
Transcription Factor Nrf2 Modulates Lipopolysaccharide-Induced Injury in Bovine Endometrial Epithelial Cells
 ,
,
Abstract
:1. Introduction
2. Results
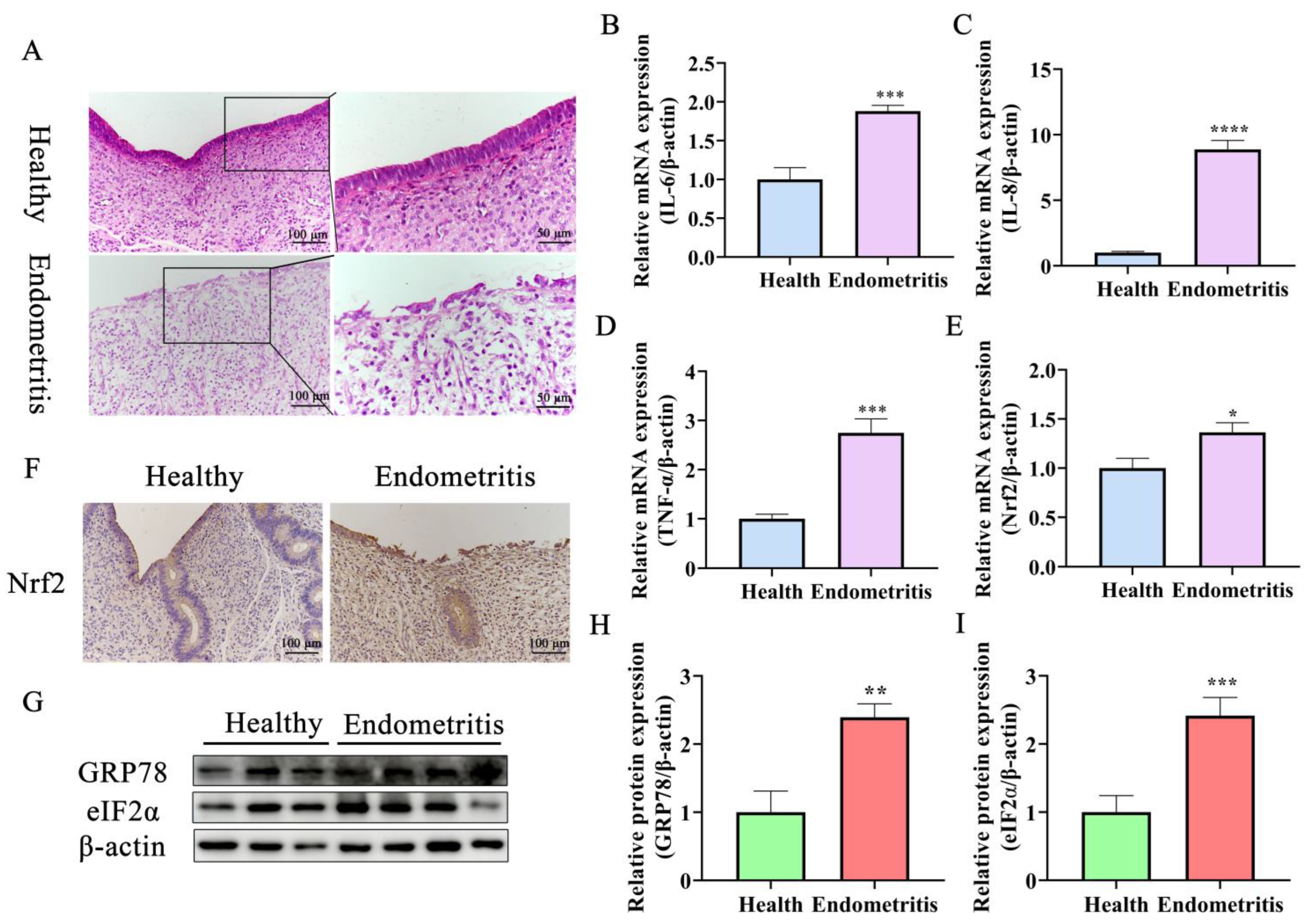
2.1. Nrf2 Expression Is Increased Significantly in Endometriotic Uteri and Is Associated with ERS and UPR
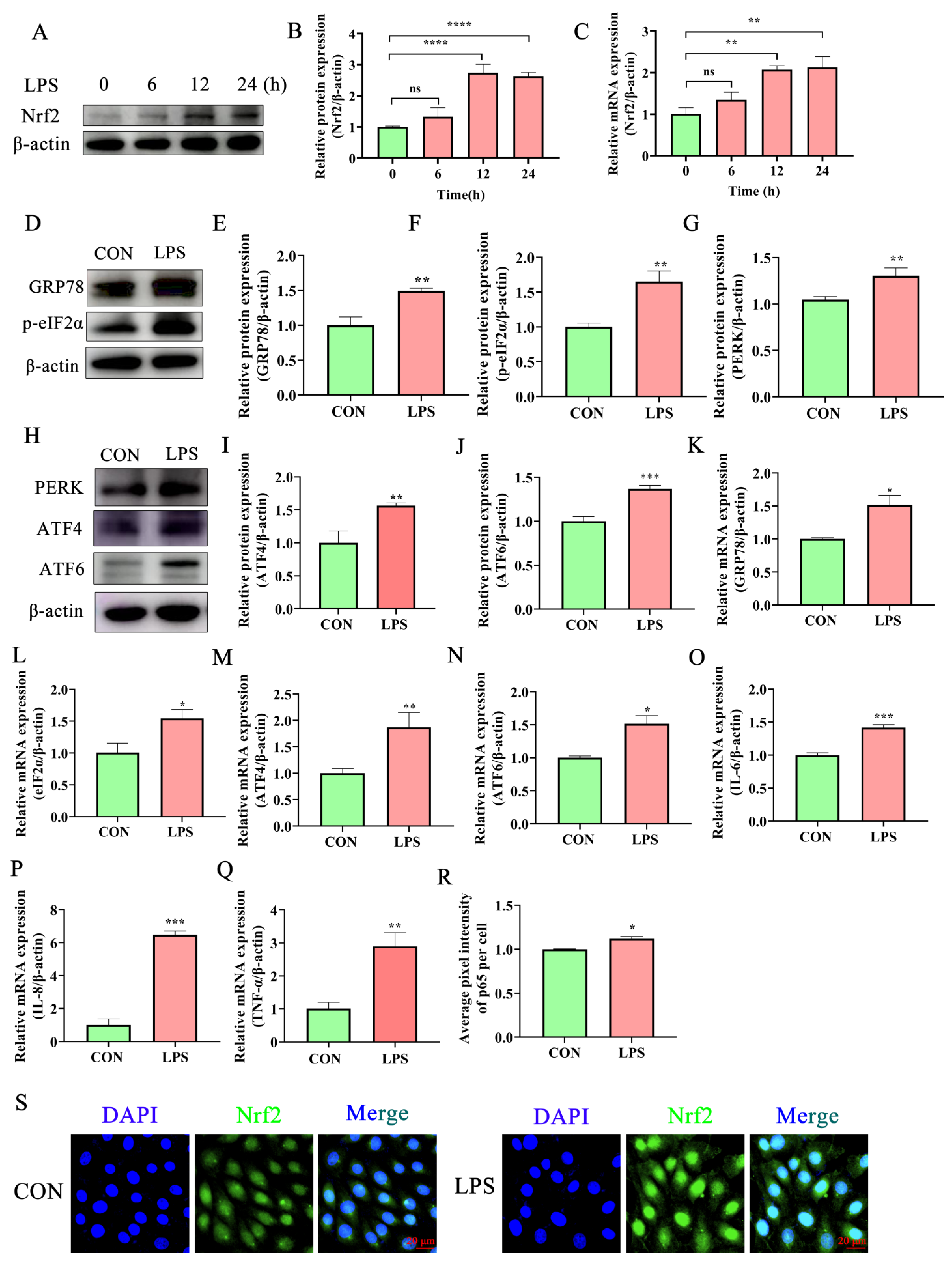
2.2. Lipopolysaccharide Activates the Inflammatory Response and ERS in Bovine Endometrial Epithelial Cells
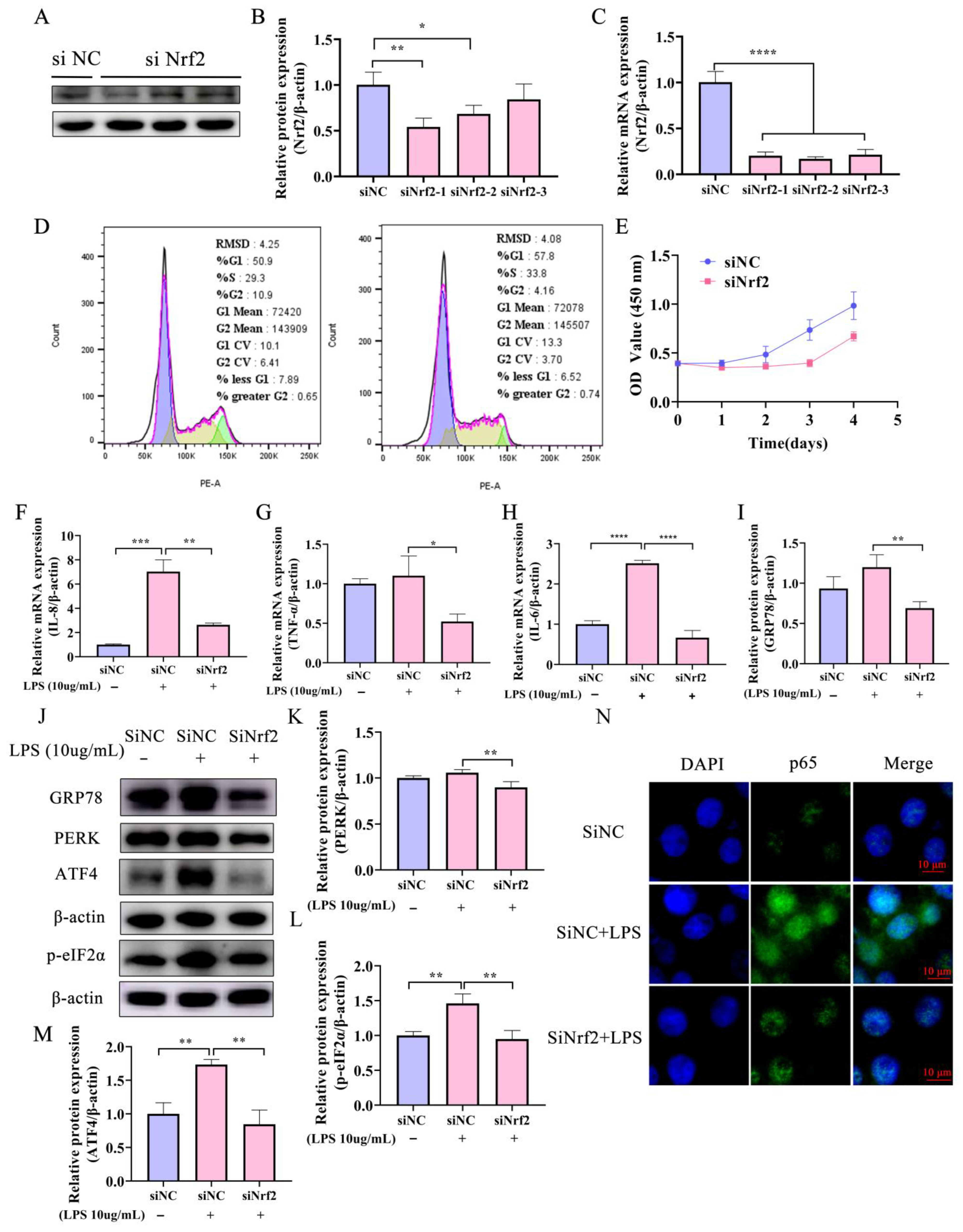
2.3. Knockdown of Nrf2 Reduces Both the LPS-Induced Inflammatory Response and ERS in Bovine Endometrial Epithelial Cells
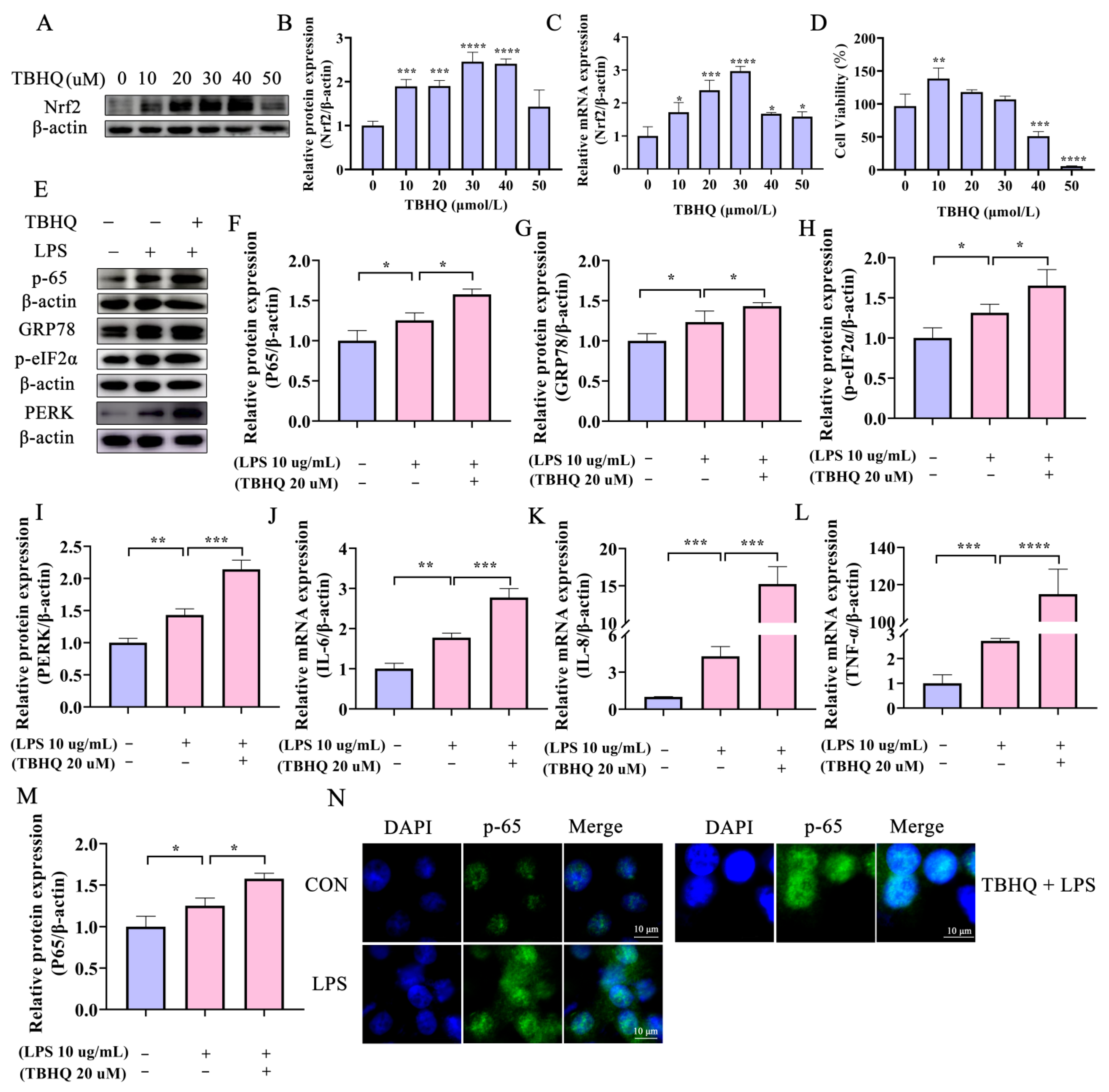
2.4. Activation of Nrf2 Aggravates the LPS-Induced Inflammatory Response in Bovine Endometrial Epithelial Cells through the ERS PERK Pathway
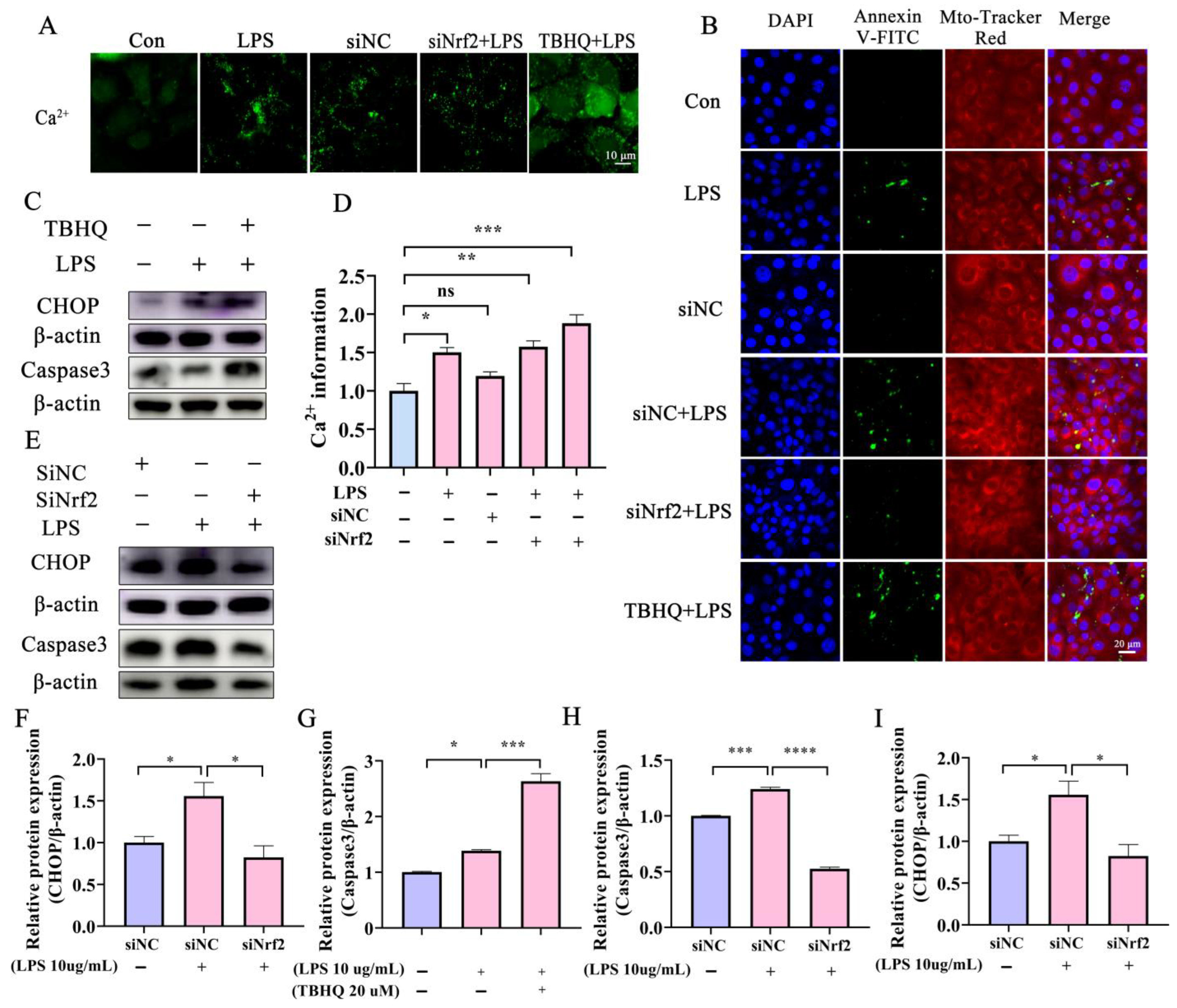
2.5. Disorder of Ca2+ and CHOP Promote ERS-Mediated Damage in Bovine Endometrial Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Bovine Uterine Collection and Cell Culture
4.2. Total RNA Extraction and Real-Time Quantitative PCR
4.3. Western Blot Analysis
4.4. Hematoxylin–Eosin Stain
4.5. Immunohistochemistry
4.6. Immunofluorescence Staining
4.7. RNA Interference
4.8. Flow Cytometry
4.9. Cell Toxicity Assay
4.10. Intracellular Ca2+ Detection
4.11. Apoptosis Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, M.M.; Causey, R.C. Clinical and subclinical endometritis in the mare: Both threats to fertility. Reprod. Domest. Anim. Zuchthyg. 2009, 44 (Suppl. S3), 10–22. [Google Scholar] [CrossRef] [PubMed]
- Lincke, A.; Drillich, M.; Heuwieser, W. Subclinical endometritis in dairy cattle and its effect on fertility—A review of recent publications. Berl. Munch. Tierarztl. Wochenschr. 2007, 120, 245–250. [Google Scholar]
- Oladejo, A.O.; Li, Y.; Wu, X.; Imam, B.H.; Shen, W.; Ding, X.Z.; Wang, S.; Yan, Z. MicroRNAome: Potential and Veritable Immunomolecular Therapeutic and Diagnostic Baseline for Lingering Bovine Endometritis. Front. Vet. Sci. 2020, 7, 614054. [Google Scholar] [CrossRef]
