
The Crucial Roles of Pitx3 in Midbrain Dopaminergic Neuron Development and Parkinson’s Disease-Associated Neurodegeneration

Abstract
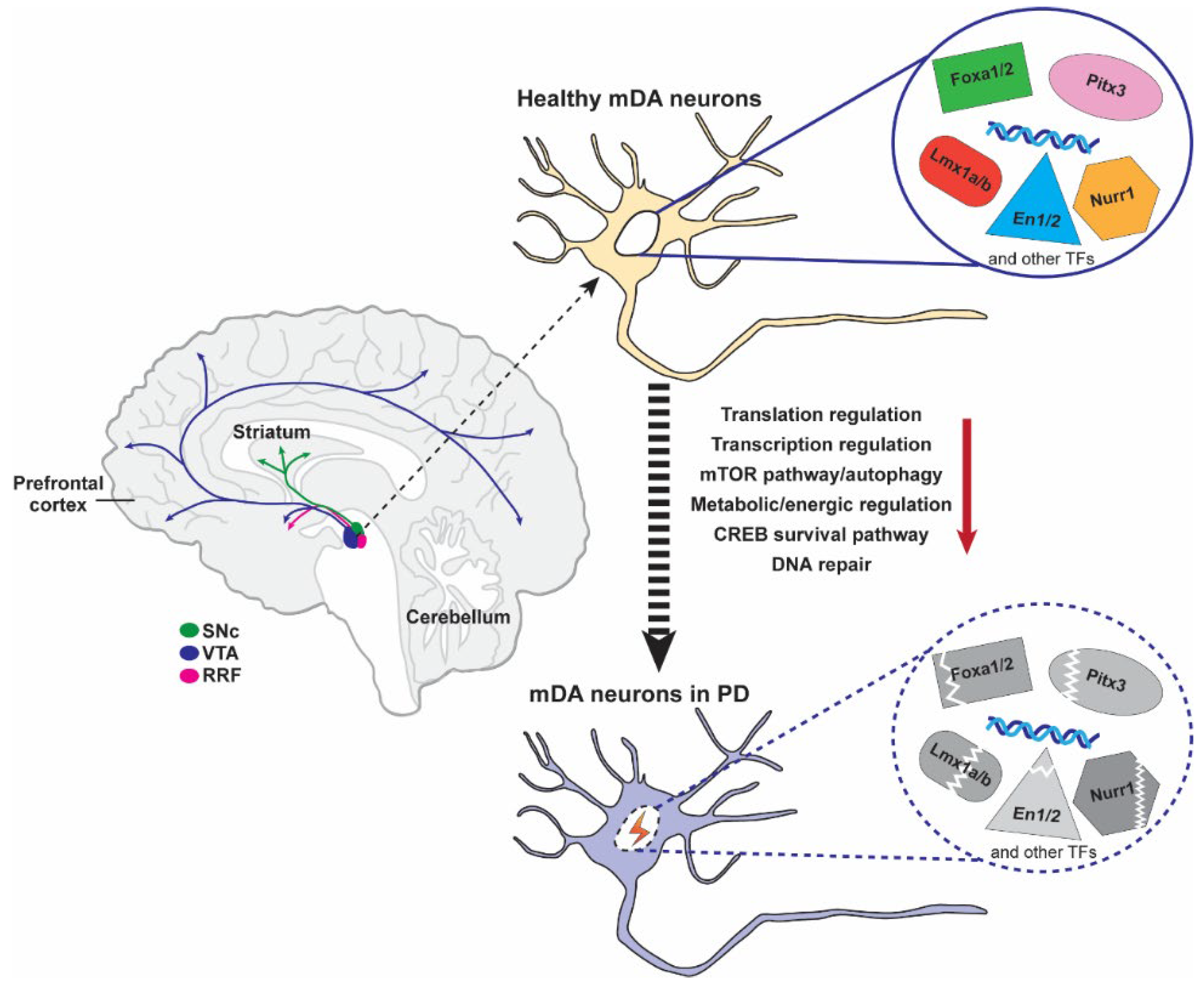
:1. Background
2. Introduction of Pitx3
2.1. Pitx3 Structure
2.2. Pitx3 Protein
2.3. Pitx3 in the Main Subgroups of mDA Neurons
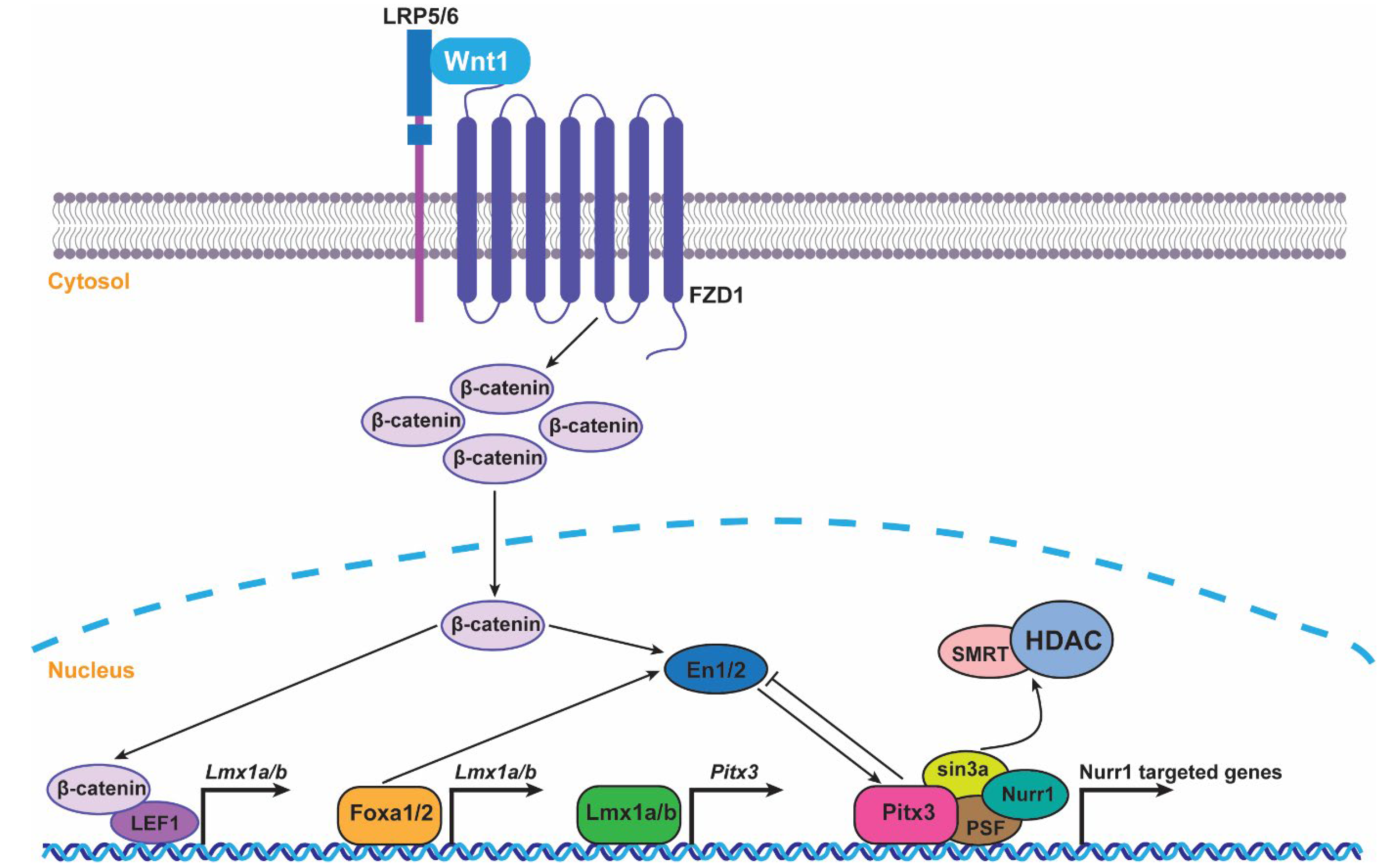
3. Pitx3 and Pitx3-Related Transcriptional Networks in mDA Neuron Development
3.1. Pitx3 and Lmx1 a/b
3.2. Pitx3 and Nurr1
3.3. Pitx3 and Engrailed-1/Engrailed-2
3.4. Pitx3 and Foxa1/2
4. The Broad Role of Pitx3 in PD: From Model to Patients
4.1. Development of Pitx3-Deficient Mouse Models
4.2. The Role of Pitx3 in Cell Therapy
4.3. Pitx3 and PD
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- GBD 2016 Neurology Collaborators. Global, regional, and national burden of neurological disorders, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Foffani, G.; Dehay, B.; Bezard, E.; Obeso, J.A. Motor and non-motor circuit disturbances in early Parkinson disease: Which happens first? Nat. Rev. Neurosci. 2022, 23, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Stecher, B.; Labrie, V.; Brundin, L.; Brundin, P. Triggers, Facilitators, and Aggravators: Redefining Parkinson’s Disease Pathogenesis. Trends Neurosci. 2019, 42, 4–13. [Google Scholar] [CrossRef]
- Panicker, N.; Ge, P.; Dawson, V.L.; Dawson, T.M. The cell biology of Parkinson’s disease. J. Cell Biol. 2021, 220, e202012095. [Google Scholar] [CrossRef]
- Stoker, T.B.; Greenland, J.C. (Eds.) Parkinson’s Disease: Pathogenesis and Clinical Aspects; Codon Publications: Brisbane, Australian, 2018. [Google Scholar]
- Smidt, M.P.; Burbach, J.P. How to make a mesodiencephalic dopaminergic neuron. Nat. Rev. Neurosci. 2007, 8, 21–32. [Google Scholar] [CrossRef]
- Burbach, J.P.; Smidt, M.P. Molecular programming of stem cells into mesodiencephalic dopaminergic neurons. Trends Neurosci. 2006, 29, 601–603. [Google Scholar] [CrossRef]
- Bayer, H.M.; Glimcher, P.W. Midbrain dopamine neurons encode a quantitative reward prediction error signal. Neuron 2005, 47, 129–141. [Google Scholar] [CrossRef]
- Smits, S.M.; Ponnio, T.; Conneely, O.M.; Burbach, J.P.; Smidt, M.P. Involvement of Nurr1 in specifying the neurotransmitter identity of ventral midbrain dopaminergic neurons. Eur. J. Neurosci. 2003, 18, 1731–1738. [Google Scholar] [CrossRef]
- Arber, S.; Costa, R.M. Networking brainstem and basal ganglia circuits for movement. Nat. Rev. Neurosci. 2022, 23, 342–360. [Google Scholar] [CrossRef]
- Cox, J.; Witten, I.B. Striatal circuits for reward learning and decision-making. Nat. Rev. Neurosci. 2019, 20, 482–494. [Google Scholar] [CrossRef]
- Kouwenhoven, W.M.; von Oerthel, L.; Smidt, M.P. Pitx3 and En1 determine the size and molecular programming of the dopaminergic neuronal pool. PLoS ONE 2017, 12, e0182421. [Google Scholar] [CrossRef]
- Gerlach, M.; Reichmann, H.; Riederer, P. Levodopa in the treatment of Parkinson’s disease: Current controversies. Mov. Disord. 2005, 20, 643. [Google Scholar] [CrossRef]
- Nemade, D.; Subramanian, T.; Shivkumar, V. An Update on Medical and Surgical Treatments of Parkinson’s Disease. Aging Dis. 2021, 12, 1021–1035. [Google Scholar] [CrossRef]
- Tian, L.; Al-Nusaif, M.; Chen, X.; Li, S.; Le, W. Roles of Transcription Factors in the Development and Reprogramming of the Dopaminergic Neurons. Int. J. Mol. Sci. 2022, 23, 845. [Google Scholar] [CrossRef]
