1. Introduction
Chickens are economically important model animals. An in-depth understanding of the mechanism of sex determination and gonadal development in chickens has both theoretical and practical applications. In poultry production, it will be of great value to the poultry industry if the sex determination mechanism can be used to control the gender of hatched chicks which is naturally half male and half female. Differences in sex chromosomes between males and females form the genetic basis for sex determination. Similar to other birds, chickens have a Z/W sex chromosome system in which males are homozygous (ZZ) and females heterozygous (ZW). The Z chromosome is a large chromosome with >1000 genes, while the W chromosome is its degenerate copy containing only 28 protein-coding genes [
1,
2]. Due to the lack of global dosage compensation, such as the X chromosome in mammals, most Z chromosome genes in chickens have a dosage difference between males and females [
3]. According to the most recent build of the chicken W chromosome (GRCg7b), all the protein-coding W genes have their homologous copies on the Z chromosome. These Z-W homologues are thought to have important roles in development [
2]. Although some of the W genes were reported to be related to early sex differentiation [
4], the Z chromosome dosage effect is a widely accepted mechanism for sex determination in chickens [
5,
6]. A recent study has supported this view. The heterozygous knockout of the Z chromosome gene
DMRT1 reportedly allows the complete development of ovaries in ZZ-type chicken embryos, indicating that
DMRT1 is a good candidate gene for chicken sex determination [
7,
8]. However, these Z
+Z
− individuals did not develop into functional females (unable to lay eggs) when they reached adulthood and remained male in appearance and body type [
7]. This suggests that chickens may have more complex mechanisms of gonadal development (especially for females) other than the sole control of Z chromosomal genes’ dosage effect, such as the synergistic effects of W chromosomal genes. Therefore, in our previous study, we examined the expression profiles of the 28 W chromosome protein-coding genes and their Z chromosome homologs in chicken embryonic gonads to screen for genes that show differential expression between females and males and identify genes that may be involved in gonadal development in chickens.
Here we studied one of the W chromosome genes
MIER3 (mesoderm-induced early response protein family member 3), which is a transcription factor [
9]. In our preliminary results,
MIER3 showed spatiotemporal expression specificity (W chromosome copy is highly expressed in the gonads, and there are differences in overall gene expression between males and females). So, we suggested that
MIER3 might be involved in the development of chicken gonads. To test this hypothesis, we investigated the detailed expression pattern of
MIER3 in chicken embryonic tissues (mainly the gonads). Additionally, we explored its potential function in embryonic gonadal development using a fadrozole (FAD, an aromatase inhibitor which can cause female-to-male sex reversal) induced sex reversal model and analyzed the downstream pathways by
MIER3 overexpression in male gonadal cells.
3. Discussion
Sex determination and gonadal development is a delicate and complex process controlled by multiple genes. Birds have Z and W sex chromosomes, with males being homozygous (ZZ) and females being heterozygous (ZW), different from the XY chromosome system in mammals, such as humans. Correspondingly, sex determination mechanisms in birds are different from those in mammals. To date, no sex-determining genes similar to
SRY (sex-determining region Y) on the human Y chromosome have been identified in chickens [
10]. Although the
DMRT1 gene on the chicken Z chromosome is considered the most likely sex-determining gene [
11], female gonadal development seems to have a more complicated mechanism, which involves multiple pathways (e.g.,
FOXL2-
AROM-
ERα,
RSPO1-
WNT4, and maybe unverified W genes) [
7,
12,
13]. Theoretically, genes on the W chromosome are likely to regulate female gonadal development in chickens.
Consequently, we explored the possibility that one of the genes on the W chromosome, MIER3, is involved in female gonadal development in chickens. We found that MIER3 gene expression in gonads at both the mRNA and protein levels and its protein distribution in gonadal tissues correlated with the gonadal phenotype. Further studies revealed that MIER3 gene might promote female gonad development through EGR1 and αGSU genes. These findings suggest that the MIER3 genes are involved in gonadal development in chicken embryos, increasing our understanding of the role of chicken W chromosome genes in gonadal development.
To predict the potential functions of the chicken
MIER3 gene, we first performed a comparative analysis of the mRNA and amino acid sequences encoded by different transcripts of the Z and W copies of the chicken
MIER3 gene and the human
MIER3 transcript 1. Notably, owing to evolutionary differences, unlike the chicken
MIER3 gene, human
MIER3 is not on the sex chromosome (located on autosome 5). Nevertheless, the amino acid sequences of human MIER3 (isoform 1) and chicken MIER3 (especially the Z-copy isoform) are generally similar, and both have three conserved structural domains, namely SANT, SANT-MTA3-like, and ELM2 [
14,
15,
16,
17], suggesting that chicken MIER3 may have functions similar to those of human MIER3. A study on human MIER3 showed that NAT9 (N-acetyltransferase 9) is a candidate for interacting with MIER3 [
18]. Breast cancer studies have revealed that
MIER3 is highly expressed in breast cancer, suggesting that human
MIER3 is closely associated with the development of breast cancer [
19]. Furthermore, human
MIER3 is reportedly lowly expressed in colorectal cancer cells, which may correlate with the degree of tumor differentiation, suggesting that
MIER3 is closely associated with colorectal carcinogenesis [
2]. However, no study has reported the function of chicken
MIER3. Notably, compared with human
MIER3, multiple transcripts of
MIER3–W (1,2,3,7) have an extra segment of amino acid sequence at the N-terminus (
Figure 1), and the phylogenetic tree revealed (
Supplementary Figure S4) that chicken
MIER3–W and
MIER3–Z are not in the closest branch, suggesting that
MIER3–W may have some specific functions.