- Saito, A.; Imaizumi, K. Unfolded Protein Response-Dependent Communication and Contact among Endoplasmic Reticulum, Mitochondria, and Plasma Membrane. Int. J. Mol. Sci. 2018, 19, 3215. [Google Scholar] [CrossRef] [Green Version]
- Senft, D.; Ronai, Z.A. UPR, autophagy, and mitochondria crosstalk underlies the ER stress response. Trends Biochem. Sci. 2015, 40, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, L.; Weng, X.; Chen, H.; Du, Y.; Diao, C.; Chen, Z.; Liu, X. Inhibition of Brd4 alleviates renal ischemia/reperfusion injury-induced apoptosis and endoplasmic reticulum stress by blocking FoxO4-mediated oxidative stress. Redox Biol. 2019, 24, 101195. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Zhang, X.; Ye, Y.; Xiong, X.; Zhang, S.; Gu, L.; Jian, Z.; Wang, H. Endoplasmic Reticulum Stress and the Unfolded Protein Response in Cerebral Ischemia/Reperfusion Injury. Front. Cell. Neurosci. 2022, 16, 864426. [Google Scholar] [CrossRef]
- Li, W.; Cao, T.; Luo, C.; Cai, J.; Zhou, X.; Xiao, X.; Liu, S. Crosstalk between ER stress, NLRP3 inflammasome, and inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 6129–6140. [Google Scholar] [CrossRef]
- Liu, J.; Qiu, R.; Liu, R.; Song, P.; Lin, P.; Chen, H.; Zhou, D.; Wang, A.; Jin, Y. Autophagy Mediates Escherichia Coli-Induced Cellular Inflammatory Injury by Regulating Calcium Mobilization, Mitochondrial Dysfunction, and Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2022, 23, 14174. [Google Scholar] [CrossRef]
- Kumari, N.; Reabroi, S.; North, B.J. Unraveling the Molecular Nexus between GPCRs, ERS, and EMT. Mediat. Inflamm. 2021, 2021, 6655417. [Google Scholar] [CrossRef]
- Ren, J.; Bi, Y.; Sowers, J.R.; Hetz, C.; Zhang, Y. Endoplasmic reticulum stress and unfolded protein response in cardiovascular diseases. Nat. Rev. Cardiol. 2021, 18, 499–521. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, J.; Yang, N.; Huang, Y.; Hu, T.; Rao, C. Endoplasmic reticulum stress-mediated cell death in liver injury. Cell Death Dis. 2022, 13, 1051. [Google Scholar] [CrossRef]