- Carmichael, K.; Sullivan, B.; Lopez, E.; Sun, L.; Cai, H. Diverse midbrain dopaminergic neuron subtypes and implications for complex clinical symptoms of Parkinson’s disease. Ageing Neurodegener Dis. 2021, 1. [Google Scholar] [CrossRef]
- Canals, I.; Quist, E.; Ahlenius, H. Transcription Factor-Based Strategies to Generate Neural Cell Types from Human Pluripotent Stem Cells. Cell Reprogram. 2021, 23, 206–220. [Google Scholar] [CrossRef]
- Mesman, S.; Smidt, M.P. Acquisition of the Midbrain Dopaminergic Neuronal Identity. Int. J. Mol. Sci. 2020, 21, 4638. [Google Scholar] [CrossRef]
- Azimi, S.M.; Sheridan, S.D.; Ghannad-Rezaie, M.; Eimon, P.M.; Yanik, M.F. Combinatorial programming of human neuronal progenitors using magnetically-guided stoichiometric mRNA delivery. Elife 2018, 7, e31922. [Google Scholar] [CrossRef]
- Di Val Cervo, P.R.; Romanov, R.A.; Spigolon, G.; Masini, D.; Martin-Montanez, E.; Toledo, E.M.; La Manno, G.; Feyder, M.; Pifl, C.; Ng, Y.-H.; et al. Induction of functional dopamine neurons from human astrocytes in vitro and mouse astrocytes in a Parkinson’s disease model. Nat. Biotechnol. 2017, 35, 444–452. [Google Scholar] [CrossRef]
- Ng, Y.H.; Chanda, S.; Janas, J.A.; Yang, N.; Kokubu, Y.; Südhof, T.C.; Wernig, M. Efficient generation of dopaminergic induced neuronal cells with midbrain characteristics. Stem Cell Reports 2021, 16, 1763–1776. [Google Scholar] [CrossRef] [PubMed]
- Smidt, M.P.; van Schaick, H.S.A.; Lanctôt, C.; Tremblay, J.J.; Cox, J.J.; van der Kleij, A.A.M.; Wolterink, G.; Drouin, J.; Burbach, J.P.H. A homeodomain gene Ptx3 has highly restricted brain expression in mesencephalic dopaminergic neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 13305–13310. [Google Scholar] [CrossRef] [PubMed]
- Smidt, M.P.; Smits, S.M.; Burbach, J.P. Homeobox gene Pitx3 and its role in the development of dopamine neurons of the substantia nigra. Cell Tissue Res. 2004, 318, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dani, J.A.; Le, W. The role of transcription factor Pitx3 in dopamine neuron development and Parkinson’s disease. Curr. Top. Med. Chem. 2009, 9, 855–859. [Google Scholar] [PubMed]
- Guo, Y.; Le, W.-D.; Jankovic, J.; Yang, H.-R.; Xu, H.-B.; Xie, W.-J.; Song, Z.; Deng, H. Systematic genetic analysis of the PITX3 gene in patients with Parkinson disease. Mov. Disord. 2011, 26, 1729–1732. [Google Scholar] [CrossRef]
- Kim, H.; Quan, X.; Seong, Y.; Kim, J. Impaired motor coordination in Pitx3 overexpression mice. Biochem. Biophys. Res. Commun. 2014, 446, 1211–1218. [Google Scholar] [CrossRef]
- Liu, H.; Wei, L.; Tao, Q.; Deng, H.; Ming, M.; Xu, P.; Le, W. Decreased NURR1 and PITX3 gene expression in Chinese patients with Parkinson’s disease. Eur. J. Neurol. 2012, 19, 870–875. [Google Scholar] [CrossRef]
- Le, W.; Nguyen, D.; Lin, X.W.; Rawal, P.; Huang, M.; Ding, Y.; Xie, W.; Deng, H.; Jankovic, J. Transcription factor PITX3 gene in Parkinson’s disease. Neurobiol. Aging 2011, 32, 750–753. [Google Scholar] [CrossRef]
- Anand, D.; Agrawal, S.A.; Slavotinek, A.; Lachke, S.A. Mutation update of transcription factor genes FOXE3, HSF4, MAF, and PITX3 causing cataracts and other developmental ocular defects. Hum. Mutat. 2018, 39, 471–494. [Google Scholar] [CrossRef]
