
Genetic Variants within SARS-CoV-2 Human Receptor Genes May Contribute to Variable Disease Outcomes in Different Ethnicities


Abstract
:1. Introduction
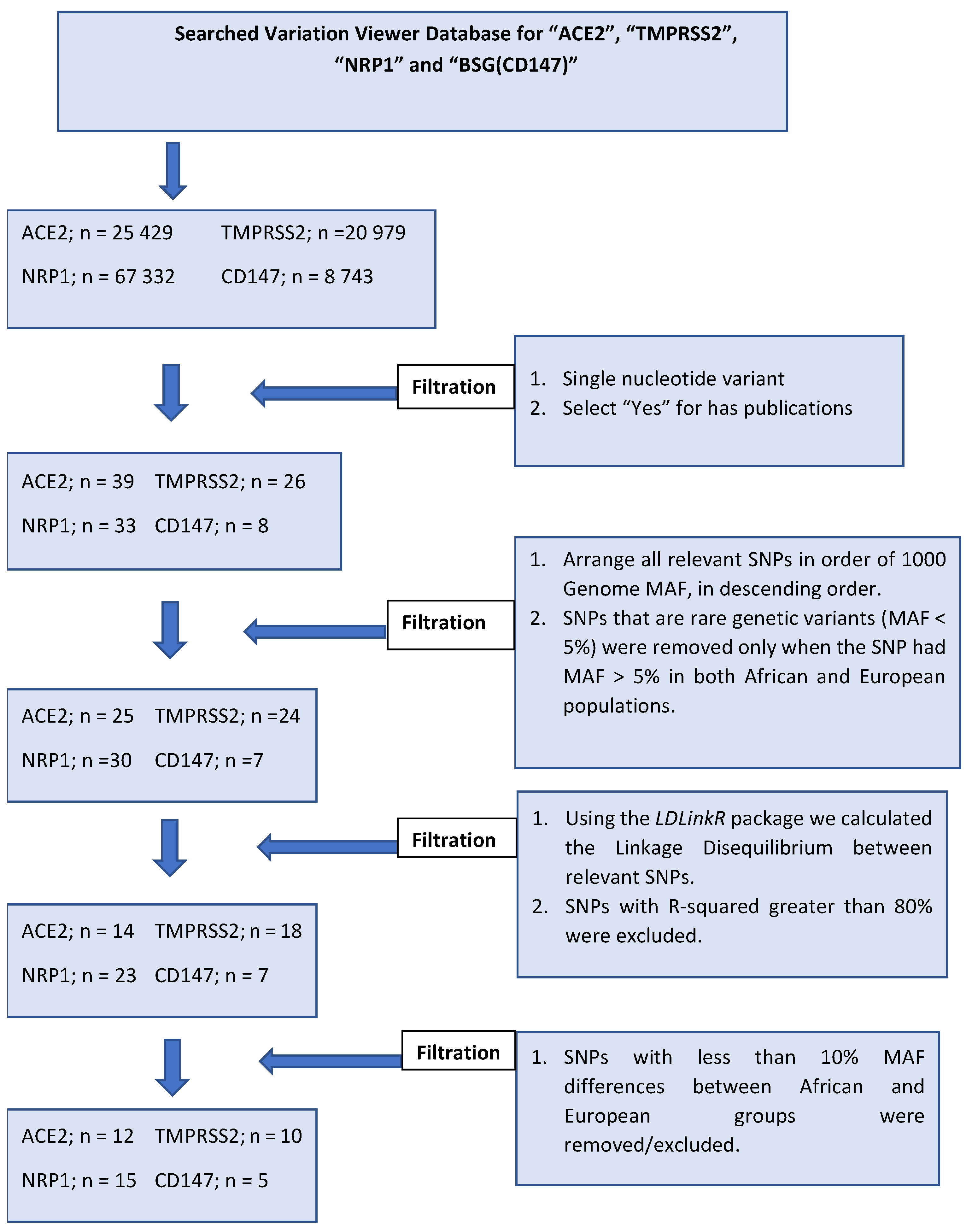
1.1. Inclusion and Exclusion Criteria for Selected SNPs
1.2. Excess Deaths Reported between January 2020 and December 2021 of the COVID-19 Pandemic Support the Theory of the “African Paradox”
1.3. SARS-CoV-2 Receptors, Physiological Significance, and Significant Regulatory SNPs
1.3.1. ACE2
1.3.2. TMPRSS2
1.3.3. NRP1
1.3.4. BSG (CD147)
1.4. Addressing the African Paradox
1.5. SARS-CoV-2 Receptor-Based Therapy and Clinical Significance
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patel, R.; Kaki, M.; Potluri, V.S.; Kahar, P.; Khanna, D. A comprehensive review of SARS-CoV-2 vaccines: Pfizer, Moderna & Johnson & Johnson. Hum. Vaccines Immunother. 2022, 18, 2002083. [Google Scholar]
- Rodriguez, E.V.; Bouazza, F.-Z.; Dauby, N.; Mullier, F.; D’otreppe, S.; Jissendi Tchofo, P.; Bartiaux, M.; Sirjacques, C.; Roman, A.; Hermans, C. Fatal vaccine-induced immune thrombotic thrombocytopenia (VITT) post Ad26. COV2. S: First documented case outside US. Infection 2022, 50, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, B.; Zhang, S.; Huang, N.; Zhao, T.; Lu, Q.B.; Cui, F. Differences in incidence and fatality of COVID-19 by SARS-CoV-2 Omicron variant versus Delta variant in relation to vaccine coverage: A world-wide review. J. Med. Virol. 2023, 95, e28118. [Google Scholar] [CrossRef] [PubMed]
- Samson, M.; Libert, F.; Doranz, B.J.; Rucker, J.; Liesnard, C.; Farber, C.-M.; Saragosti, S.; Lapouméroulie, C.; Cognaux, J.; Forceille, C. Resistance to HIV-1 infection in caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature 1996, 382, 722–725. [Google Scholar] [CrossRef] [PubMed]
- De Silva, E.; Stumpf, M.P.H. HIV and the CCR5-Δ32 resistance allele. FEMS Microbiol. Lett. 2004, 241, 1–12. [Google Scholar] [CrossRef]
- Martinson, J.J.; Chapman, N.H.; Rees, D.C.; Liu, Y.-T.; Clegg, J.B. Global distribution of the CCR5 gene 32-basepair deletion. Nat. Genet. 1997, 16, 100–103. [Google Scholar] [CrossRef]
- Lederman, M.M.; Pike, E. Ten Years HIV Free: An Interview with “The Berlin Patient,” Timothy Ray Brown. Pathog. Immun. 2017, 2, 422–430. [Google Scholar]
- Brown, T.R. I am the Berlin patient: A personal reflection. AIDS Res. Hum. Retrovir. 2015, 31, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Peppa, D.; Hill, A.L.; Gálvez, C.; Salgado, M.; Pace, M.; Mccoy, L.E.; Griffith, S.A.; Thornhill, J.; Alrubayyi, A. Evidence for HIV-1 cure after CCR5Δ32/Δ32 allogeneic haemopoietic stem-cell transplantation 30 months post analytical treatment interruption: A case report. Lancet HIV 2020, 7, e340–e347. [Google Scholar] [CrossRef]
- Li, H.; Liu, Z.; Han, Q.; Li, Y.; Chen, J. Association of genetic polymorphism of low-density lipoprotein receptor with chronic viral hepatitis C infection in Han Chinese. J. Med. Virol. 2006, 78, 1289–1295. [Google Scholar]
- Harada, R.; Kimura, M.; Sato, Y.; Taniguchi, T.; Tomonari, T.; Tanaka, T.; Tanaka, H.; Muguruma, N.; Shinomiya, H.; Honda, H. APOB codon 4311 polymorphism is associated with hepatitis C virus infection through altered lipid metabolism. BMC Gastroenterol. 2018, 18, 24. [Google Scholar] [CrossRef]
- Naga, M.; Amin, M.; Algendy, D.; Elbadry, A.; Fawzi, M.; Foda, A.; Esmat, S.; Sabry, D.; Rashed, L.; Gabal, S.; et al. Low-density lipoprotein receptor genetic polymorphism in chronic hepatitis C virus Egyptian patients affects treatment response. World J. Gastroenterol. 2015, 21, 11141–11151. [Google Scholar] [CrossRef]
- Hu, H.H.; Liu, J.; Lin, Y.L.; Luo, W.S.; Chu, Y.J.; Chang, C.L.; Jen, C.L.; Lee, M.H.; Lu, S.N.; Wang, L.Y.; et al. The rs2296651 (S267F) variant on NTCP (SLC10A1) is inversely associated with chronic hepatitis B and progression to cirrhosis and hepatocellular carcinoma in patients with chronic hepatitis B. Gut 2016, 65, 1514–1521. [Google Scholar] [CrossRef]
- Chuaypen, N.; Tuyapala, N.; Pinjaroen, N.; Payungporn, S.; Tangkijvanich, P. Association of NTCP polymorphisms with clinical outcome of hepatitis B infection in Thai individuals. BMC Med. Genet. 2019, 20, 87. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Mayi, B.S.; Leibowitz, J.A.; Woods, A.T.; Ammon, K.A.; Liu, A.E.; Raja, A. The role of Neuropilin-1 in COVID-19. PLoS Pathog. 2021, 17, e1009153. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, W.; Zhang, Z.; Deng, Y.; Lian, J.-Q.; Du, P.; Wei, D.; Zhang, Y.; Sun, X.-X.; Gong, L. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct. Target. Ther. 2020, 5, 283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, C.Z.; Swaroop, M.; Xu, M.; Wang, L.; Lee, J.; Wang, A.Q.; Pradhan, M.; Hagen, N.; Chen, L.; et al. Heparan sulfate assists SARS-CoV-2 in cell entry and can be targeted by approved drugs in vitro. Cell Discov. 2020, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Vahidy, F.S.; Nicolas, J.C.; Meeks, J.R.; Khan, O.; Pan, A.; Jones, S.L.; Masud, F.; Sostman, H.D.; Phillips, R.; Andrieni, J.D. Racial and ethnic disparities in SARS-CoV-2 pandemic: Analysis of a COVID-19 observational registry for a diverse US metropolitan population. BMJ Open 2020, 10, e039849. [Google Scholar] [CrossRef]
- Delshad, M.; Sanaei, M.J.; Pourbagheri-Sigaroodi, A.; Bashash, D. Host genetic diversity and genetic variations of SARS-CoV-2 in COVID-19 pathogenesis and the effectiveness of vaccination. Int. Immunopharmacol. 2022, 111, 109128. [Google Scholar] [CrossRef]
- Bwire, G.; Ario, A.R.; Eyu, P.; Ocom, F.; Wamala, J.F.; Kusi, K.A.; Ndeketa, L.; Jambo, K.C.; Wanyenze, R.K.; Talisuna, A.O. The COVID-19 pandemic in the African continent. BMC Med. 2022, 20, 167. [Google Scholar] [CrossRef]
- WHO. WHO Coronavirus (COVID-19) Dashboard|WHO Coronavirus (COVID-19) Dashboard with Vaccination Data; WHO: Geneva, Switzerland, 2021; pp. 1–5. [Google Scholar]
- European Centre for Disease Prevention and Control (CDC). European Centre for Disease Prevention and Control (CDC) COVID-19 Situation Update Worldwide, as of 30 June 2020; European Centre for Disease Prevention and Control: Solna, Sweden, 2020. [Google Scholar]
- Chitungo, I.; Dzobo, M.; Hlongwa, M.; Dzinamarira, T. COVID-19: Unpacking the low number of cases in Africa. Public Health Pract. 2020, 1, 100038. [Google Scholar] [CrossRef]