- Zhou, R.; Ma, Y.; Tao, Z.; Qiu, S.; Gong, Z.; Tao, L.; Zhu, Y. Melatonin Inhibits Glucose-Induced Apoptosis in Osteoblastic Cell Line Through PERK-eIF2α-ATF4 Pathway. Front. Pharmacol. 2020, 11, 602307. [Google Scholar] [CrossRef]
- Yi, S.; Chen, K.; Zhang, L.; Shi, W.; Zhang, Y.; Niu, S.; Jia, M.; Cong, B.; Li, Y. Endoplasmic Reticulum Stress Is Involved in Stress-Induced Hypothalamic Neuronal Injury in Rats via the PERK-ATF4-CHOP and IRE1-ASK1-JNK Pathways. Front. Cell. Neurosci. 2019, 13, 190. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta. Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Xing, X.; Zhang, Y.; Zhang, C.; Wu, Y.; Chen, Y.; Meng, R.; Jia, H.; Cheng, Y.; Zhang, Y.; et al. Lead exposure activates the Nrf2/Keap1 pathway, aggravates oxidative stress, and induces reproductive damage in female mice. Ecotoxicol. Environ. Saf. 2021, 207, 111231. [Google Scholar] [CrossRef]
- Sun, T.C.; Liu, X.C.; Yang, S.H.; Song, L.L.; Zhou, S.J.; Deng, S.L.; Tian, L.; Cheng, L.Y. Melatonin Inhibits Oxidative Stress and Apoptosis in Cryopreserved Ovarian Tissues via Nrf2/HO-1 Signaling Pathway. Front. Mol. Biosci. 2020, 7, 163. [Google Scholar] [CrossRef]
- Hybertson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative stress in health and disease: The therapeutic potential of Nrf2 activation. Mol. Asp. Med. 2011, 32, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, D.; Yang, Z.; Xue, P.; Liu, X. Nrf2/HO-1 pathway is involved the anti-inflammatory action of intrauterine infusion of platelet-rich plasma against lipopolysaccharides in endometritis. Immunopharmacol. Immunotoxicol. 2022, 44, 119–128. [Google Scholar] [CrossRef]