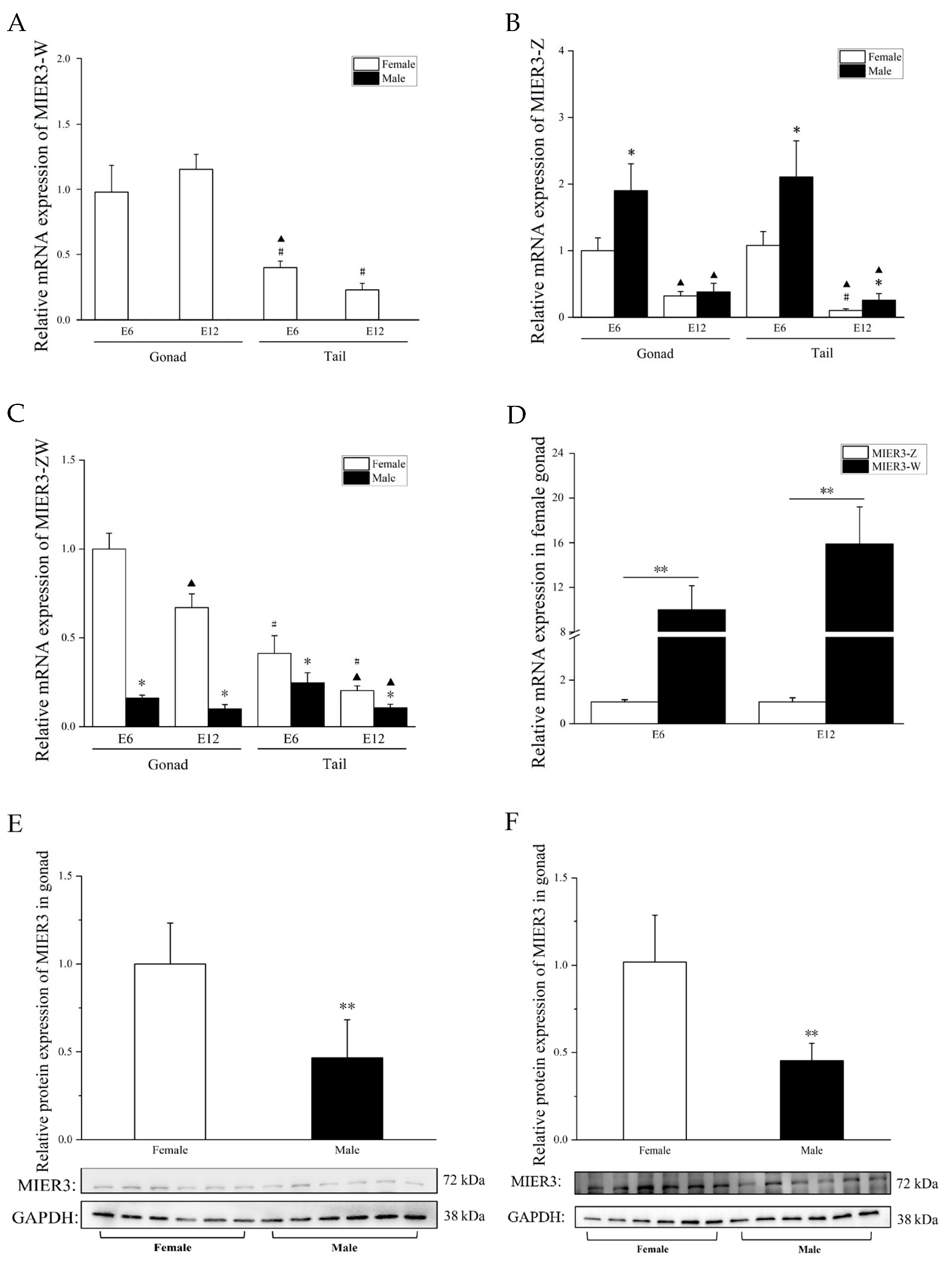
Additional evidence comes from the specific expression pattern of
MIER-W in the chicken embryonic gonads.
MIER3–W was significantly more expressed in chicken embryonic gonads than in other measured tissues (
Figure 2A and
Figure S3A,D);
MIER3–Z was not (
Figure 2B and
Figure S3B,E), suggesting that
MIER3–W may have certain functions related to gonadal development. Furthermore,
MIER3–W expression was significantly higher than
MIER3–Z in E6 and E12 chicken embryo gonads (
Figure 2D), suggesting that
MIER3–W is not just compensation for
MIER3–Z. This result is in line with the previous RNA-seq data of the W genes and contradicts the previous report that the W chromosome gene compensates for the Z chromosome copy [
2]. However, consistent with previous reports, there was a dosage effect of
MIER3–Z between males and females in most embryonic tissues [
2]. For example,
MIER3–Z was higher in males than in females in both E6 and E12 gonadal and tail tissues (
Figure 1B) and was also higher in males than in females in multiple non-gonadal tissues (E6: cerebrum; E12: liver, kidney, cerebrum, leg muscle, glandular stomach, and intestine).