- Semina, E.V.; Reiter, R.S.; Murray, J.C. Isolation of a new homeobox gene belonging to the Pitx/Rieg family: Expression during lens development and mapping to the aphakia region on mouse chromosome 19. Hum. Mol. Genet. 1997, 6, 2109–2116. [Google Scholar] [CrossRef]
- Messmer, K.; Remington, M.P.; Skidmore, F.; Fishman, P.S. Induction of tyrosine hydroxylase expression by the transcription factor Pitx3. Int. J. Dev. Neurosci. 2007, 25, 29–37. [Google Scholar] [CrossRef]
- Chen, H.; Song, Z.; Yuan, L.; Xiong, W.; Yang, Z.; Gong, L.; Deng, H. Genetic analysis of PITX3 variants in patients with essential tremor. Acta Neurol. Scand. 2017, 135, 373–376. [Google Scholar] [CrossRef]
- Lamonerie, T.; Tremblay, J.J.; Lanctôt, C.; Therrien, M.; Gauthier, Y.; Drouin, J. Ptx1, a bicoid-related homeo box transcription factor involved in transcription of the pro-opiomelanocortin gene. Genes Dev. 1996, 10, 1284–1295. [Google Scholar] [CrossRef]
- Gage, P.J.; Camper, S.A. Pituitary homeobox 2, a novel member of the bicoid-related family of homeobox genes, is a potential regulator of anterior structure formation. Hum. Mol. Genet. 1997, 6, 457–464. [Google Scholar] [CrossRef]
- Semina, E.V.; Ferrell, R.E.; Mintz-Hittner, H.A.; Bitoun, P.; Alward, W.L.M.; Reiter, R.S.; Funkhauser, C.; Daack-Hirsch, S.; Murray, J.C. A novel homeobox gene PITX3 is mutated in families with autosomal-dominant cataracts and ASMD. Nat. Genet. 1998, 19, 167–170. [Google Scholar] [CrossRef]
- Brouwer, A.; Berge, D.T.; Wiegerinck, R.; Meijlink, F. The OAR/aristaless domain of the homeodomain protein Cart1 has an attenuating role in vivo. Mech. Dev. 2003, 120, 241–252. [Google Scholar] [CrossRef]
- Bäckström, D.; Domellöf, M.E.; Granåsen, G.; Linder, J.; Mayans, S.; Elgh, E.; Mo, S.J.; Forsgren, L. PITX3 genotype and risk of dementia in Parkinson’s disease: A population-based study. J. Neurol. Sci. 2017, 381, 278–284. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Ho, H.Y.; Kuehner, E.; Zhao, S.; Li, M. Pitx3 regulates tyrosine hydroxylase expression in the substantia nigra and identifies a subgroup of mesencephalic dopaminergic progenitor neurons during mouse development. Dev. Biol. 2005, 282, 467–479. [Google Scholar] [CrossRef]
- Smidt, M.P.; Smits, S.M.; Bouwmeester, H.; Hamers, F.P.T.; van der Linden, A.J.A.; Hellemons, A.J.C.G.M.; Graw, J.; Burbach, J.P.H. Early developmental failure of substantia nigra dopamine neurons in mice lacking the homeodomain gene Pitx3. Development 2004, 131, 1145–1155. [Google Scholar] [CrossRef]
- Nunes, I.; Tovmasian, L.T.; Silva, R.M.; Burke, R.E.; Goff, S.P. Pitx3 is required for development of substantia nigra dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2003, 100, 4245–4250. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Wang, Y.; Li, S.; Cai, H.; Le, W. The essential role of transcription factor Pitx3 in preventing mesodiencephalic dopaminergic neurodegeneration and maintaining neuronal subtype identities during aging. Cell Death Dis. 2021, 12, 1008. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.M.; Burbach, J.P.; Smidt, M.P. Developmental origin and fate of meso-diencephalic dopamine neurons. Prog. Neurobiol. 2006, 78, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.Q.; Kioussi, C. Pitx genes in development and disease. Cell Mol. Life Sci. 2021, 78, 4921–4938. [Google Scholar] [CrossRef] [PubMed]