- Fu, K.; Chen, H.; Wang, Z.; Cao, R. Andrographolide attenuates inflammatory response induced by LPS via activating Nrf2 signaling pathway in bovine endometrial epithelial cells. Res. Vet. Sci. 2021, 134, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, D.; Yan, Z. Increasing of matrix metalloproteinase 3 in bovine endometritis. Theriogenology 2021, 175, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, D.; Wu, X.; He, B.; Cheng, Z.; Szenci, O.; Song, P.; Shao, D.; Zhang, S.; Yan, Z. Decreasing of S100A4 in bovine endometritis in vivo and in vitro. Theriogenology 2020, 153, 68–73. [Google Scholar] [CrossRef]
- Adeniran, S.O.; Zheng, P.; Feng, R.; Adegoke, E.O.; Huang, F.; Ma, M.; Wang, Z.; Ifarajimi, O.O.; Li, X.; Zhang, G. The Antioxidant Role of Selenium via GPx1 and GPx4 in LPS-Induced Oxidative Stress in Bovine Endometrial Cells. Biol. Trace Elem. Res. 2022, 200, 1140–1155. [Google Scholar] [CrossRef]
- Deng, S.; Wu, D.; Li, L.; Li, J.; Xu, Y. TBHQ attenuates ferroptosis against 5-fluorouracil-induced intestinal epithelial cell injury and intestinal mucositis via activation of Nrf2. Cell. Mol. Biol. Lett. 2021, 26, 48. [Google Scholar] [CrossRef]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. CMLS 2016, 73, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Gao, S.; Wang, J.; Yu, F.; Ye, C. Protective effects of chicoric acid on LPS-induced endometritis in mice via inhibiting ferroptosis by Nrf2/HO-1 signal axis. Int. Immunopharmacol. 2022, 113, 109435. [Google Scholar] [CrossRef]
- Cheng, C.; Ma, H.; Liu, G.; Fan, S.; Deng, Y.; Jiang, J.; Feng, J.; Guo, Z. The role of Nrf2 signaling pathway in the mud crab (Scylla paramamosain) in response to Vibrio parahaemolyticus infection. Fish Shellfish Immunol. 2023, 136, 108729. [Google Scholar] [CrossRef]
- Suzuki, T.; Takahashi, J.; Yamamoto, M. Molecular Basis of the KEAP1-NRF2 Signaling Pathway. Mol. Cells 2023, 46, 133–141. [Google Scholar] [CrossRef]
- Wang, L.; He, C. Nrf2-mediated anti-inflammatory polarization of macrophages as therapeutic targets for osteoarthritis. Front. Immunol. 2022, 13, 967193. [Google Scholar] [CrossRef]