- Kircheis, R.; Schuster, M.; Planz, O. COVID-19: Mechanistic Model of the African Paradox Supports the Central Role of the NF-κB Pathway. Viruses 2021, 13, 1887. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.; Mackenzie, M.J.; Amegah, A.K.; Ezeh, A.; Gadanya, M.A.; Omigbodun, A.; Sarki, A.M.; Thistle, P.; Ziraba, A.K.; Stranges, S. The conundrum of low COVID-19 mortality burden in sub-Saharan Africa: Myth or reality? Glob. Health Sci. Pract. 2021, 9, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, Y.; Guan, T.; Lai, Y.; Shen, Y.; Zeyaweiding, A.; Zhao, H.; Li, F.; Maimaiti, T. ACE2 polymorphisms associated with cardiovascular risk in Uygurs with type 2 diabetes mellitus. Cardiovasc. Diabetol. 2018, 17, 127. [Google Scholar] [CrossRef]
- Lieb, W.; Graf, J.; Götz, A.; König, I.R.; Mayer, B.; Fischer, M.; Stritzke, J.; Hengstenberg, C.; Holmer, S.R.; Döring, A.; et al. Association of angiotensin-converting enzyme 2 (ACE2) gene polymorphisms with parameters of left ventricular hypertrophy in men. Results of the MONICA Augsburg echocardiographic substudy. J. Mol. Med. 2006, 84, 88–96. [Google Scholar] [CrossRef]
- Zhao, Q.; Hixson, J.E.; Rao, D.C.; Gu, D.; Jaquish, C.E.; Rice, T.; Shimmin, L.C.; Chen, J.; Cao, J.; Kelly, T.N.; et al. Genetic variants in the apelin system and blood pressure responses to dietary sodium interventions: A family-based association study. J. Hypertens. 2010, 28, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.H.; Wang, Y.B.; Wang, H.; Sun, K.; Zhang, W.L.; Song, X.D.; Cheng, J.Z.; Wu, H.Y.; Zhou, X.L.; Hui, R.T. Polymorphisms of angiotensin-converting enzyme (ACE) and ACE2 are not associated with orthostatic blood pressure dysregulation in hypertensive patients. Acta Pharmacol. Sin. 2009, 30, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Gu, D.; Kelly, T.N.; Hixson, J.E.; Rao, D.C.; Jaquish, C.E.; Chen, J.; Huang, J.; Chen, C.S.; Gu, C.C.; et al. Association of genetic variants in the apelin-APJ system and ACE2 with blood pressure responses to potassium supplementation: The GenSalt study. Am. J. Hypertens. 2010, 23, 606–613. [Google Scholar] [CrossRef]
- Lozano-Gonzalez, K.; Padilla-Rodríguez, E.; Texis, T.; Gutiérrez, M.N.; Rodríguez-Dorantes, M.; Cuevas-Córdoba, B.; Ramírez-García, E.; Mino-León, D.; Sánchez-García, S.; Gonzalez-Covarrubias, V. Allele Frequency of ACE2 Intron Variants and Its Association with Blood Pressure. DNA Cell Biol. 2020, 39, 2095–2101. [Google Scholar] [CrossRef]
- Srivastava, A.; Pandey, R.K.; Singh, P.P.; Kumar, P.; Rasalkar, A.A.; Tamang, R.; van Driem, G.; Shrivastava, P.; Chaubey, G. Most frequent South Asian haplotypes of ACE2 share identity by descent with East Eurasian populations. PLoS ONE 2020, 15, e0238255. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Hsu, Y.-W.; Chang, C.-M.; Wu, C.-C.; Ou, J.-C.; Tsai, Y.-R.; Chiu, W.-T.; Chang, W.-C.; Chiang, Y.-H.; Chen, K.-Y. The influence of BMX gene polymorphisms on clinical symptoms after mild traumatic brain injury. BioMed Res. Int. 2014, 2014, 293687. [Google Scholar] [PubMed]
- Meng, N.; Zhang, Y.; Ma, J.; Li, H.; Zhou, F.; Qu, Y. Association of polymorphisms of angiotensin I converting enzyme 2 with retinopathy in type 2 diabetes mellitus among Chinese individuals. Eye 2015, 29, 266–271. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Noroozi, R.; Vafaee, R.; Branicki, W.; Poṡpiech, E.; Pyrc, K.; Łabaj, P.P.; Omrani, M.D.; Taheri, M.; Sanak, M. Effects of host genetic variations on response to, susceptibility and severity of respiratory infections. Biomed. Pharmacother. 2020, 128, 110296. [Google Scholar] [CrossRef]
- Iyer, G.R.; Samajder, S.; Zubeda, S.; Devi, S.N.S.; Mali, V.; Pv, S.K.; Sharma, A.; Abbas, N.Z.; Bora, N.S.; Narravula, A.; et al. Infectivity and Progression of COVID-19 Based on Selected Host Candidate Gene Variants. Front. Genet. 2020, 11, 861. [Google Scholar] [CrossRef]
- Luostari, K.; Hartikainen, J.M.; Tengström, M.; Palvimo, J.J.; Kataja, V.; Mannermaa, A.; Kosma, V.M. Type II transmembrane serine protease gene variants associate with breast cancer. PLoS ONE 2014, 9, e102519. [Google Scholar] [CrossRef]
- Posadas-Sánchez, R.; Fragoso, J.M.; Sánchez-Muñoz, F.; Rojas-Velasco, G.; Ramírez-Bello, J.; López-Reyes, A.; Martínez-Gómez, L.E.; Sierra-Fernández, C.; Rodríguez-Reyna, T.; Regino-Zamarripa, N.E.; et al. Association of the Transmembrane Serine Protease-2 (TMPRSS2) Polymorphisms with COVID-19. Viruses 2020, 14, 1976. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhou, J.; To, K.K.; Chu, H.; Li, C.; Wang, D.; Yang, D.; Zheng, S.; Hao, K.; Bossé, Y.; et al. Identification of TMPRSS2 as a Susceptibility Gene for Severe 2009 Pandemic A(H1N1) Influenza and A(H7N9) Influenza. J. Infect. Dis. 2015, 212, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Haralambieva, I.H.; Dhiman, N.; Ovsyannikova, I.G.; Vierkant, R.A.; Pankratz, V.S.; Jacobson, R.M.; Poland, G.A. 2′-5′-Oligoadenylate synthetase single-nucleotide polymorphisms and haplotypes are associated with variations in immune responses to rubella vaccine. Hum. Immunol. 2010, 71, 383–391. [Google Scholar] [CrossRef]
- Senapati, S.; Kumar, S.; Singh, A.K.; Banerjee, P.; Bhagavatula, S. Assessment of risk conferred by coding and regulatory variations of TMPRSS2 and CD26 in susceptibility to SARS-CoV-2 infection in human. J. Genet. 2020, 99, 53. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.L.; Huang, W.; Tang, N.L.; Lam, L.C. MxA polymorphisms are associated with risk and age-at-onset in Alzheimer disease and accelerated cognitive decline in Chinese elders. Rejuvenation Res. 2012, 15, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Minashkin, M.M.; Grigortsevich, N.Y.; Kamaeva, A.S.; Barzanova, V.V.; Traspov, A.A.; Godkov, M.A.; Ageev, F.A.; Petrikov, S.S.; Pozdnyakova, N.V. The Role of Genetic Factors in the Development of Acute Respiratory Viral Infection COVID-19: Predicting Severe Course and Outcomes. Biomedicines 2022, 10, 549. [Google Scholar] [CrossRef] [PubMed]
- Lorés-Motta, L.; van Asten, F.; Muether, P.S.; Smailhodzic, D.; Groenewoud, J.M.; Omar, A.; Chen, J.; Koenekoop, R.K.; Fauser, S.; Hoyng, C.B.; et al. A genetic variant in NRP1 is associated with worse response to ranibizumab treatment in neovascular age-related macular degeneration. Pharm. Genom. 2016, 26, 20–27. [Google Scholar] [CrossRef]