- Ang, S.L. Transcriptional control of midbrain dopaminergic neuron development. Development 2006, 133, 3499–3506. [Google Scholar] [CrossRef] [PubMed]
- Smidt, M.; Asbreuk, C.H.J.; Cox, J.J.; Chen, H.; Johnson, R.L.; Burbach, J.P.H. A second independent pathway for development of mesencephalic dopaminergic neurons requires Lmx1b. Nat. Neurosci. 2000, 3, 337–341. [Google Scholar] [CrossRef]
- Andersson, E.; Tryggvason, U.; Deng, Q.; Friling, S.; Alekseenko, Z.; Robert, B.; Perlmann, T.; Ericson, J. Identification of intrinsic determinants of midbrain dopamine neurons. Cell 2006, 124, 393–405. [Google Scholar] [CrossRef]
- Alavian, K.N.; Scholz, C.; Simon, H.H. Transcriptional regulation of mesencephalic dopaminergic neurons: The full circle of life and death. Mov. Disord. 2008, 23, 319–328. [Google Scholar] [CrossRef]
- Doucet-Beaupré, H.; Gilbert, C.; Profes, M.S.; Chabrat, A.; Pacelli, C.; Giguère, N.; Rioux, V.; Charest, J.; Deng, Q.; Laguna, A.; et al. Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2016, 113, E4387–E4396. [Google Scholar] [CrossRef]
- Chung, S.; Leung, A.; Han, B.S.; Chang, M.Y.; Moon, J.I.; Kim, C.H.; Hong, S.; Pruszak, J.; Isacson, O.; Kim, K.S. Wnt1-lmx1a forms a novel autoregulatory loop and controls midbrain dopaminergic differentiation synergistically with the SHH-FoxA2 pathway. Cell Stem Cell 2009, 5, 646–658. [Google Scholar] [CrossRef]
- Villaescusa, J.C.; Li, B.; Toledo, E.M.; Rivetti di Val Cervo, P.; Yang, S.; Stott, S.R.; Kaiser, K.; Islam, S.; Gyllborg, D.; Laguna-Goya, R.; et al. A PBX1 transcriptional network controls dopaminergic neuron development and is impaired in Parkinson’s disease. EMBO J. 2016, 35, 1963–1978. [Google Scholar] [CrossRef]
- Nouri, P.; Götz, S.; Rauser, B.; Irmler, M.; Peng, C.; Trümbach, D.; Kempny, C.; Lechermeier, C.G.; Bryniok, A.; Dlugos, A.; et al. Dose-Dependent and Subset-Specific Regulation of Midbrain Dopaminergic Neuron Differentiation by LEF1-Mediated WNT1/b-Catenin Signaling. Front. Cell Dev. Biol. 2020, 8, 587778. [Google Scholar] [CrossRef]
- Andersson, E.R.; Saltó, C.; Villaescusa, J.C.; Cajanek, L.; Yang, S.; Bryjova, L.; Nagy, I.I.; Vainio, S.J.; Ramirez, C.; Bryja, V.; et al. Wnt5a cooperates with canonical Wnts to generate midbrain dopaminergic neurons in vivo and in stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, E602–E610. [Google Scholar] [CrossRef]
- Prakash, N.; Brodski, C.; Naserke, T.; Puelles, E.; Gogoi, R.; Hall, A.; Panhuysen, M.; Echevarria, D.; Sussel, L.; Weisenhorn, D.M.V.; et al. A Wnt1-regulated genetic network controls the identity and fate of midbrain-dopaminergic progenitors in vivo. Development 2006, 133, 89–98. [Google Scholar] [CrossRef]
- Zetterstrom, R.H.; Solomin, L.; Jansson, L.; Hoffer, B.J.; Olson, L.; Perlmann, T. Dopamine neuron agenesis in Nurr1-deficient mice. Science 1997, 276, 248–250. [Google Scholar] [CrossRef]
- Saucedo-Cardenas, O.; Quintana-Hau, J.D.; Le, W.-D.; Smidt, M.P.; Cox, J.J.; De Mayo, F.; Burbach, J.P.H.; Conneely, O.M. Nurr1 is essential for the induction of the dopaminergic phenotype and the survival of ventral mesencephalic late dopaminergic precursor neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 4013–4018. [Google Scholar] [CrossRef]
- Jiang, C.; Wan, X.; He, Y.; Pan, T.; Jankovic, J.; Le, W. Age-dependent dopaminergic dysfunction in Nurr1 knockout mice. Exp. Neurol. 2005, 191, 154–162. [Google Scholar] [CrossRef]