- Yasuo, T.; Kitaya, K. Challenges in Clinical Diagnosis and Management of Chronic Endometritis. Diagnostics 2022, 12, 2711. [Google Scholar] [CrossRef]
- Kitaya, K.; Takeuchi, T.; Mizuta, S.; Matsubayashi, H.; Ishikawa, T. Endometritis: New time, new concepts. Fertil. Steril. 2018, 110, 344–350. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, S.; Liu, J.; Ta, N. Inhibition of TLR2/TLR4 alleviates the Neisseria gonorrhoeae infection damage in human endometrial epithelial cells via Nrf2 and NF-Kβsignaling. J. Reprod. Immunol. 2020, 142, 103192. [Google Scholar] [CrossRef]
- Jiang, P.Y.; Zhu, X.J.; Zhang, Y.N.; Zhou, F.F.; Yang, X.F. Protective effects of apigenin on LPS-induced endometritis via activating Nrf2 signaling pathway. Microb. Pathog. 2018, 123, 139–143. [Google Scholar] [CrossRef]
- Fu, K.; Wang, Z.; Cao, R. Berberine attenuates the inflammatory response by activating the Keap1/Nrf2 signaling pathway in bovine endometrial epithelial cells. Int. Immunopharmacol. 2021, 96, 107738. [Google Scholar] [CrossRef]
- Gao, F.; Li, H.; Feng, Y.; Tian, W.; Cao, R.; Fu, K. Aucubin ameliorates the LPS-induced inflammatory response in bovine endometrial epithelial cells by inhibiting NF-κB and activating the Keap1/Nrf2 signalling pathway. Reprod. Domest. Anim. Zuchthyg. 2021, 56, 972–982. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Fusco, R.; Licata, P.; Peritore, A.F.; D’Amico, R.; Cordaro, M.; Siracusa, R.; Cuzzocrea, S.; Crupi, R. Protective Effect of Hydroxytyrosol on LPS-Induced Inflammation and Oxidative Stress in Bovine Endometrial Epithelial Cell Line. Vet. Sci. 2020, 7, 161. [Google Scholar] [CrossRef]
- Cui, L.; Guo, J.; Wang, Z.; Zhang, J.; Li, W.; Dong, J.; Liu, K.; Guo, L.; Li, J.; Wang, H.; et al. Meloxicam inhibited oxidative stress and inflammatory response of LPS-stimulated bovine endometrial epithelial cells through Nrf2 and NF-κB pathways. Int. Immunopharmacol. 2023, 116, 109822. [Google Scholar] [CrossRef]