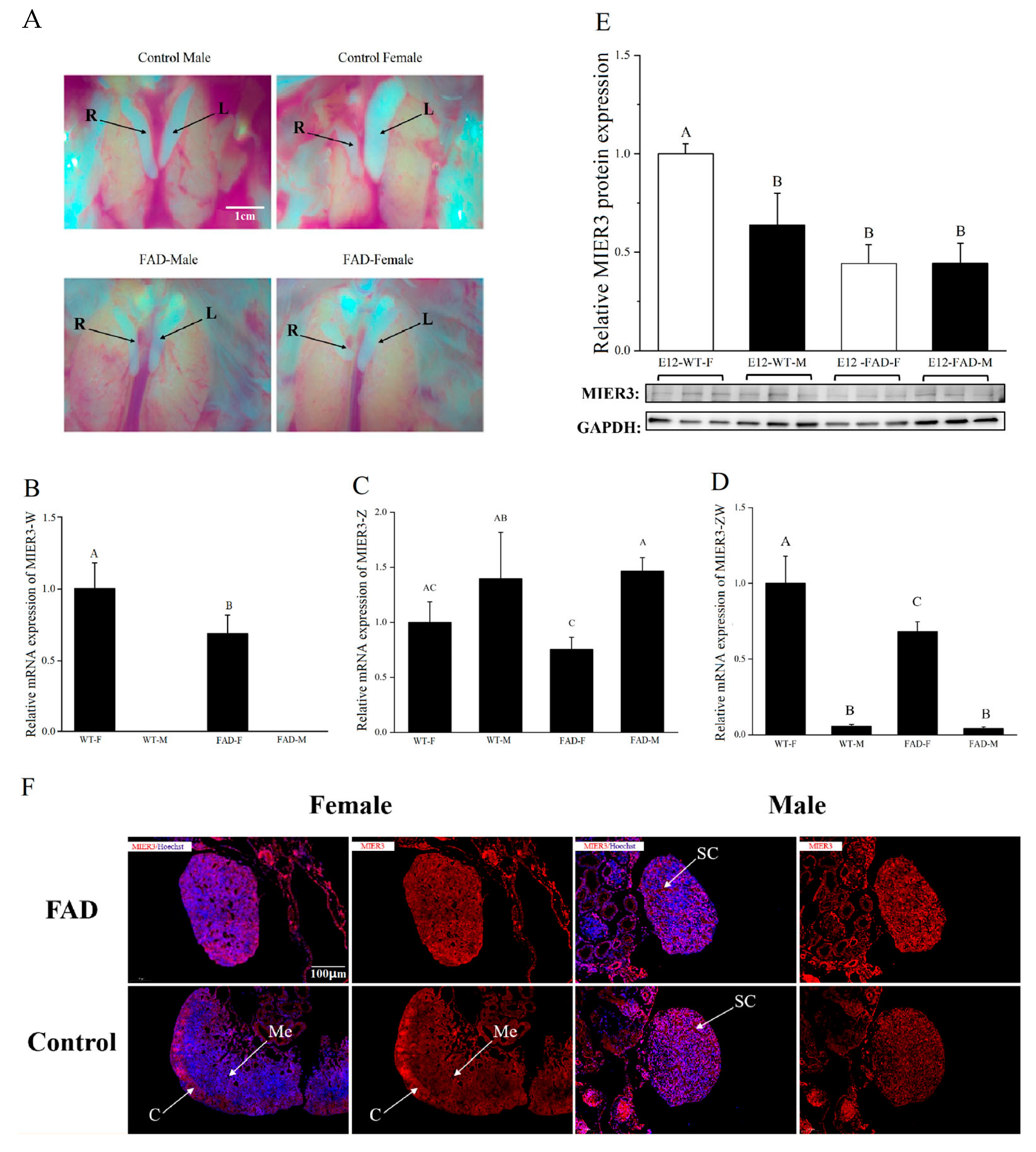
Notably, in the gonads of FAD-treated female embryos, the mRNA expression levels of both MIER3–W and MIER3–ZW were reduced, and a significant reduction was observed in protein levels. Additionally, FAD treatment affected MIER3 protein distribution in gonadal cells (from high expression in the female cortex to uniform expression in the medulla), suggesting that the MIER3 gene is closely related to the gonadal phenotype.