- Pulcrano, S.; De Gregorio, R.; De Sanctis, C.; Lahti, L.; Perrone-Capano, C.; Ponti, D.; di Porzio, U.; Perlmann, T.; Caiazzo, M.; Volpicelli, F.; et al. Lmx1a-Dependent Activation of miR-204/211 Controls the Timing of Nurr1-Mediated Dopaminergic Differentiation. Int. J. Mol. Sci. 2022, 23, 6961. [Google Scholar] [CrossRef]
- Argyrofthalmidou, M.; Spathis, A.D.; Maniati, M.; Poula, A.; Katsianou, M.A.; Sotiriou, E.; Manousaki, M.; Perier, C.; Papapanagiotou, I.; Papadopoulou-Daifoti, Z.; et al. Nurr1 repression mediates cardinal features of Parkinson’s disease in alpha-synuclein transgenic mice. Hum. Mol. Genet. 2021, 30, 1469–1483. [Google Scholar] [CrossRef]
- García-Yagüe, J.; Lastres-Becker, I.; Stefanis, L.; Vassilatis, D.K.; Cuadrado, A. alpha-Synuclein Induces the GSK-3-Mediated Phosphorylation and Degradation of NURR1 and Loss of Dopaminergic Hallmarks. Mol. Neurobiol. 2021, 58, 6697–6711. [Google Scholar] [CrossRef]
- Katunar, M.R.; Saez, T.; Brusco, A.; Antonelli, M.C. Immunocytochemical expression of dopamine-related transcription factors Pitx3 and Nurr1 in prenatally stressed adult rats. J. Neurosci. Res. 2009, 87, 1014–1022. [Google Scholar] [CrossRef]
- Le, W.; Pan, T.; Huang, M.; Xu, P.; Xie, W.; Zhu, W.; Zhang, X.; Deng, H.; Jankovic, J. Decreased NURR1 gene expression in patients with Parkinson’s disease. J. Neurol. Sci. 2008, 273, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Martinat, C.; Bacci, J.-J.; Leete, T.; Kim, J.; Vanti, W.B.; Newman, A.H.; Cha, J.H.; Gether, U.; Wang, H.; Abeliovich, A. Cooperative transcription activation by Nurr1 and Pitx3 induces embryonic stem cell maturation to the midbrain dopamine neuron phenotype. Proc. Natl. Acad. Sci. USA 2006, 103, 2874–2879. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, F.M.J.; van Erp, S.; van der Linden, A.J.A.; von Oerthel, L.; Burbach, J.P.H.; Smidt, M.P. Pitx3 potentiates Nurr1 in dopamine neuron terminal differentiation through release of SMRT-mediated repression. Development 2009, 136, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Rekaik, H.; de Thé, F.-X.B.; Prochiantz, A.; Fuchs, J.; Joshi, R.L. Dissecting the role of Engrailed in adult dopaminergic neurons--Insights into Parkinson disease pathogenesis. FEBS Lett. 2015, 589 Pt A, 3786–3794. [Google Scholar] [CrossRef]
- Prochiantz, A.; Di Nardo, A.A. Homeoprotein signaling in the developing and adult nervous system. Neuron 2015, 85, 911–925. [Google Scholar] [CrossRef]
- Simon, H.H.; Saueressig, H.; Wurst, W.; Goulding, M.D.; O'Leary, D.D.M. Fate of midbrain dopaminergic neurons controlled by the engrailed genes. J. Neurosci. 2001, 21, 3126–3134. [Google Scholar] [CrossRef]
- Nordströma, U.; Beauvais, G.; Ghosh, A.; Pulikkaparambil Sasidharan, B.C.; Lundblad, M.; Fuchs, J.; Joshi, R.L.; Lipton, J.W.; Roholt, A.; Medicetty, S.; et al. Progressive nigrostriatal terminal dysfunction and degeneration in the engrailed1 heterozygous mouse model of Parkinson’s disease. Neurobiol. Dis. 2015, 73, 70–82. [Google Scholar] [CrossRef]
- Blaudin de The, F.X.; Rekaik, H.; Prochiantz, A.; Fuchs, J.; Joshi, R.L. Neuroprotective Transcription Factors in Animal Models of Parkinson Disease. Neural. Plast. 2016, 2016, 6097107. [Google Scholar] [CrossRef]
- Danielian, P.S.; McMahon, A.P. Engrailed-1 as a target of the Wnt-1 signalling pathway in vertebrate midbrain development. Nature 1996, 383, 332–334. [Google Scholar] [CrossRef]