- Grupe, A.; Li, Y.; Rowland, C.; Nowotny, P.; Hinrichs, A.L.; Smemo, S.; Kauwe, J.S.; Maxwell, T.J.; Cherny, S.; Doil, L.; et al. A scan of chromosome 10 identifies a novel locus showing strong association with late-onset Alzheimer disease. Am. J. Hum. Genet. 2006, 78, 78–88. [Google Scholar] [CrossRef]
- Nejentsev, S.; Smink, L.J.; Smyth, D.; Bailey, R.; Lowe, C.E.; Payne, F.; Masters, J.; Godfrey, L.; Lam, A.; Burren, O.; et al. Sequencing and association analysis of the type 1 diabetes-linked region on chromosome 10p12-q11. BMC Genet. 2007, 8, 24. [Google Scholar] [CrossRef]
- Fan, S.H.; Shen, Z.Y.; Xiao, Y.M. Functional polymorphisms of the neuropilin 1 gene are associated with the risk of tetralogy of Fallot in a Chinese Han population. Gene 2018, 653, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Thornton-Wells, T.A.; Moore, J.H.; Martin, E.R.; Pericak-Vance, M.A.; Haines, J.L. Confronting complexity in late-onset Alzheimer disease: Application of two-stage analysis approach addressing heterogeneity and epistasis. Genet. Epidemiol. 2008, 32, 187–203. [Google Scholar] [CrossRef]
- Hong, J.M.; Kim, T.H.; Kim, H.J.; Park, E.K.; Yang, E.K.; Kim, S.Y. Genetic association of angiogenesis- and hypoxia-related gene polymorphisms with osteonecrosis of the femoral head. Exp. Mol. Med. 2010, 42, 376–385. [Google Scholar] [CrossRef]
- Weng, Y.; Chen, T.; Ren, J.; Lu, D.; Liu, X.; Lin, S.; Xu, C.; Lou, J.; Chen, X.; Tang, L. The Association Between Extracellular Matrix Metalloproteinase Inducer Polymorphisms and Coronary Heart Disease: A Potential Way to Predict Disease. DNA Cell Biol. 2020, 39, 244. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-S.; Li, F.-F.; Sun, L.-D.; Li, D.; Su, J.; Kuang, Y.-H.; Chen, G.; Chen, X.-P.; Chen, X. A miRNA-492 binding-site polymorphism in BSG (basigin) confers risk to psoriasis in central south Chinese population. Hum. Genet. 2011, 130, 749–757. [Google Scholar] [CrossRef]
- Jin, W.; Wu, W.; Yang, K.; Shen, F.; Fu, N.; Feng, Y.; Fu, Y. The Single Nucleotide Polymorphisms of Chromosome 9p21 and CD147 Were Relevant with the Carotid Plaque Risk in Acute Cerebral Infarction Patients Among Chinese Han Population. J. Mol. Neurosci. 2020, 70, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Mcintosh, A.M.; Simen, A.A.; Evans, K.L.; Hall, J.; MacIntyre, D.J.; Blackwood, D.; Morris, A.D.; Smith, B.H.; Dominiczak, A.; Porteous, D.; et al. Genetic variation in Hyperpolarization-activated cyclic nucleotide-gated channels and its relationship with neuroticism, cognition and risk of depression. Front. Genet. 2012, 3, 116. [Google Scholar] [CrossRef]
- Campbell, M.C.; Tishkoff, S.A. African genetic diversity: Implications for human demographic history, modern human origins, and complex disease mapping. Annu. Rev. Genomics. Hum. Genet. 2008, 9, 403–433. [Google Scholar] [CrossRef]
- Yu, N.; Chen, F.-C.; Ota, S.; Jorde, L.B.; Pamilo, P.; Patthy, L.; Ramsay, M.; Jenkins, T.; Shyue, S.-K.; Li, W.-H. Larger Genetic Differences Within Africans Than Between Africans and Eurasians. Genetics 2002, 161, 269–274. [Google Scholar] [CrossRef]
- Tucci, S.; Akey, J.M. The long walk to African genomics. Genome Biol. 2019, 20, 130. [Google Scholar] [CrossRef]
- Choudhury, A.; Aron, S.; Botigué, L.R.; Sengupta, D.; Botha, G.; Bensellak, T.; Wells, G.; Kumuthini, J.; Shriner, D.; Fakim, Y.J.; et al. High-depth African genomes inform human migration and health. Nature 2020, 586, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E. Global estimates of excess deaths from COVID-19. Nature 2022, 613, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Msemburi, W.; Karlinsky, A.; Knutson, V.; Aleshin-Guendel, S.; Chatterji, S.; Wakefield, J. The WHO estimates of excess mortality associated with the COVID-19 pandemic. Nature 2022, 613, 130–137. [Google Scholar] [CrossRef]
- Wang, H.; Paulson, K.R.; Pease, S.A.; Watson, S.; Comfort, H.; Zheng, P.; Aravkin, A.Y.; Bisignano, C.; Barber, R.M.; Alam, T. Estimating excess mortality due to the COVID-19 pandemic: A systematic analysis of COVID-19-related mortality, 2020–2021. Lancet 2022, 399, 1513–1536. [Google Scholar] [CrossRef]
- Wachira, L.J.; Arena, R.; Sallis, J.F.; Lambert, E.V.; Ong’Wen, O.M.; Laddu, D.R.; Onywera, V.; Oyeyemi, A.L. Why are COVID-19 effects less severe in Sub-Saharan Africa? Moving more and sitting less may be a primary reason. Prog. Cardiovasc. Dis. 2022, 71, 103–105. [Google Scholar] [CrossRef]
- Wamai, R.G.; Hirsch, J.L.; van Damme, W.; Alnwick, D.; Bailey, R.C.; Hodgins, S.; Alam, U.; Anyona, M. What Could Explain the Lower COVID-19 Burden in Africa despite Considerable Circulation of the SARS-CoV-2 Virus? Int. J. Environ. Res. Public Health 2021, 18, 8638. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic support for the out-of-Africa theory of human evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 6720–6722. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.; Hsieh, F.; Baronas, E.; Godbout, K.; Gosselin, M.; Stagliano, N.; Donovan, M.; Woolf, B.; Robison, K.; Jeyaseelan, R. A novel angiotensin-converting enzyme–related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9. Circ. Res. 2000, 87, e1–e9. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Rauf, A.; Khan, H.; Abu-Izneid, T. Renin-angiotensin-aldosterone (RAAS): The ubiquitous system for homeostasis and pathologies. Biomed. Pharmacother. 2017, 94, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Santos RA, S.; Sampaio, W.O.; Alzamora, A.C.; Motta-Santos, D.; Alenina, N.; Bader, M.; Campagnole-Santos, M.J. The ACE2/angiotensin-(1–7)/MAS axis of the renin-angiotensin system: Focus on angiotensin-(1–7). Physiol. Rev. 2017, 98, 505–553. [Google Scholar] [CrossRef] [PubMed]
- Hamming, I.; Timens, W.; Bulthuis, M.; Lely, A.; Navis, G.V.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef]
- Gu, J.; Korteweg, C. Pathology and pathogenesis of severe acute respiratory syndrome. Am. J. Pathol. 2007, 170, 1136–1147. [Google Scholar] [CrossRef]
- Gu, J.; Gong, E.; Zhang, B.; Zheng, J.; Gao, Z.; Zhong, Y.; Zou, W.; Zhan, J.; Wang, S.; Xie, Z. Multiple organ infection and the pathogenesis of SARS. J. Exp. Med. 2005, 202, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Beacon, T.H.; Delcuve, G.P.; Davie, J.R. Epigenetic regulation of ACE2, the receptor of the SARS-CoV-2 virus1. Genome 2021, 64, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Velkoska, E.; Freeman, M.; Wai, B.; Lancefield, T.F.; Burrell, L.M. From gene to protein—Experimental and clinical studies of ACE2 in blood pressure control and arterial hypertension. Front. Physiol. 2014, 5, 227. [Google Scholar] [CrossRef] [PubMed]