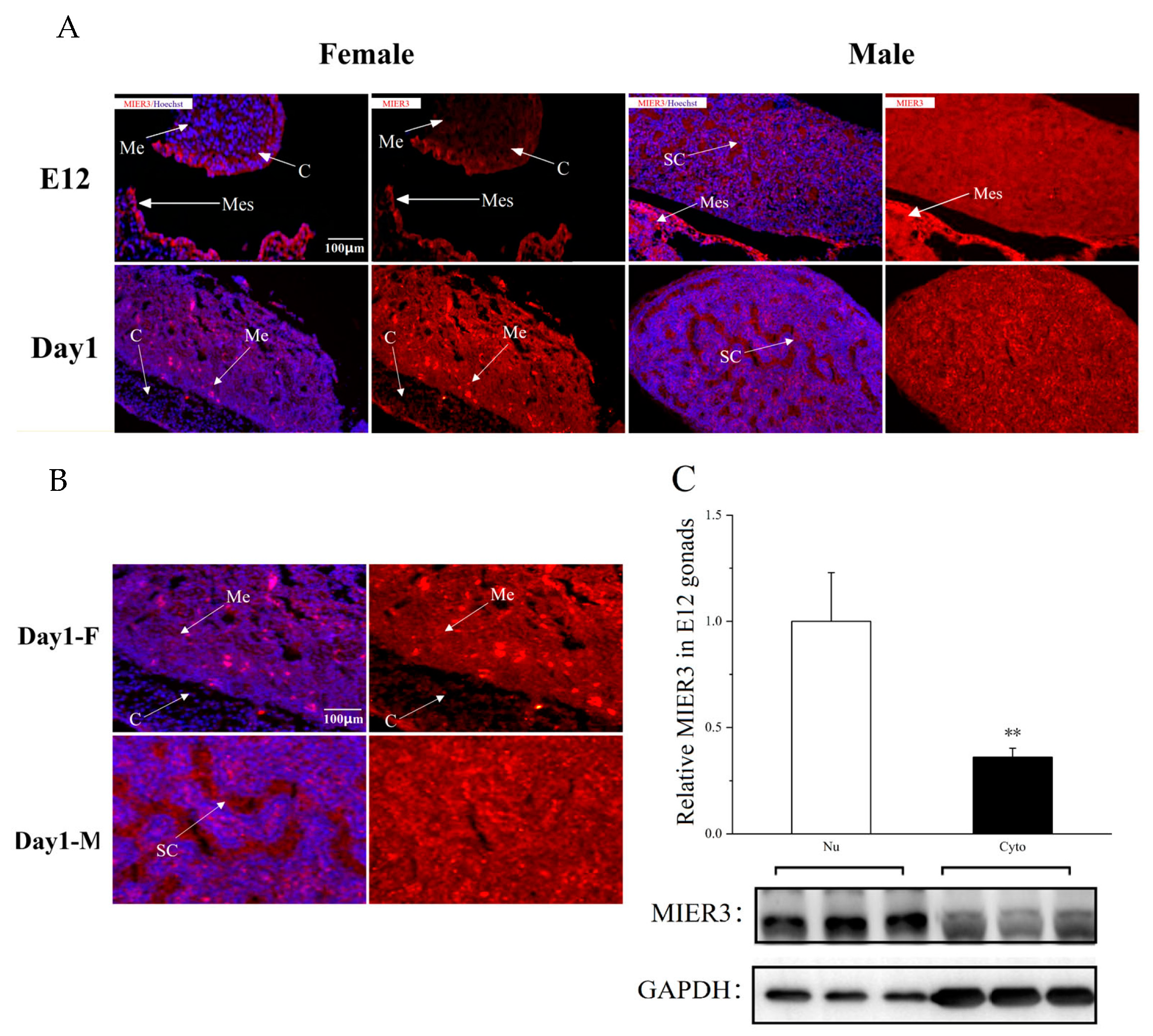
Furthermore, we used IF (Immunofluorescence) to determine the cellular localization of MIER3 protein in male and female embryonic gonads at different developmental stages (E12 and Day 1).
MIER3 gene had different distribution patterns of protein localization in the female and male gonads. In E12 embryos, MIER3 protein was highly expressed in the cortical layer of female gonads and uniformly expressed in the medullary layer of male gonads; in Day 1, it was highly expressed in the medullary layer of female gonads, with very low expression in the cortical layer. Since it has been demonstrated that
MIER3 can affect cell proliferation [
20], its high expression in the cortex of female gonads at early stages may contribute to the maintenance of the female cortex.
TGIF1 may be involved in this process, as it was identified as a regulator of proximal cortical medulla development [
21]. Our transcriptome sequencing results showed that
MIER3 overexpression increased
TGIF1 gene expression (
Supplementary Table S4). Furthermore, MIER3 protein is expressed in the nucleus and cytoplasm, with high levels in the nucleus, which is consistent with the results of Roya Derwish and Man Peng [
17,
22], proving that MIER3 is a nuclear protein. However, the MIER3 protein was expressed intracellularly and extracellularly, which was quite obvious in male gonads (
Figure 3B), suggesting that MIER3 may also function as a secretory protein.
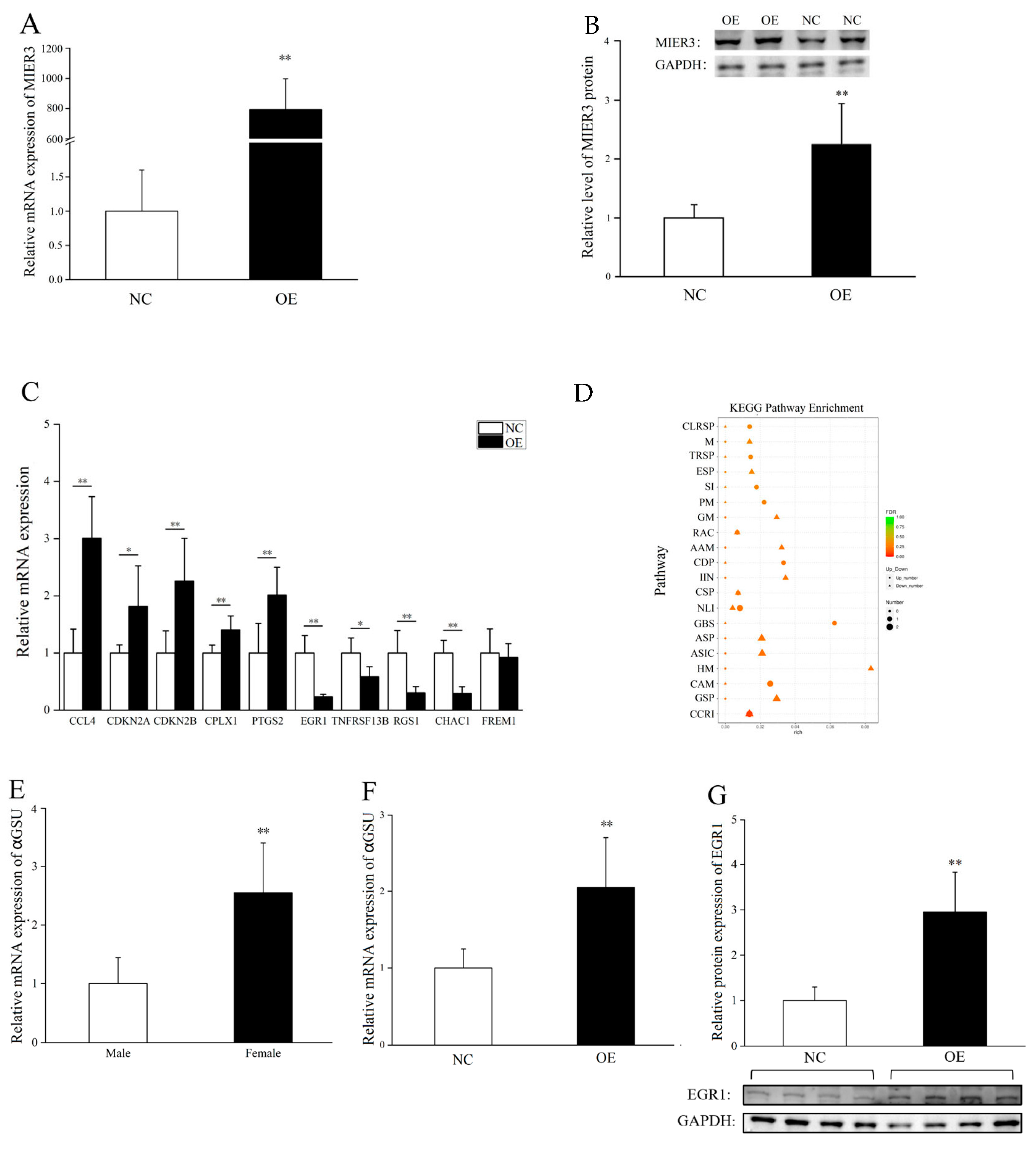
To explore MIER3–related signaling pathways and genes, we overexpressed MIER3–W in E9.5 male gonadal cells. With transcriptome sequencing, we screened a series of differential genes and signaling pathways, especially the GnRH signaling pathway (GnRH secreted by the hypothalamus acts on receptors in the anterior pituitary to regulate αGSU, LH, and FSHβ production and release). We found that MIER3 overexpression can significantly increase EGR1 protein and αGSU (a gene downstream of EGR1) mRNA expression. Furthermore, αGSU was significantly higher in female gonads than in male gonads at E6, suggesting that the MIER3 gene may promote the development of female gonads through EGR1 and αGSU. However, there was a contradiction between the mRNA and EGR1 protein levels in the gonadal cell’s MIER3 overexpression, indicating the existence of post-transcriptional regulation of EGR1. The detailed mechanism of this process warrants further study. In addition, we noticed that the overexpression of MIER3–W had no effect on E2 pathway genes (FOXL2, AROM, and ERα). Given the fact that FAD treatment (aromatase inhibition) decreased the expression of MIER3, it is suggested that MIER3 expression is downstream of the E2 pathway.