- Veenvliet, J.V.; dos Santos, M.T.M.A.; Kouwenhoven, W.M.; von Oerthel, L.; Lim, J.L.; van der Linden, A.J.A.; Koerkamp, M.J.A.G.; Holstege, F.C.P.; Smidt, M.P. Specification of dopaminergic subsets involves interplay of En1 and Pitx3. Development 2013, 140, 3373–3384. [Google Scholar] [CrossRef]
- Haubenberger, D.; Reinthaler, E.; Mueller, J.C.; Pirker, W.; Katzenschlager, R.; Froehlich, R.; Bruecke, T.; Daniel, G.; Auff, E.; Zimprich, A. Association of transcription factor polymorphisms PITX3 and EN1 with Parkinson’s disease. Neurobiol. Aging 2011, 32, 302–307. [Google Scholar] [CrossRef]
- Jackson, B.C.; Carpenter, C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse forkhead box (FOX) gene families. Hum. Genomics. 2010, 4, 345–352. [Google Scholar] [CrossRef]
- Ang, S.L. Foxa1 and Foxa2 transcription factors regulate differentiation of midbrain dopaminergic neurons. Adv. Exp. Med. Biol. 2009, 651, 58–65. [Google Scholar]
- Pristerà, A.; Lin, W.; Kaufmann, A.-K.; Brimblecombe, K.R.; Threlfell, S.; Dodson, P.D.; Magill, P.J.; Fernandes, C.; Cragg, S.J.; Ang, S.-L. Transcription factors FOXA1 and FOXA2 maintain dopaminergic neuronal properties and control feeding behavior in adult mice. Proc. Natl. Acad. Sci. USA 2015, 112, E4929–E4938. [Google Scholar] [CrossRef]
- Sasaki, H.; Hogan, B.L. Differential expression of multiple fork head related genes during gastrulation and axial pattern formation in the mouse embryo. Development 1993, 118, 47–59. [Google Scholar] [CrossRef]
- Burtscher, I.; Lickert, H. Foxa2 regulates polarity and epithelialization in the endoderm germ layer of the mouse embryo. Development 2009, 136, 1029–1038. [Google Scholar] [CrossRef]
- Shih, D.Q.; Navas, M.A.; Kuwajima, S.; Duncan, S.A.; Stoffel, M. Impaired glucose homeostasis and neonatal mortality in hepatocyte nuclear factor 3alpha-deficient mice. Proc. Natl. Acad. Sci. USA 1999, 96, 10152–10157. [Google Scholar] [CrossRef]
- Stott, S.R.W.; Metzakopian, E.; Lin, W.; Kaestner, K.H.; Hen, R.; Ang, S.-L. Foxa1 and foxa2 are required for the maintenance of dopaminergic properties in ventral midbrain neurons at late embryonic stages. J. Neurosci. 2013, 33, 8022–8034. [Google Scholar] [CrossRef]
- Kittappa, R.; Chang, W.W.; Awatramani, R.B.; McKay, R.D.G. The foxa2 gene controls the birth and spontaneous degeneration of dopamine neurons in old age. PLoS. Biol. 2007, 5, e325. [Google Scholar] [CrossRef]
- Oh, S.-M.; Chang, M.-Y.; Song, J.-J.; Rhee, Y.-H.; Joe, E.-H.; Lee, H.-S.; Yi, S.-H.; Lee, S.-H. Combined Nurr1 and Foxa2 roles in the therapy of Parkinson’s disease. EMBO Mol. Med. 2015, 7, 510–525. [Google Scholar] [CrossRef]
- Lin, W.; Metzakopian, E.; Mavromatakis, Y.E.; Gao, N.; Balaskas, N.; Sasaki, H.; Briscoe, J.; Whitsett, J.A.; Goulding, M.; Kaestner, K.H.; et al. Foxa1 and Foxa2 function both upstream of and cooperatively with Lmx1a and Lmx1b in a feedforward loop promoting mesodiencephalic dopaminergic neuron development. Dev. Biol. 2009, 333, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Wallen, A.; Perlmann, T. Transcriptional control of dopamine neuron development. Ann. N. Y. Acad. Sci. 2003, 991, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Blandini, F.; Armentero, M.T. Animal models of Parkinson’s disease. FEBS J. 2012, 279, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Vingill, S.; Connor-Robson, N.; Wade-Martins, R. Are rodent models of Parkinson’s disease behaving as they should? Behav. Brain. Res. 2018, 352, 133–141. [Google Scholar] [CrossRef]