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M.G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: The major cell entry receptor for SARS-CoV-2. Lung 2020, 198, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Samavati, L.; Uhal, B.D. ACE2, much more than just a receptor for SARS-CoV-2. Front. Cell. Infect. Microbiol. 2020, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.J.; Tipnis, S.R.; Guy, J.L.; Rice, G.I.; Hooper, N.M. ACEH/ACE2 is a novel mammalian metallocarboxypeptidase and a homologue of angiotensin-converting enzyme insensitive to ACE inhibitors. Can. J. Physiol. Pharmacol. 2002, 80, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.-K.; Huang, I.-C.; Xu, K.; Vasilieva, N. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J. 2005, 24, 1634–1643. [Google Scholar] [CrossRef]
- Dhangadamajhi, G.; Mohapatra, B.N.; Kar, S.K.; Ranjit, M. Gene polymorphisms in angiotensin I converting enzyme (ACE I/D) and angiotensin II converting enzyme (ACE2 C→T) protect against cerebral malaria in Indian adults. Infect. Genet. Evol. 2010, 10, 337–341. [Google Scholar] [CrossRef]
- Rigat, B.; Hubert, C.; Alhenc-Gelas, F.; Cambien, F.; Corvol, P.; Soubrier, F. An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. J. Clin. Investig. 1990, 86, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Delgado, J.; Walther, T.; Rodriguez, A. The high blood pressure-malaria protection hypothesis. Circ. Res. 2016, 119, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Sampson, U.K.; Edwards, T.L.; Jahangir, E.; Munro, H.; Wariboko, M.; Wassef, M.G.; Fazio, S.; Mensah, G.A.; Kabagambe, E.K.; Blot, W.J. Factors associated with the prevalence of hypertension in the southeastern United States: Insights from 69 211 blacks and whites in the southern community cohort study. Circ. Cardiovasc. Qual. Outcomes 2014, 7, 33–54. [Google Scholar] [CrossRef] [PubMed]
- Riordan, J.F. Angiotensin-I-converting enzyme and its relatives. Genome Biol. 2003, 4, 225. [Google Scholar] [CrossRef] [PubMed]
- Rusmini, M.; Uva, P.; Amoroso, A.; Tolomeo, M.; Cavalli, A. How Genetics Might Explain the Unusual Link Between Malaria and COVID-19. Front. Med. 2021, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Pouladi, N.; Abdolahi, S. Investigating the ACE2 polymorphisms in COVID-19 susceptibility: An in silico analysis. Mol. Genet. Genom. Med. 2021, 9, e1672. [Google Scholar] [CrossRef] [PubMed]
- De, A.; Tiwari, A.; Dash, M.; Sinha, A. ACE2 mutation might explain lower COVID-19 burden in malaria endemic areas. Hum. Cell 2021, 34, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.I.H.; Albashir, A.A.D.; Elawad, O.A.M.A.; Homeida, A. Malaria and COVID-19: Unmasking their ties. Malar. J. 2020, 19, 457. [Google Scholar] [CrossRef]
- Badraoui, R.; Adnan, M.; Bardakci, F.; Alreshidi, M.M. Chloroquine and hydroxychloroquine interact differently with ACE2 domains reported to bind with the coronavirus spike protein: Mediation by ACE2 polymorphism. Molecules 2021, 26, 673. [Google Scholar] [CrossRef]
- Acquah, S. Implications of COVID-19 Pandemic on Evolution of Diabetes in Malaria-Endemic African Region. J. Diabetes Res. 2020, 2020, 8205261. [Google Scholar] [CrossRef] [PubMed]
- Möhlendick, B.; Schönfelder, K.; Breuckmann, K.; Elsner, C.; Babel, N.; Balfanz, P.; Dahl, E.; Dreher, M.; Fistera, D.; Herbstreit, F. ACE2 polymorphism and susceptibility for SARS-CoV-2 infection and severity of COVID-19. Pharm. Genom. 2021, 31, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Li, J.Y.; Wang, C.; Zhang, L.M.; Qiao, H. The ACE 2 G8790A polymorphism: Involvement in type 2 diabetes mellitus combined with cerebral stroke. J. Clin. Lab. Anal. 2017, 31, e22033. [Google Scholar] [CrossRef] [PubMed]
- Sabater Molina, M.; Nicolás Rocamora, E.; Bendicho, A.I.; Vázquez, E.G.; Zorio, E.; Rodriguez, F.D.; Gil Ortuño, C.; Rodríguez, A.I.; Sánchez-López, A.J.; Jara Rubio, R.; et al. Polymorphisms in ACE, ACE2, AGTR1 genes and severity of COVID-19 disease. PLoS ONE 2022, 17, e0263140. [Google Scholar] [CrossRef]
- Alimoradi, N.; Sharqi, M.; Firouzabadi, D.; Sadeghi, M.M.; Moezzi, M.I.; Firouzabadi, N. SNPs of ACE1 (rs4343) and ACE2 (rs2285666) genes are linked to SARS-CoV-2 infection but not with the severity of disease. Virol. J. 2022, 19, 48. [Google Scholar] [CrossRef] [PubMed]
- Karakaş Çelik, S.; Çakmak Genç, G.; Pişkin, N.; Açikgöz, B.; Altinsoy, B.; Kurucu Işsiz, B.; Dursun, A. Polymorphisms of ACE (I/D) and ACE2 receptor gene (Rs2106809, Rs2285666) are not related to the clinical course of COVID-19: A case study. J. Med. Virol. 2021, 93, 5947–5952. [Google Scholar] [CrossRef]
- Abdelsattar, S.; Kasemy, Z.A.; Ewida, S.F.; Abo-Elsoud, R.A.A.; Zytoon, A.A.; Abdelaal, G.A.; Abdelgawad, A.S.; Khalil, F.O.; Kamel, H. ACE2 and TMPRSS2 SNPs as Determinants of Susceptibility to, and Severity of, a COVID-19 Infection. Br. J. Biomed. Sci. 2022, 79, 10238. [Google Scholar] [CrossRef]
- Patel, S.K.; Wai, B.; Ord, M.; MacIsaac, R.J.; Grant, S.; Velkoska, E.; Panagiotopoulos, S.; Jerums, G.; Srivastava, P.; Burrell, L.M. Association of ACE2 genetic variants with blood pressure, left ventricular mass, and cardiac function in Caucasians with type 2 diabetes. Am. J. Hypertens. 2012, 25, 216–222. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, T.; Li, Y.; Guan, T.; Lai, Y.; Shen, Y.; Zeyaweiding, A.; Maimaiti, T.; Li, F.; Zhao, H.; et al. Association of ACE2 polymorphisms with susceptibility to essential hypertension and dyslipidemia in Xinjiang, China. Lipids Health Dis. 2018, 17, 241. [Google Scholar] [CrossRef]
- Hamet, P.; Pausova, Z.; Attaoua, R.; Hishmih, C.; Haloui, M.; Shin, J.; Paus, T.; Abrahamowicz, M.; Gaudet, D.; Santucci, L.; et al. SARS-CoV-2 Receptor ACE2 Gene Is Associated with Hypertension and Severity of COVID 19: Interaction with Sex, Obesity, and Smoking. Am. J. Hypertens. 2021, 34, 367–376. [Google Scholar] [CrossRef]
- Faridzadeh, A.; Mahmoudi, M.; Ghaffarpour, S.; Zamani, M.S.; Hoseinzadeh, A.; Naghizadeh, M.M.; Ghazanfari, T. The role of ACE1 I/D and ACE2 polymorphism in the outcome of Iranian COVID-19 patients: A case-control study. Front. Genet. 2022, 13, 955965. [Google Scholar] [CrossRef]