4. Materials and Methods
4.1. Animal Ethics Statement
All animal protocols were approved by the animal welfare committee of the Yangzhou University [permission number: SYXK(Su) IACUC 2012–0029] and comply with the associated guidelines.
4.2. Animal and Sample Collection
The eggs were all obtained from Hyline grey laying hens from Conway Farm (Yangzhou, China) and incubated in the laboratory of Yangzhou University. They were incubated at 37.5 °C with 60% humidity, blunt end up, and rotated every 30 min until the desired embryonic stages. On E6 (n = 6), E12 (n = 6), and hatch (gonadal morphology fully developed, n = 6), the eggs were removed from the incubator, and the embryos were carefully dissected to expose and collect the gonads and other tissues. The gonads and tails of E6 and E12 embryos were collected into 1.5 mL Eppendorf tubes for RNA extraction with TRIzol (Cat. No. DP424; Tiangen Biotech Co., Ltd., Beijing, China) and 200 μL of RIPA Lysis buffer (Cat. No. C1053; Applygen Technologies Co., Ltd., Beijing, China) for protein extraction. The complex renal tissues (gonad and mesonephros) were collected from the gonads of E12 and hatch, fixed in 4% paraformaldehyde, and subjected to paraffin sectioning and immunostaining.
4.3. Building a Phylogenetic Tree
Using MEGA 5.05 software, phylogenetic trees of amino acid sequences of MIER3 genes from different animals were constructed using the adjacency method, including: chicken (XP_004937308.1, XP_015155926.1), homo sapiens (XP_011541519.1), Mus musculus (NP_766181.2), Bos taurus (NP_001291684.1), Sus scrofa (XP_013840248.1), Canis lupus familiaris (XP_038315053.1), Equus caballus (XP_023481416.1), Felis catus (XP_023112535.1), Oryctolagus cuniculus (XP_008260452.1), Chelonia mydas (XP_037758759.1), Protopterus annectens (XP_043919732.1), Trachemys scripta elegans (XP_034630109.1) Lagopus leucura (XP_042731061.1), Cygnus olor (XP_0403955799.1), Hirundo rustica (XP_039946523.1), Oxyura jamaicensis (XP_035166467.1), Zootoca vivipara (XP_034956846.1), Phocoena sinus (XP_032484360.1), Aythya fuligula (XP_032062173.1), Anas platyrhynchos (XP_038025982.1), and anser cygnoides (XP_047905371.1).