- Dawson, T.M.; Ko, H.S.; Dawson, V.L. Genetic animal models of Parkinson’s disease. Neuron 2010, 66, 646–661. [Google Scholar] [CrossRef]
- Chesselet, M.F.; Richter, F. Modelling of Parkinson’s disease in mice. Lancet Neurol. 2011, 10, 1108–1118. [Google Scholar] [CrossRef]
- Langley, M.R.; Ghaisas, S.; Palanisamy, B.N.; Ay, M.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Characterization of nonmotor behavioral impairments and their neurochemical mechanisms in the MitoPark mouse model of progressive neurodegeneration in Parkinson’s disease. Exp. Neurol. 2021, 341, 113716. [Google Scholar] [CrossRef]
- van den Munckhof, P.; Gilbert, F.; Chamberland, M.; Levesque, D.; Drouin, J. Striatal neuroadaptation and rescue of locomotor deficit by L-dopa in aphakia mice, a model of Parkinson’s disease. J. Neurochem. 2006, 96, 160–170. [Google Scholar] [CrossRef]
- Song, B.; Feldmann, J.W.; Cao, S.; Feitosa, M.; Kong, Y.; Kim, W.; Schweitzer, A.; Leblanc, P.; Schweitzer, J.S.; Kim, K.-S. A Pitx3-deficient developmental mouse model for fine motor, olfactory, and gastrointestinal symptoms of Parkinson’s disease. Neurobiol. Dis. 2022, 170, 105777. [Google Scholar] [CrossRef]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef]
- Le, W.; Zhang, L.; Xie, W.; Li, S.; Dani, J.A. Pitx3 deficiency produces decreased dopamine signaling and induces motor deficits in Pitx3(-/-) mice. Neurobiol. Aging 2015, 36, 3314–3320. [Google Scholar] [CrossRef]
- Chen, X.; Yang, Z.; Shao, Y.; Kim, K.; Wang, Y.; Wang, Y.; Wu, H.; Xu, X.; Le, W. Pitx3 deficiency promotes age-dependent alterations in striatal medium spiny neurons. Front. Aging Neurosci. 2022, 14, 960479. [Google Scholar] [CrossRef]
- Suarez, L.M.; Alberquilla, S.; García-Montes, J.R.; Moratalla, R. Differential Synaptic Remodeling by Dopamine in Direct and Indirect Striatal Projection Neurons in Pitx3(-/-) Mice, a Genetic Model of Parkinson’s Disease. J. Neurosci. 2018, 38, 3619–3630. [Google Scholar] [CrossRef]
- Mitalipov, S.; Wolf, D. Totipotency, pluripotency and nuclear reprogramming. Adv. Biochem. Eng. Biotechnol. 2009, 114, 185–199. [Google Scholar]
- Wang, M.; Ling, K.-H.; Tan, J.J.; Lu, C.-B. Development and Differentiation of Midbrain Dopaminergic Neuron: From Bench to Bedside. Cells 2020, 9, 1489. [Google Scholar] [CrossRef]
- Paquet-Durand, F.; Bicker, G. Human model neurons in studies of brain cell damage and neural repair. Curr. Mol. Med. 2007, 7, 541–554. [Google Scholar] [CrossRef]
- Pleasure, S.J.; Lee, V.M. NTera 2 cells: A human cell line which displays characteristics expected of a human committed neuronal progenitor cell. J. Neurosci. Res. 1993, 35, 585–602. [Google Scholar] [CrossRef]
- Eskandarian Boroujeni, M.; Aliaghaei, A.; Maghsoudi, N.; Gardaneh, M. Complementation of dopaminergic signaling by Pitx3-GDNF synergy induces dopamine secretion by multipotent Ntera2 cells. J. Cell Biochem. 2020, 121, 200–212. [Google Scholar] [CrossRef]
- Wegmeyer, H.; Bröske, A.-M.; Leddin, M.; Kuentzer, K.; Nisslbeck, A.K.; Hupfeld, J.; Wiechmann, K.; Kuhlen, J.; Von Schwerin, C.; Stein, C.; et al. Mesenchymal stromal cell characteristics vary depending on their origin. Stem Cells Dev. 2013, 22, 2606–2618. [Google Scholar] [CrossRef]