- Marciniak, S.J.; Chambers, J.E.; Ron, D. Pharmacological targeting of endoplasmic reticulum stress in disease. Nat. Rev. Drug Discov. 2022, 21, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Coleman, O.I.; Haller, D. ER Stress and the UPR in Shaping Intestinal Tissue Homeostasis and Immunity. Front. Immunol. 2019, 10, 2825. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, R.L.; Mesgarzadeh, J.S.; Hendershot, L.M. Reshaping endoplasmic reticulum quality control through the unfolded protein response. Mol. Cell 2022, 82, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Dixon, B.J.; Doycheva, D.M.; Li, B.; Zhang, Y.; Hu, Q.; He, Y.; Guo, Z.; Nowrangi, D.; Flores, J.; et al. IRE1α inhibition decreased TXNIP/NLRP3 inflammasome activation through miR-17-5p after neonatal hypoxic-ischemic brain injury in rats. J. Neuroinflamm. 2018, 15, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Lu, W.; Doycheva, D.M.; Gamdzyk, M.; Hu, X.; Liu, R.; Zhang, J.H.; Tang, J. IRE1α inhibition attenuates neuronal pyroptosis via miR-125/NLRP1 pathway in a neonatal hypoxic-ischemic encephalopathy rat model. J. Neuroinflamm. 2020, 17, 152. [Google Scholar] [CrossRef]
- Hu, X.; Li, D.; Wang, J.; Guo, J.; Li, Y.; Cao, Y.; Zhang, N.; Fu, Y. Melatonin inhibits endoplasmic reticulum stress-associated TXNIP/NLRP3 inflammasome activation in lipopolysaccharide-induced endometritis in mice. Int. Immunopharmacol. 2018, 64, 101–109. [Google Scholar] [CrossRef]
- Sakai, S.; Inoue, Y.; Tanaka, K.; Yamamoto, Y.; Iwata, H.; Kimura, K. Hyperthermia alters interleukin-6 production in response to lipopolysaccharide via endoplasmic reticulum stress in bovine endometrial cells. J. Cell. Physiol. 2022, 237, 1087–1099. [Google Scholar] [CrossRef]
- Hu, H.; Tian, M.; Ding, C.; Yu, S. The C/EBP Homologous Protein (CHOP) Transcription Factor Functions in Endoplasmic Reticulum Stress-Induced Apoptosis and Microbial Infection. Front. Immunol. 2018, 9, 3083. [Google Scholar] [CrossRef] [Green Version]