4.4. FAD Treatment of Chicken Embryos
E2.5 (Embryonic day 2.5) embryos were injected with FAD, an aromatase inhibitor that causes gonadal sex reversal in female embryos [
23]. A small hole was made in the blunt end of the egg, and 0.2 mg FAD dissolved in phosphate buffer solution (PBS) was injected into the air sac. The eggs were then sealed and re-incubated until E12. Next, gonad samples were collected for RNA (n = 6) or protein (n = 6) extraction, as described above.
4.5. Determination of Genetic Sex of Chicken Embryos
DNA was extracted from chicken embryonic tissues (wings or toes) using a commercial kit (Cat. No. DC102-01; Novozyme Biotechnology Co., Ltd., Nanjing, China). Subsequently, PCR was performed to amplify the CHD gene (Chromobox-helicase-DNA binding gene) sequence located on both sex chromosomes. The CHD-forward/reward primer sequences were as follows: CHD-F, AGTGCATTGCAGAAGCAATATT; CHD-R, GCCTCCTGTTTATTATAGAATTCAT. The female (ZW) had two bands at 506 bp and 351 bp, and the male (ZZ) had only one band at 506 bp.
4.6. RNA Isolation, cDNA Synthesis, and qPCR
Total RNA was extracted from the gonads and other tissues using TRIzol universal total RNA extraction reagent (Cat. No. DP424; Tiangen Biotech Co., Ltd.) according to the manufacturer’s instructions. For embryos at E6, five pairs of gonads were pooled for RNA isolation (6 pools for each sex), and for embryos at other stages (E12 and hatch) and in cell experiments, total RNA was extracted from six pairs of gonads of each sex (n = 6). The HiScript Q RT SuperMix for qPCR reverse transcription kit (Cat. No. R123-01; Novozyme Biotechnology Co., Ltd., Nanjing, China) was used for cDNA synthesis. Quantitative PCR was performed using a VazymeAceQ qPCR SYBR Green Master Mix kit (Cat. No. Q111- 02/03; Vazyme Biotech Co., Ltd., Nanjing, China), according to the manufacturer’s instructions. The hydroxymethylbilane synthase gene was used as a control for the spatiotemporal expression of the
MIER3 gene in chicken embryos [
24], and the actin (
β-actin) gene was used as a control for the rest of the assays. The primer sequences used for qPCR analysis are listed in
Supplementary Table S1.
4.7. Protein Extraction and Western Blotting
Nuclear and cytoplasmic protein fractions were prepared from E12 female gonads (n = 3) using the “NE-PER Nucleus and Cytoplasm Extraction Reagent” kit (Cat. No. 78833; Thermo Scientific Co., Ltd., Boston, MA, USA), according to the manufacturer’s instructions. Total protein from the gonads was extracted using RIPA buffer (Cat. No. C1053; Applygen Technologies Co., Ltd., Beijing, China) according to the manufacturer’s instructions, and protein concentrations in the samples were determined using the Enhanced BCA Protein Analysis Kit (Cat. No. P0010; Beyotime, Shanghai, China). Immunoblot analysis was performed as described previously [
25]. Briefly, 4 μg of protein sample was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to PVDF (polyvinylidene fluoride) membranes. After sealing, the membranes were incubated with the primary antibody overnight at 4°C and then incubated with secondary antibody for 1 h at room temperature (15–25 °C) before imaging. Scanned images were analyzed using Image Studio Lite Ver 5.2. MIER3 antibody (bs-49157R) (Boosun Biotechnology Co., Ltd., Beijing, China), EGR1 antibody (ab133695, Abcam, Cambridge, UK), and GAPDH antibody (bs-0755R, Boosun Biotechnology Co., Ltd., Beijing, China) were used as a control.
4.8. Paraffin Sectioning and Immunostaining
For histological analysis, the complex renal tissues were placed in 4% paraformaldehyde for 24 h. Next, the tissue shape was adjusted under a stereomicroscope. Subsequently, the tissue and corresponding labels were placed in a dehydration cassette, dehydrated with low to high ethanol concentrations, treated with xylene, and cleared for paraffin embedding. The embedded tissues were serially sliced to a thickness of 3 μm. Next, the slices were floated on a spreader of 40 °C warm water to spread the tissues. After, the tissues were fished up with slides, baked in an oven at 60 °C, dried, and removed for storage at room temperature. Immunohistochemistry was performed as previously described [
26]. Briefly, the slides were washed in PBS at 37 °C for 30 min and closed in PBS containing 10% donkey serum, 1% BSA (Bovine Serum Albumin), and 0.3% Triton X-100 at 24 °C for 2 h. The slides were incubated with primary antibody overnight at 4 °C, washed in PBS containing 0.3% Triton X-100 before incubation, and then incubated with secondary antibody at room temperature for 2 h. Lastly, the slides were washed in PBS containing 0.3% Triton X-100, and sections were treated with Hoechst solution (10 mg/mL) for 5 min to stain cell nuclei. The working concentrations of the antibodies used were as follows: MIER3 antibody, 5 mg/mL; and Hoechst 33,342 (Cat. No. C1025; Beyotime Biotechnology Co., Ltd., Shanghai, China), 1:100.