- Saniasiaya, J.; Kulasegarah, J. Dizziness and COVID-19. Ear Nose Throat J. 2021, 100, 29–30. [Google Scholar] [CrossRef] [PubMed]
- Korres, G.; Kitsos, D.K.; Kaski, D.; Tsogka, A.; Giannopoulos, S.; Giannopapas, V.; Sideris, G.; Tyrellis, G.; Voumvourakis, K. The Prevalence of Dizziness and Vertigo in COVID-19 Patients: A Systematic Review. Brain Sci. 2022, 12, 948. [Google Scholar] [CrossRef] [PubMed]
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Hearing loss, tinnitus, and dizziness in COVID-19: A systematic review and meta-analysis. Can. J. Neurol. Sci. 2022, 49, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Quan, B.; Guo, Y.; Zhang, J.; Lu, Y.; Feng, G.; Wu, Q.; Fang, F.; Cheng, L.; Jiao, N.; et al. The course of clinical diagnosis and treatment of a case infected with coronavirus disease 2019. J. Med. Virol. 2020, 92, 461–463. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, J.; Xia, L.; Cheng, X.; Lu, D. 18F-FDG PET/CT and serial chest CT findings in a COVID-19 patient with dynamic clinical characteristics in different period. Clin. Nucl. Med. 2020, 45, 495–496. [Google Scholar] [CrossRef]
- Chen, J.; Peng, S.; Zhang, B.; Liu, Z.; Liu, L.; Zhang, W. An uncommon manifestation of COVID-19 pneumonia on CT scan with small cavities in the lungs: A case report. Medicine 2020, 99, e21240. [Google Scholar] [CrossRef] [PubMed]
- Wander, P.L.; Lowy, E.; Beste, L.A.; Tulloch-Palomino, L.; Korpak, A.; Peterson, A.C.; Kahn, S.E.; Boyko, E.J. The incidence of diabetes among 2,777,768 veterans with and without recent SARS-CoV-2 infection. Diabetes Care 2022, 45, 782–788. [Google Scholar] [CrossRef]
- Jalaleddine, N.; Bouzid, A.; Hachim, M.; Sharif-Askari, N.S.; Mahboub, B.; Senok, A.; Halwani, R.; Hamoudi, R.A.; Al Heialy, S. ACE2 polymorphisms impact COVID-19 severity in obese patients. Sci. Rep. 2022, 12, 21491. [Google Scholar] [CrossRef]
- Elbadri, S.A.; Abdallah, N.M.A.; El-Shokry, M.; Gaber, A.; Elsayed, M.K. Association between single nucleotide polymorphism of human angiotensin-converting enzyme 2 gene locus and clinical severity of COVID-19. Egypt. J. Med. Hum. Genet. 2022, 23, 1–8. [Google Scholar] [CrossRef]
- He, Y.; Yang, W.; Liu, S.; Gan, L.; Zhang, F.; Mu, C.; Wang, J.; Qu, L.; Wang, R.; Deng, J.; et al. Interactions between angiotensin-converting enzyme-2 polymorphisms and high salt intake increase the risk of hypertension in the Chinese Wa population. Int. J. Clin. Exp. Pathol. 2017, 10, 11159–11168. [Google Scholar] [PubMed]
- Darbani, B. The expression and polymorphism of entry machinery for COVID-19 in human: Juxtaposing population groups, gender, and different tissues. Int. J. Environ. Res. Public Health 2020, 17, 3433. [Google Scholar] [CrossRef]
- Khayat, A.S.; de Assumpção, P.P.; Meireles Khayat, B.C.; Thomaz Araújo, T.M.; Batista-Gomes, J.A.; Imbiriba, L.C.; Ishak, G.; de Assumpção, P.B.; Moreira, F.C.; Burbano, R.R. ACE2 polymorphisms as potential players in COVID-19 outcome. PLoS ONE 2020, 15, e0243887. [Google Scholar] [CrossRef]
- Gemmati, D.; Tisato, V. Genetic Hypothesis and Pharmacogenetics Side of Renin-Angiotensin-System in COVID-19. Genes 2020, 11, 1044. [Google Scholar] [CrossRef] [PubMed]
- Paniri, A.; Hosseini, M.M.; Moballegh-Eslam, M.; Akhavan-Niaki, H. Comprehensive in silico identification of impacts of ACE2 SNPs on COVID-19 susceptibility in different populations. Gene Rep. 2021, 22, 100979. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, L.; Feng, Z.; Wan, S.; Huang, P.; Sun, X.; Wen, F.; Huang, X.; Ning, G.; Wang, W. Comparative genetic analysis of the novel coronavirus (2019-nCoV/SARS-CoV-2) receptor ACE2 in different populations. Cell Discov. 2020, 6, 11. [Google Scholar] [CrossRef]
- Guo, X.; Chen, Z.; Xia, Y.; Lin, W.; Li, H. Investigation of the genetic variation in ACE2 on the structural recognition by the novel coronavirus (SARS-CoV-2). J. Transl. Med. 2020, 18, 321. [Google Scholar] [CrossRef]
- Abe, M.; Tahara, M.; Sakai, K.; Yamaguchi, H.; Kanou, K.; Shirato, K.; Kawase, M.; Noda, M.; Kimura, H.; Matsuyama, S. TMPRSS2 is an activating protease for respiratory parainfluenza viruses. J. Virol. 2013, 87, 11930–11935. [Google Scholar] [CrossRef]
- Glowacka, I.; Bertram, S.; Müller, M.A.; Allen, P.; Soilleux, E.; Pfefferle, S.; Steffen, I.; Tsegaye, T.S.; He, Y.; Gnirss, K. Evidence that TMPRSS2 activates the severe acute respiratory syndrome coronavirus spike protein for membrane fusion and reduces viral control by the humoral immune response. J. Virol. 2011, 85, 4122–4134. [Google Scholar] [CrossRef]
- Bertram, S.; Dijkman, R.; Habjan, M.; Heurich, A.; Gierer, S.; Glowacka, I.; Welsch, K.; Winkler, M.; Schneider, H.; Hofmann-Winkler, H. TMPRSS2 activates the human coronavirus 229E for cathepsin-independent host cell entry and is expressed in viral target cells in the respiratory epithelium. J. Virol. 2013, 87, 6150–6160. [Google Scholar] [CrossRef]
- Shirato, K.; Kawase, M.; Matsuyama, S. Middle East respiratory syndrome coronavirus infection mediated by the transmembrane serine protease TMPRSS2. J. Virol. 2013, 87, 12552–12561. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.; Uckeley, Z.M.; Doldan, P.; Stanifer, M.; Boulant, S.; Lozach, P.Y. TMPRSS2 expression dictates the entry route used by SARS-CoV-2 to infect host cells. EMBO J. 2021, 40, e107821. [Google Scholar] [CrossRef] [PubMed]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; Rohde, C. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef]
- Wilson, S.K.; Greer, B.; Hooper, J.; Zijlstra, A.; Walker, B.; Quigley, J.P.; Hawthorne, S. The membrane-anchored serine protease, TMPRSS2, activates PAR-2 in prostate cancer cells. Biochem. J. 2005, 388, 967–972. [Google Scholar] [CrossRef]
- Ko, C.-J.; Huang, C.-C.; Lin, H.-Y.; Juan, C.-P.; Lan, S.-W.; Shyu, H.-Y.; Wu, S.-R.; Hsiao, P.-W.; Huang, H.-P.; Shun, C.-T.; et al. Androgen-induced TMPRSS2 activates matriptase and promotes extracellular matrix degradation, prostate cancer cell invasion, tumor growth, and metastasis. Cancer Res. 2015, 75, 2949–2960. [Google Scholar] [CrossRef] [PubMed]
- Lubieniecka, J.M.; Cheteri, M.K.; Stanford, J.L.; Ostrander, E.A. Met160Val polymorphism in the TRMPSS2 gene and risk of prostate cancer in a population-based case-control study. Prostate 2004, 59, 357–359. [Google Scholar] [CrossRef]