- Asmat, T.M.; Tenenbaum, T.; Jonsson, A.B.; Schwerk, C.; Schroten, H. Impact of calcium signaling during infection of Neisseria meningitidis to human brain microvascular endothelial cells. PLoS ONE 2014, 9, e114474. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zeng, Q.; Ling, M.; Ma, R.; Chen, H.; Lin, F.; Li, Z.; Pan, L. Inhibition of IP3R/Ca2+ Dysregulation Protects Mice From Ventilator-Induced Lung Injury via Endoplasmic Reticulum and Mitochondrial Pathways. Front. Immunol. 2021, 12, 729094. [Google Scholar] [CrossRef]
- Szymański, J.; Janikiewicz, J.; Michalska, B.; Patalas-Krawczyk, P.; Perrone, M.; Ziółkowski, W.; Duszyński, J.; Pinton, P.; Dobrzyń, A.; Więckowski, M.R. Interaction of Mitochondria with the Endoplasmic Reticulum and Plasma Membrane in Calcium Homeostasis, Lipid Trafficking and Mitochondrial Structure. Int. J. Mol. Sci. 2017, 18, 1576. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Zhou, S.; Xie, W.; Meng, M.; Ma, N.; Zhang, H.; Wang, Y.; Chang, G.; Shen, X. STIM1-Orai1 Interaction Exacerbates LPS-Induced Inflammation and Endoplasmic Reticulum Stress in Bovine Hepatocytes through Store-Operated Calcium Entry. Genes 2022, 13, 874. [Google Scholar] [CrossRef]
- Zeeshan, H.M.; Lee, G.H.; Kim, H.R.; Chae, H.J. Endoplasmic Reticulum Stress and Associated ROS. Int. J. Mol. Sci. 2016, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Liu, C.; Sun, M.; Liu, J.; Lin, P.; Wang, A.; Jin, Y. Oxidativae Stress Induces Bovine Endometrial Epithelial Cell Damage through Mitochondria-Dependent Pathways. Animals 2022, 12, 2444. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ID | Sequences (5′-3′) | Product Size (bp) |
|---|---|---|---|
| β-actin | NM_173979.3 | F: CCGCAACCAGTTCGCCAT R: CCCACGTACGAGTCCTTCTG | 180 |
| TNF-α | NM_173966.3 | F: CTCCTTCCTCCTGGTTGCAG R: CACCTGGGGACTGCTCTTC | 92 |
| IL-6 | NM_173923.2 | F: CAGATCCTGAAGCAAAAGATCGC R: CCCACTCGTTTGAAGACTGC | 91 |