4.9. Construction of MIER3–W Overexpression Vector
Based on the coding region of chicken MIER3–W (NCBI gene ID: 426615, accession: XM_015300440.3), the MIER3–W overexpression plasmid was designed by Jima Genetics Co., Ltd., Shanghai, China. The plasmid was lentivirally packaged by Kinkai Rui Biological Engineering Co., Ltd., Wuhan, China.
4.10. Isolation and Processing of Chicken Embryo Gonadal Cells
On E9.5 (right after sex determination, early stage of gonadal development, n = 6), cells from the female and male gonads were isolated from chicken embryos using collagenase (Cat. No. 17104019; Gibco). Briefly, E9.5 chicken embryo gonads were collected in 1.5 mL centrifuge tubes with PBS. After aspirating and discarding PBS, 500 μL of pre-warmed collagenase was added, and the tissues were pipetted and heated at 37 °C for 30 s. This step was repeated until the gonads became invisible. Subsequently, 800 μL of 10% bovine serum (FBS)-Dulbecco’s Modified Eagle Medium (DMEM) was added to the digestion solution, mixed by pipetting, and centrifuged at 700× g for 5 min at room temperature. After aspirating and discarding the supernatant, we added 1 mL of PBS to resuspend and wash the cells, which were centrifuged at 700× g for 5 min at room temperature. Next, the supernatant was discarded, and 1 mL of complete medium (DMEM with 10% fetal bovine serum, 1% penicillin-streptomycin solution (100 IU/mL), and 10 μL EGF (20 ng/mL) was added, and the mixture was filtered into a 50 mL conical tube using 70 μm nylon mesh. After counting the cells, a complete medium was added to plate and culture the cells at 1 × 106 cells/well. The cells were then transfected with MIER3 overexpression plasmid and empty vector plasmid at 70% cell fusion using Lipofectamine 2000 (Cat. No. 11668-027, Invitrogen, Carlsbad, CA, USA). Briefly, 4 μL of Lipofectamine 2000, MIER3 overexpression plasmid and empty vector plasmid were diluted with 96 μL Opti-MEM. Next, the diluted Lipofectamine 2000 was mixed with diluted MIER3 overexpression plasmid and empty vector and left to stand at room temperature for 23 min. The mixture was then added to the cultured cells. After 48 h of transfection, the cells were washed twice with PBS and then collected with 1 mL of TRIzol reagent or 100 μL of RIPA lysate and subjected to phenotypic assays.
4.11. Transcriptome Sequencing
Total RNA was extracted from cells transfected with MIER3 overexpression vector or empty vector (n = 3) and sent to Personal Biotechnology Co., Ltd. for transcriptome sequencing. The quantity and quality of total RNA samples were analyzed with a Qubit 2.0 (ThermoFisher Scientific Inc., Waltham, MA, USA) and Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA), respectively. The qualified total RNA samples were subsequently used to construct cDNA libraries by reverse transcription. The quality of this library was checked and sequenced using synthetic technology on the Illumina high-throughput sequencing platform HiSeq X-ten. Clean data with a score of ≥Q30 were selected for further analysis based on the calculated Phred scores of a large amount of raw data. The clean data were annotated using the HISAT2 system by comparing the reference genome sequence of chickens. Individual genes were assembled by comparing the reads using the StringTie program. Next, the expression of individual genes was calculated by normalizing the fragment per kilobase transcript/fragment per million map maps (FPKM) reads. Differential gene expression analysis was performed using DESeq (1.20.0) to screen for differentially expressed genes under the following conditions: expression difference fold |log2FoldChange| > 1 and significance p < 0.05. Gene ontology (GO) enrichment analysis was performed using top GO, and the p was calculated by the hypergeometric distribution method (the criterion for significant enrichment was p < 0.05). GO terms with significant enrichment of differential genes were identified to determine the main biological functions of the differential genes. KEGG pathway enrichment analysis was performed using clusterProfiler (3.4.4) software, focusing on significantly enriched pathways with p < 0.05.