- Wang, A.; Chiou, J.; Poirion, O.B.; Buchanan, J.; Valdez, M.J.; Verheyden, J.M.; Hou, X.; Kudtarkar, P.; Narendra, S.; Newsome, J.M.; et al. Single-cell multiomic profiling of human lungs reveals cell-type-specific and age-dynamic control of SARS-CoV2 host genes. eLife 2020, 9, e62522. [Google Scholar] [CrossRef]
- Asselta, R.; Paraboschi, E.M.; Mantovani, A.; Duga, S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Aging 2020, 12, 10087. [Google Scholar] [CrossRef]
- Kehdy, F.S.; Pita-Oliveira, M.; Scudeler, M.M.; Torres-Loureiro, S.; Zolini, C.; Moreira, R.; Michelin, L.A.; Alvim, I.; Silva-Carvalho, C.; Furlan, V.C. Human-SARS-CoV-2 interactome and human genetic diversity: TMPRSS2-rs2070788, associated with severe influenza, and its population genetics caveats in Native Americans. Genet. Mol. Biol. 2021, 44, e20200484. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Choudhari, R.; Nema, V.; Khan, A.A. ACE2 and TMPRSS2 polymorphisms in various diseases with special reference to its impact on COVID-19 disease. Microb. Pathog. 2021, 150, 104621. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.K.; Srivastava, A.; Singh, P.P.; Chaubey, G. Genetic association of TMPRSS2 rs2070788 polymorphism with COVID-19 case fatality rate among Indian populations. Infect. Genet. Evol. 2022, 98, 105206. [Google Scholar] [CrossRef]
- Schönfelder, K.; Breuckmann, K.; Elsner, C.; Dittmer, U.; Fistera, D.; Herbstreit, F.; Risse, J.; Schmidt, K.; Sutharsan, S.; Taube, C. Transmembrane serine protease 2 polymorphisms and susceptibility to severe acute respiratory syndrome coronavirus type 2 infection: A German case-control study. Front. Genet. 2021, 12, 667231. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, M.; Hay Mele, B.; Benetti, E.; Fallerini, C.; Baldassarri, M.; Furini, S.; Frullanti, E.; Mari, F.; Study, G.-C.M.; Andreotti, G. Protective role of a TMPRSS2 variant on severe COVID-19 outcome in young males and elderly women. Genes 2021, 12, 596. [Google Scholar] [CrossRef]
- Wulandari, L.; Hamidah, B.; Pakpahan, C.; Damayanti, N.S.; Kurniati, N.D.; Adiatmaja, C.O.; Wigianita, M.R.; Soedarsono; Husada, D.; Tinduh, D.; et al. Initial study on TMPRSS2 p. Val160Met genetic variant in COVID-19 patients. Hum. Genom. 2021, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Ravikanth, V.; Sasikala, M.; Naveen, V.; Latha, S.S.; Parsa, K.V.L.; Vijayasarathy, K.; Amanchy, R.; Avanthi, S.; Govardhan, B.; Rakesh, K. A variant in TMPRSS2 is associated with decreased disease severity in COVID-19. Meta Gene 2021, 29, 100930. [Google Scholar] [CrossRef]
- David, A.; Parkinson, N.; Peacock, T.P.; Pairo-Castineira, E.; Khanna, T.; Cobat, A.; Tenesa, A.; Sancho-Shimizu, V.; Casanova, J.-L.; Abel, L. A common TMPRSS2 variant has a protective effect against severe COVID-19. Curr. Res. Transl. Med. 2022, 70, 103333. [Google Scholar] [CrossRef]
- Torre-Fuentes, L.; Matías-Guiu, J.; Hernández-Lorenzo, L.; Montero-Escribano, P.; Pytel, V.; Porta-Etessam, J.; Gómez-Pinedo, U.; Matías-Guiu, J.A. ACE2, TMPRSS2, and Furin variants and SARS-CoV-2 infection in Madrid, Spain. J. Med. Virol. 2021, 93, 863–869. [Google Scholar] [CrossRef]
- Rokni, M.; Heidari Nia, M.; Sarhadi, M.; Mirinejad, S.; Sargazi, S.; Moudi, M.; Saravani, R.; Rahdar, S.; Kargar, M. Association of TMPRSS2 Gene Polymorphisms with COVID-19 Severity and Mortality: A Case-Control Study with Computational Analyses. Appl. Biochem. Biotechnol. 2022, 194, 3507–3526. [Google Scholar] [CrossRef] [PubMed]
- Villapalos-García, G.; Zubiaur, P.; Rivas-Durán, R.; Campos-Norte, P.; Arévalo-Román, C.; Fernández-Rico, M.; Fraile, L.G.-F.; Fernández-Campos, P.; Soria-Chacartegui, P.; de Córdoba-Oñate, S.F. Transmembrane protease serine 2 (TMPRSS2) rs75603675, comorbidity, and sex are the primary predictors of COVID-19 severity. Life Sci. Alliance 2022, 5, e202201396. [Google Scholar] [CrossRef]
- Soker, S.; Takashima, S.; Miao, H.Q.; Neufeld, G.; Klagsbrun, M. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 1998, 92, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Staton, C.; Kumar, I.; Reed, M.; Brown, N. Neuropilins in physiological and pathological angiogenesis. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2007, 212, 237–248. [Google Scholar] [CrossRef]
- He, Z.; Tessier-Lavigne, M. Neuropilin is a receptor for the axonal chemorepellent Semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef]
- Teesalu, T.; Sugahara, K.N.; Kotamraju, V.R.; Ruoslahti, E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proc. Natl. Acad. Sci. USA 2009, 106, 16157–16162. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.-E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van Der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A. Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Chapoval, S.P.; Keegan, A.D. Perspectives and potential approaches for targeting neuropilin 1 in SARS-CoV-2 infection. Mol. Med. 2021, 27, 162. [Google Scholar] [CrossRef]
- Hashemi, S.M.A.; Thijssen, M.; Hosseini, S.Y.; Tabarraei, A.; Pourkarim, M.R.; Sarvari, J. Human gene polymorphisms and their possible impact on the clinical outcome of SARS-CoV-2 infection. Arch. Virol. 2021, 166, 2089–2108. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, W. Neurological manifestations in coronavirus disease 2019 (COVID-19) patients: A systematic review of literature. CNS Spectr. 2020, 27, 145–156. [Google Scholar] [CrossRef]
- Pollock, C.E.; Sutherland, H.G.; Maher, B.H.; Lea, R.A.; Haupt, L.M.; Frith, A.; Macgregor, E.A.; Griffiths, L.R. The NRP1 migraine risk variant shows evidence of association with menstrual migraine. J. Headache Pain 2018, 19, 31. [Google Scholar] [CrossRef]
- Gormley, P.; Anttila, V.; Winsvold, B.S.; Palta, P.; Esko, T.; Pers, T.H.; Farh, K.-H.; Cuenca-Leon, E.; Muona, M.; Furlotte, N.A. Meta-analysis of 375,000 individuals identifies 38 susceptibility loci for migraine. Nat. Genet. 2016, 48, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Chen, L.; Zhong, W.D.; Zhang, Z.; Mi, L.; Zhang, Y.; Liao, C.G.; Bian, H.J.; Jiang, J.L. HAb18G (CD147), a cancer-associated biomarker and its role in cancer detection. Histopathology 2009, 54, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Pushkarsky, T.; Zybarth, G.; Dubrovsky, L.; Yurchenko, V.; Tang, H.; Guo, H.; Toole, B.; Sherry, B.; Bukrinsky, M. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A. Proc. Natl. Acad. Sci. USA 2001, 98, 6360–6365. [Google Scholar] [CrossRef]