| IL-8 | NM_173925.2 | F: CATTCCACACCTTTCCACCC R: AGGCAGACCTCGTTTCCATT | 116 |
| Nrf2 | XM_005202312.4 | F: ATTCAAGTGCCACAGTAA R: AAAGTAGCAGAGGAGGG | 100 |
| GRP78 | NM_001075148.1 | F: GTGCCCACCAAGAAGTCTCA R: GTCAGGGGTCGTTCACCTTC | 92 |
| PERK | NM_001098086.1 | F: CACAGGGACCTCAAGCCTTC R: TCCTCGTCTTGGTCCATTGC | 98 |
| ATF4 | NM_001034342.2 | F: GCTTAAGCCATGGCGCTTTT R: ATGTTGCGAGGTTTTGGTGC | 111 |
| ATF6 | XM_024989877.1 | F: TACTTCCAGCAGCACCCAAG R: GCACCACGGTCTGACCTTTA | 150 |
| eIF2α | NM_175813.2 | F: ACCACCCTGGAGAGAACAGA R: GTGACCACTTTGGGCTCCAT | 119 |
| IRE1 | XM_024980955.1 | F: GCCATGAGGAATAAGAAGCACC R: TGGCATGGTAGGTGTGTGAG | 136 |
| Gene | Sequence (5′ to 3′) |
|---|---|
| si-NC | F: UUCUCCGAACGUGUCACGUTT R: ACGUGACACGUUCGGAGAATT |
| Si-Nrf2-1 | F: GCCCAUUGAUCUCUCUGAUTT R: AUCAGAGAGAUCAAUGGGCTT |
| Si-Nrf2-2 | F: GGAGCAAGAUUUAGAUCAUTT R: AUGAUCUAAAUCUUGCUCCTT |
| Si-Nrf2-3 | F: GAGGCCAGAUAUUAAGAAATT R: UUUCUUAAUAUCUGGCCUCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, P.; Liu, C.; Sun, M.; Liu, J.; Lin, P.; Chen, H.; Zhou, D.; Tang, K.; Wang, A.; Jin, Y. Transcription Factor Nrf2 Modulates Lipopolysaccharide-Induced Injury in Bovine Endometrial Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 11221. https://doi.org/10.3390/ijms241311221
Song P, Liu C, Sun M, Liu J, Lin P, Chen H, Zhou D, Tang K, Wang A, Jin Y. Transcription Factor Nrf2 Modulates Lipopolysaccharide-Induced Injury in Bovine Endometrial Epithelial Cells. International Journal of Molecular Sciences. 2023; 24(13):11221. https://doi.org/10.3390/ijms241311221
Chicago/Turabian StyleSong, Pengjie, Chen Liu, Mingkun Sun, Jianguo Liu, Pengfei Lin, Huatao Chen, Dong Zhou, Keqiong Tang, Aihua Wang, and Yaping Jin. 2023. "Transcription Factor Nrf2 Modulates Lipopolysaccharide-Induced Injury in Bovine Endometrial Epithelial Cells" International Journal of Molecular Sciences 24, no. 13: 11221. https://doi.org/10.3390/ijms241311221