4.12. Cell Phenotype
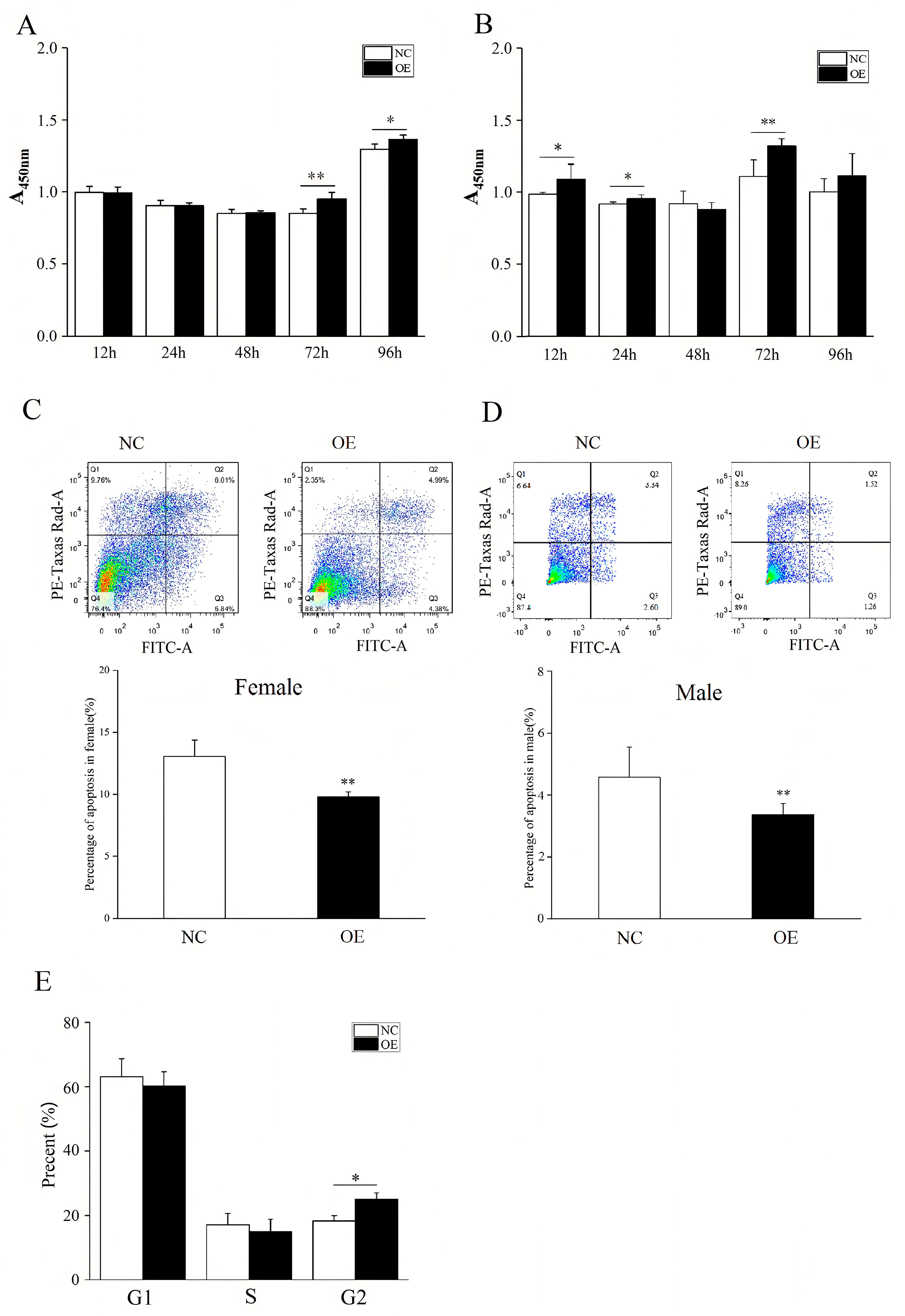
The proliferative ability of chicken embryonic gonadal cells was assessed using CCK-8 (Cat. No. A311-02; Vazyme Biotech Co., Ltd.) according to the manufacturer’s instructions. The cells transfected with MIER3 overexpression vector or negative control vector (n = 4) were inoculated in 96-well plates (100 μL/well) at 2 × 103/well, and five replicate wells were set up for each group. The cells were incubated for 12, 24, 48, 72, and 96 h with 10 μL CCK-8 in each well, and the absorbance (A450 nm) value of each well was measured at 450 nm by enzyme marker after incubation for 2 h at 37 °C. The background A450 nm value was removed by zeroing the blank control group, and the cell growth curve was statistically analyzed and plotted. Flow cytometry was used to detect apoptosis in chicken embryonic gonadal cells, and MIER3 overexpression vector or negative control vector was transfected according to the instructions of the cell Apoptosis Kit (Cat. No. A211; Novozyme Biotechnology Co., Ltd. Nanjing, China). Untreated cells were used as a negative control. Additionally, for flow cytometry detection, annexin V-FITC and propidium iodide (PI) monochromes were required to regulate compensation. After 48 h, cells in each group were digested with trypsin without EDTA and collected. Next, the cells were washed twice with precooled PBS, and then 100 µL of 1× binding buffer was added and gently blown into a single-cell suspension. In addition, annexin V-FITC and PI staining were performed on the samples for flow cytometry detection to regulate compensation. After transfection with the MIER3 overexpression vector for 48 h, the cells in each group were digested with trypsin without EDTA and collected. The cells were washed twice with precooled PBS and then gently blown into a single-cell suspension with 100 µL of 1× binding buffer. Subsequently, 5 μL Annexin V-FITC and 5 μL PI staining solution were added at room temperature (20–25 °C) for 10 min, followed by 400 μL of 1× binding buffer, and the mixture was blended gently. The samples were analyzed by flow cytometry within 1 h of staining, and the apoptosis distribution map was statistically analyzed and drawn. According to the manufacturer’s instructions for the kit (Cat. No. C1052; Beyotime Biotechnology Co., Ltd., Shanghai, China), the cell cycle of chicken embryonic gonad cells was detected by flow cytometry. Briefly, the cells in each group were collected by trypsin digestion after transfection with the MIER3 overexpression and negative control vectors for 48 h. After 1 mL of precooled PBS was added to resuspended cells, 1 mL of 70% ethanol was added to fix the cells at 4 °C for 12 h, and then precooled 1 mL PBS was added to resuspended cells. Lastly, 0.5 mL of PI staining solution was added to the pre-prepared. After the resuspended cells were precipitated and incubated at 37 °C in the dark for 30 min, the cell cycle distribution was detected by flow cytometry.
4.13. Statistical Analysis
All values are expressed as the mean ± standard deviation. SPSS (version 16.0, SPSS China, Shanghai, China) was used to perform a Student’s t-test or one-way analysis of variance (Duncan’s test) to test for statistical significance differences between or among different groups. Statistical significance was set at p < 0.05.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