- Zhang, M.-Y.; Zhang, Y.; Wu, X.-D.; Zhang, K.; Lin, P.; Bian, H.-J.; Qin, M.-M.; Huang, W.; Wei, D.; Zhang, Z. Disrupting CD147-RAP2 interaction abrogates erythrocyte invasion by Plasmodium falciparum. Blood J. Am. Soc. Hematol. 2018, 131, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhang, W.; Wang, S.J.; Yu, X.L.; Tang, J.; Huang, W.; Li, Y.; Cui, H.Y.; Guo, Y.S.; Tavernier, J. HAb18G/CD147 promotes cell motility by regulating annexin II-activated RhoA and Rac1 signaling pathways in hepatocellular carcinoma cells. Hepatology 2011, 54, 2012–2024. [Google Scholar] [CrossRef]
- Lu, M.; Wu, J.; Hao, Z.; Shang, Y.; Xu, J.; Nan, G.; Li, X.; Chen, Z.; Bian, H. Basolateral CD147 induces hepatocyte polarity loss by E-cadherin ubiquitination and degradation in hepatocellular carcinoma progress. Hepatology 2018, 68, 317–332. [Google Scholar] [CrossRef]
- Chen, Z.; Mi, L.; Xu, J.; Yu, J.; Wang, X.; Jiang, J.; Xing, J.; Shang, P.; Qian, A.; Li, Y. Function of HAb18G/CD147 in invasion of host cells by severe acute respiratory syndrome coronavirus. J. Infect. Dis. 2005, 191, 755–760. [Google Scholar] [CrossRef]
- Shilts, J.; Crozier, T.W.; Greenwood, E.J.; Lehner, P.J.; Wright, G.J. No evidence for basigin/CD147 as a direct SARS-CoV-2 spike binding receptor. Sci. Rep. 2021, 11, 413. [Google Scholar] [CrossRef]
- Fenizia, C.; Galbiati, S.; Vanetti, C.; Vago, R.; Clerici, M.; Tacchetti, C.; Daniele, T. SARS-CoV-2 Entry: At the Crossroads of CD147 and ACE2. Cells 2021, 10, 1434. [Google Scholar] [CrossRef]
- Geng, J.; Chen, L.; Yuan, Y.; Wang, K.; Wang, Y.; Qin, C.; Wu, G.; Chen, R.; Zhang, Z.; Wei, D. CD147 antibody specifically and effectively inhibits infection and cytokine storm of SARS-CoV-2 and its variants delta, alpha, beta, and gamma. Signal Transduct. Target. Ther. 2021, 6, 347. [Google Scholar] [CrossRef]
- Mao, Y.; Yan, J.; Wang, C.; Wang, Z.; Liu, P.; Yuan, W. CD147 expression level and rs8259 T/A polymorphism of CD147 in patients with acute coronary syndrome. Zhonghua Xin Xue Guan Bing Za Zhi 2014, 42, 566–570. [Google Scholar]
- Li, M.-P.; Hu, X.-L.; Yang, Y.-L.; Zhang, Y.-J.; Zhou, J.-P.; Peng, L.-M.; Tang, J.; Chen, X.-P. Basigin rs8259 polymorphism confers decreased risk of chronic heart failure in a Chinese population. Int. J. Environ. Res. Public Health 2017, 14, 211. [Google Scholar] [CrossRef]
- Latini, A.; Agolini, E.; Novelli, A.; Borgiani, P.; Giannini, R.; Gravina, P.; Smarrazzo, A.; Dauri, M.; Andreoni, M.; Rogliani, P. COVID-19 and genetic variants of protein involved in the SARS-CoV-2 entry into the host cells. Genes 2020, 11, 1010. [Google Scholar] [CrossRef]
- Crosnier, C.; Bustamante, L.Y.; Bartholdson, S.J.; Bei, A.K.; Theron, M.; Uchikawa, M.; Mboup, S.; Ndir, O.; Kwiatkowski, D.P.; Duraisingh, M.T. Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum. Nature 2011, 480, 534–537. [Google Scholar] [CrossRef]
- Xiong, L.; Edwards, C.K., III; Zhou, L. The biological function and clinical utilization of CD147 in human diseases: A review of the current scientific literature. Int. J. Mol. Sci. 2014, 15, 17411–17441. [Google Scholar] [CrossRef]
- Gudowska-Sawczuk, M.; Mroczko, B. The Role of Neuropilin-1 (NRP-1) in SARS-CoV-2 Infection: Review. J. Clin. Med. 2021, 10, 2772. [Google Scholar] [CrossRef] [PubMed]
- Petersen, D.; Steyl, C.; Scholtz, D.; Baker, B.; Abdullah, I.; Uren, C.; Möller, M. African Genetic Representation in the Context of SARS-CoV-2 Infection and COVID-19 Severity. Front. Genet. 2022, 13, 909117. [Google Scholar] [CrossRef] [PubMed]
- Mostafa-Hedeab, G. ACE2 as drug target of COVID-19 virus treatment, simplified updated review. Rep. Biochem. Mol. Biol. 2020, 9, 97. [Google Scholar] [CrossRef]
- Vitiello, A.; Ferrara, F.; Porta, R. Remdesivir and COVID-19 infection, therapeutic benefits or unnecessary risks? Ir. J. Med. Sci. 2021, 190, 1637–1638. [Google Scholar] [CrossRef] [PubMed]
- Charoute, H.; Elkarhat, Z.; Elkhattabi, L.; El Fahime, E.; Oukkache, N.; Rouba, H.; Barakat, A. Computational screening of potential drugs against COVID-19 disease: The Neuropilin-1 receptor as molecular target. VirusDisease 2022, 33, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kiso, M.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; Imai, M.; Takeda, M.; Kinoshita, N.; Ohmagari, N.; Gohda, J.; Semba, K. The anticoagulant nafamostat potently inhibits SARS-CoV-2 S protein-mediated fusion in a cell fusion assay system and viral infection in vitro in a cell-type-dependent manner. Viruses 2020, 12, 629. [Google Scholar] [CrossRef]
- Uddin, H.; Zonder, J.A.; Azmi, A.S. Exportin 1 inhibition as antiviral therapy. Drug Discov. Today 2020, 25, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Yagin, F.H.; Cicek, İ.B.; Alkhateeb, A.; Yagin, B.; Colak, C.; Azzeh, M.; Akbulut, S. Explainable artificial intelligence model for identifying COVID-19 gene biomarkers. Comput. Biol. Med. 2023, 154, 106619. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variant ID | Mutation | Disease(s) | Genetic Ancestry | Proposed Mechanism | Citation | MAF |
|---|---|---|---|---|---|---|
| ACE2 | ||||||
| rs1978124 | T>C | Cardiovascular disease | Uygurs—East Asian | Intron variant | [27] | AFR: 0.90 |
| EUR: 0.53 | ||||||
| SAS: 0.78 | ||||||
| rs233575 | G>A | Cardiovascular disease, Hypertension | East Asian and European | Intron variant | [27,28] | AFR: 0.99 |
| EUR: 0.65 | ||||||
| SAS: 0.82 | ||||||
| rs1514282 | T>C | Blood pressure | East Asian | Intron variant | [29] | AFR: 0.32 |
| EUR: 0.00 | ||||||
| SAS: 0.09 | ||||||
| rs16997078 | A>G | No disease associations found | Intron Variant | AFR: 0.24 | ||
| EUR: 0.00 | ||||||
| SAS: 0.09 | ||||||
| rs2097723 | T>C | Blood pressure | East Asian | Intron Variant | [30] | AFR: 0.07 |
| EUR: 0.28 | ||||||
| SAS: 0.22 | ||||||
| rs879922 | C>G | Hypertension, Cardiovascular disease, blood pressure and T2D. | East Asian | Intron Variant | [27,31] | AFR: 0.45 |
| EUR: 0.65 | ||||||
| SAS: 0.71 | ||||||
| rs2106809 | A>G | Blood pressure | European | Intron Variant | [32] | AFR: 0.09 |
| EUR: 0.25 | ||||||
| SAS: 0.48 | ||||||
| rs2048683 | T>G | Cardiovascular disease | East Asian | Intron Variant | [27] | AFR: 0.81 |
| EUR: 0.65 | ||||||
| SAS: 0.80 | ||||||
