J.; Shentu, L.M.; Ma, N.; Wang, L.Y.; Zhang, G.M.; Sun, Y.; Wang, Y.; Li, J.; Mu, Y.L. Inhibition of NF-κB and Wnt/β-catenin/GSK3β signaling pathways ameliorates cardiomyocyte hypertrophy and fibrosis in streptozotocin (STZ)-induced type 1 diabetic rats. Curr. Med. Sci. 2020, 40, 35–47. [Google Scholar] [CrossRef]
- Nihashi, Y.; Miyoshi, M.; Umezawa, K.; Shimosato, T.; Takaya, T. Identification of a novel osteogenetic oligodeoxynucleotide (osteoDN) that promotes osteoblast differentiation in a TLR9-independent manner. Nanomaterials 2022, 12, 1680. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acid. Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Takaya, T.; Kawamura, T.; Morimoto, T.; Ono, K.; Kita, T.; Shimatsu, A.; Hasegawa, K. Identification of p300-targeted acetylated residues in GATA4 during hypertrophic responses in cardiac myocytes. J. Biol. Chem. 2008, 283, 9828–9835. [Google Scholar] [CrossRef]
- Liu, W.; Brown, K.; Legros, S.; Foley, A.C. Nodal mutant eXtraembryonic ENdoderm (XEN) stem cells upregulate markers for the anterior visceral endoderm and impact the timing of cardiac differentiation in mouse embryoid bodies. Biol. Open 2012, 1, 208–219. [Google Scholar] [CrossRef]
- Sakurai, H.; Okawa, Y.; Inami, Y.; Nishio, N.; Isobe, K. Paraxial mesodermal progenitors derived from mouse embryonic stem cells contribute to muscle regeneration via differentiation into muscle satellite cells. Stem Cells 2008, 26, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; McDonald, C.; Petrenko, N.B.; Leblanc, M.; Wang, T.; Giguere, V.; Evans, R.M.; Patel, V.V.; Pei, L. Estrogen-related receptor α (ERRα) and ERRγ are essential coordinators of cardiac metabolism and function. Mol. Cell. Biol. 2015, 35, 1281–1298. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Mapped Reads | % Mapped |
|---|---|---|---|---|
| Control, day 4 | 50,439,826 | 497,66,540 | 46,032,264 | 92.50% |
| Control, day 5 | 54,721,348 | 54,064,028 | 50,082,857 | 92.64% |
| Control, day 6 | 54,782,002 | 54,174,286 | 50,116,711 | 92.51% |
| Control, day 7 | 48,692,658 | 48,064,434 | 44,481,973 | 92.55% |
| iSN04-d4, day 5 | 59,925,566 | 59,225,592 | 54,811,573 | 92.55% |
| iSN04-d4, day 6 | 52,889,966 | 52,342,122 | 47,145,750 | 90.07% |
| iSN04-d4, day 7 | 60,513,932 | 59,768,940 | 55,336,829 | 92.58% |
| iSN04-d5, day 6 | 56,021,784 | 55,366,368 | 51,361,265 | 92.77% |
| iSN04-d5, day 7 | 55,013,818 | 54,174,286 | 50,213,955 | 92.51% |
| Subset | Gene |
|---|---|
| Day 6, High-ctrl | Cer1, Daam2, Dkk1, Fzd10, Sfrp5, Sox17, Wnt3, Wnt5a, Wnt6, Wnt8a |
| Day 6, High-d5 | Fzd4, Mapk10, Prkcb, Wnt1 |
| Day 7, High-ctrl | Cer1, Dkk1, Fzd10, Sox17, Wnt3, Wnt6 |
| Day 7, High-d5 | Sfrp5, Wnt1, Wnt2, Wnt5a, Wnt11, Wnt16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishioka, M.; Nihashi, Y.; Sunagawa, Y.; Umezawa, K.; Shimosato, T.; Kagami, H.; Morimoto, T.; Takaya, T. Myogenetic Oligodeoxynucleotide Induces Myocardial Differentiation of Murine Pluripotent Stem Cells. Int. J. Mol. Sci. 2023, 24, 14380. https://doi.org/10.3390/ijms241814380
Ishioka M, Nihashi Y, Sunagawa Y, Umezawa K, Shimosato T, Kagami H, Morimoto T, Takaya T. Myogenetic Oligodeoxynucleotide Induces Myocardial Differentiation of Murine Pluripotent Stem Cells. International Journal of Molecular Sciences. 2023; 24(18):14380. https://doi.org/10.3390/ijms241814380
Chicago/Turabian StyleIshioka, Mina, Yuma Nihashi, Yoichi Sunagawa, Koji Umezawa, Takeshi Shimosato, Hiroshi Kagami, Tatsuya Morimoto, and Tomohide Takaya. 2023. "Myogenetic Oligodeoxynucleotide Induces Myocardial Differentiation of Murine Pluripotent Stem Cells" International Journal of Molecular Sciences 24, no. 18: 14380. https://doi.org/10.3390/ijms241814380