| rs4646120 | G>A | Shared haplotype between South Asian and East Eurasians for host susceptibility to SARS-CoV-2. | South Asian | Intron Variant | [33] | AFR: 0.68 |
| EUR: 0.53 | ||||||
| SAS: 0.78 | ||||||
| rs4646140 | C>T | Blood pressure | East Asian | Intron Variant | [31] | AFR: 0.13 |
| EUR: 0.00 | ||||||
| SAS: 0.09 | ||||||
| rs35697037 | G>A | Significant correlation with dizziness symptoms in mild traumatic brain injury. | East Asian | Intron Variant | [34] | AFR: 0.27 |
| EUR: 0.38 | ||||||
| SAS: 0.24 | ||||||
| rs714205 | C>G | Hypertension, and Retinopathy in T2D | East Asian | Intron Variant | [29,35] | AFR: 0.10 |
| EUR: 0.21 | ||||||
| SAS: 0.47 | ||||||
| TMPRSS2 | ||||||
| rs463727 | T>A | Susceptibility and severity of respiratory disease | East Asian And European | 500 KB downstream variant | [36] | AFR: 0.08 |
| EUR: 0.46 | ||||||
| SAS: 0.34 | ||||||
| rs35041537 | C>T | Infectivity and Progression of SARS-CoV-2 | South Asian | Intron Variant | [37] | AFR: 0.11 |
| EUR: 0.46 | ||||||
| SAS: 0.34 | ||||||
| rs7275220 | G>A | Breast Cancer | European | Intron Variant | [38] | AFR: 0.49 |
| EUR: 0.75 | ||||||
| SAS: 0.66 | ||||||
| rs456298 | T>A | Associated with high risk of SARS-CoV-2 | Mexican -European | 3 Prime UTR Variant | [39] | AFR: 0.63 |
| EUR: 0.83 | ||||||
| SAS: 0.68 | ||||||
| rs2070788 | G>A | Susceptibility to infectious disease | East Asian | Intron Variant | [40] | AFR: 0.73 |
| EUR: 0.54 | ||||||
| SAS: 0.53 | ||||||
| rs8127290 | G>A | OAS variant associated with rubella | European | None | [41] | AFR: 0.35 |
| EUR: 0.16 | ||||||
| SAS: 0.22 | ||||||
| rs383510 | T>C | Susceptibility to infectious disease | East Asian | Intron Variant | [40] | AFR: 0.67 |
| EUR: 0.51 | ||||||
| SAS: 0.55 | ||||||
| rs11910678 | T>C | Susceptibility to SARS-CoV-2 | None | 3 Prime UTR Variant | [42] | AFR: 0.14 |
| EUR: 0.00 | ||||||
| SAS: 0.003 | ||||||
| rs1557372 | C>T | Alzheimer’s disease | East Asian | None | [43] | AFR: 0.50 |
| EUR: 0.37 | ||||||
| SAS: 0.32 | ||||||
| rs75603675 | C>A | Associated with SARS-CoV-2 disease susceptibility and severity | Eastern European | Missense Variant | [44] | AFR: 0.30 |
| EUR: 0.40 | ||||||
| SAS: 0.22 | ||||||
| NRP1 | ||||||
| rs2804495 | G>T | Neovascular age-related macular degeneration | European | Intron variant | [45] | AFR: 0.24 |
| EUR: 0.71 | ||||||
| SAS: 0.57 | ||||||
| rs927099 | T>C | Late-onset Alzheimer disease. | European | Intron Variant | [46] | AFR: 0.88 |
| EUR: 0.50 | ||||||
| SAS: 0.55 | ||||||
| rs1048804 | A>G | Type 1 diabetes | European | Synonymous Variant | [47] | AFR: 0.57 |
| EUR: 0.25 | ||||||
| SAS: 0.37 | ||||||
| rs10080 | G>A | Risk of tetralogy of Fallot | East Asian | Non-Coding Transcript Variant | [48] | AFR: 0.70 |
| EUR: 0.43 | ||||||
| SAS: 0.50 | ||||||
| rs1319013 | T>G | Late-onset Alzheimer disease | European | Intron variant | [49] | AFR: 0.37 |
| EUR: 0.53 | ||||||
| SAS: 0.52 | ||||||
| rs1571781 | A>G | Late-onset Alzheimer disease | European | Intron variant | [46] | AFR: 0.78 |
| EUR: 0.62 | ||||||
| SAS: 0.51 | ||||||
| rs2070296 | C>T | Associated with worse response to ranibizumab treatment in neovascular age-related macular degeneration | European | Synonymous Variant | [45] | AFR: 0.32 |
| EUR: 0.16 | ||||||
| SAS: 0.23 | ||||||
| rs2506144 | C>T | No disease associations | European | Non-Coding Transcript Variant | AFR: 0.33 | |
| EUR: 0.18 | ||||||
| SAS: 0.09 | ||||||
| rs1888686 | T>C | Late-onset Alzheimer disease | European | Intron Variant | [46] | AFR: 0.34 |
| EUR: 0.21 | ||||||
| SAS: 0.26 | ||||||
| rs12573218 | C>T | Osteonecrosis of the femoral head | East Asian | Intron Variant | [50] | AFR: 0.02 |
| EUR: 0.15 | ||||||
| SAS: 0.10 | ||||||
| rs1010826 | G>A | Type 1 diabetes | European | Intron Variant | [47] | AFR: 0.35 |
| EUR: 0.23 | ||||||
| SAS: 0.41 | ||||||
| rs1331326 | C>T | Late-onset Alzheimer disease | European | Intron Variant | [46] | AFR: 0.53 |
| EUR: 0.65 | ||||||
| SAS: 0.65 | ||||||
| rs1888685 | C>T | Late-onset Alzheimer disease | European | Intron Variant | [46] | AFR: 0.01 |
| EUR: 0.13 | ||||||
| SAS: 0.23 | ||||||
| rs12358370 | C>G | Osteonecrosis of the femoral head | East Asian | Intron Variant | [50] | AFR: 0.03 |
| EUR: 0.15 | ||||||
| SAS: 0.12 | ||||||
| rs2228638 | C>T | Risk of tetralogy of Fallot | East Asian | Missense Variant | [48] | AFR: 0.01 |
| EUR: 0.11 | ||||||
| SAS: 0.13 | ||||||
| CD147 | ||||||
| rs8637 | A>G | Coronary Heart Disease | East Asian | 3 Prime UTR Variant | [51] | AFR: 0.97 |
| EUR: 0.47 | ||||||
| SAS: 0.83 | ||||||
| rs8259 | T>A | Coronary Heart Disease, A miRNA-492 binding-site polymorphism in BSG (basigin) confers risk to psoriasis in central south Chinese population. | East Asian | 3 Prime UTR Variant | [51,52] | AFR: 0.56 |
| EUR: 0.30 | ||||||
| SAS: 0.63 | ||||||
| rs4919862 | T>C | Carotid Plaque Risk in Acute Cerebral Infarction | East Asian | Intron Variant | [53] | AFR: 0.98 |
| EUR: 0.75 | ||||||
| SAS: 0.92 | ||||||
| rs4919859 | G>C | Coronary Heart Disease | East Asian | Intron Variant, 2 KB Upstream Variant | [51] | AFR: 0.51 |
| EUR: 0.35 | ||||||
| SAS: 0.50 | ||||||
| rs6758 | G>A | Coronary Heart Disease, Hyperpolarization-activated cyclic nucleotide-gated channels and its relationship with neuroticism, cognition, and risk of depression | East Asian and European | 3 Prime UTR Variant | [51,54] | AFR: 0.21 |
| EUR: 0.09 | ||||||
| SAS: 0.25 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adimulam, T.; Arumugam, T.; Gokul, A.; Ramsuran, V. Genetic Variants within SARS-CoV-2 Human Receptor Genes May Contribute to Variable Disease Outcomes in Different Ethnicities. Int. J. Mol. Sci. 2023, 24, 8711. https://doi.org/10.3390/ijms24108711
Adimulam T, Arumugam T, Gokul A, Ramsuran V. Genetic Variants within SARS-CoV-2 Human Receptor Genes May Contribute to Variable Disease Outcomes in Different Ethnicities. International Journal of Molecular Sciences. 2023; 24(10):8711. https://doi.org/10.3390/ijms24108711
Chicago/Turabian StyleAdimulam, Theolan, Thilona Arumugam, Anmol Gokul, and Veron Ramsuran. 2023. "Genetic Variants within SARS-CoV-2 Human Receptor Genes May Contribute to Variable Disease Outcomes in Different Ethnicities" International Journal of Molecular Sciences 24, no. 10: 8711. https://doi.org/10.3390/ijms24108711