- Trzaska, K.A.; Kuzhikandathil, E.V.; Rameshwar, P. Specification of a dopaminergic phenotype from adult human mesenchymal stem cells. Stem Cells 2007, 25, 2797–2808. [Google Scholar] [CrossRef]
- Suon, S.; Yang, M.; Iacovitti, L. Adult human bone marrow stromal spheres express neuronal traits in vitro and in a rat model of Parkinson’s disease. Brain Res. 2006, 1106, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Hermann, A.; List, C.; Habisch, H.-J.; Vukicevic, V.; Ehrhart-Bornstein, M.; Brenner, R.; Bernstein, P.; Fickert, S.; Storch, A. Age-dependent neuroectodermal differentiation capacity of human mesenchymal stromal cells: Limitations for autologous cell replacement strategies. Cytotherapy 2010, 12, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Hedlund, E.; Hwang, M.; Kim, D.; Shin, B.-S.; Hwang, D.-Y.; Kang, U.J.; Isacson, O.; Kim, K.-S. The homeodomain transcription factor Pitx3 facilitates differentiation of mouse embryonic stem cells into AHD2-expressing dopaminergic neurons. Mol. Cell Neurosci. 2005, 28, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, E.; Pruszak, J.; Lardaro, T.; Ludwig, W.; Viñuela, A.; Kim, K.-S.; Isacson, O. Embryonic stem cell-derived Pitx3-enhanced green fluorescent protein midbrain dopamine neurons survive enrichment by fluorescence-activated cell sorting and function in an animal model of Parkinson’s disease. Stem Cells 2008, 26, 1526–3156. [Google Scholar] [CrossRef]
- Brundin, P.; Karlsson, J.; Emgård, M.; Schierle, G.S.K.; Hansson, O.; Petersén, A.; Castilho, R.F. Improving the survival of grafted dopaminergic neurons: A review over current approaches. Cell Transplant. 2000, 9, 179–195. [Google Scholar] [CrossRef]
- Haas, S.J.-P.; Beckmann, S.; Petrov, S.; Andressen, C.; Wree, A.; Schmitt, O. Transplantation of immortalized mesencephalic progenitors (CSM14.1 cells) into the neonatal parkinsonian rat caudate putamen. J. Neurosci. Res. 2007, 85, 778–786. [Google Scholar] [CrossRef]
- Hong, S.; Chung, S.; Leung, K.; Hwang, I.; Moon, J.; Kim, K.-S. Functional roles of Nurr1, Pitx3, and Lmx1a in neurogenesis and phenotype specification of dopamine neurons during in vitro differentiation of embryonic stem cells. Stem Cells Dev. 2014, 23, 477–487. [Google Scholar] [CrossRef]
- Tiklová, K.; Björklund, Å.K.; Lahti, L.; Fiorenzano, A.; Nolbrant, S.; Gillberg, L.; Volakakis, N.; Yokota, C.; Hilscher, M.M.; Hauling, T.; et al. Single-cell RNA sequencing reveals midbrain dopamine neuron diversity emerging during mouse brain development. Nat. Commun. 2019, 10, 581. [Google Scholar] [CrossRef]
- Fuchs, J.; Mueller, J.C.; Lichtner, P.; Schulte, C.; Munz, M.; Berg, D.; Wüllner, U.; Illig, T.; Sharma, M.; Gasser, T. The transcription factor PITX3 is associated with sporadic Parkinson’s disease. Neurobiol. Aging 2009, 30, 731–738. [Google Scholar] [CrossRef]
- Bergman, O.; Håkansson, A.; Westberg, L.; Nordenström, K.; Belin, A.C.; Sydow, O.; Olson, L.; Holmberg, B.; Eriksson, E.; Nissbrandt, H. PITX3 polymorphism is associated with early onset Parkinson’s disease. Neurobiol. Aging 2010, 31, 114–117. [Google Scholar] [CrossRef]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef]
- Prakash, N. Developmental pathways linked to the vulnerability of adult midbrain dopaminergic neurons to neurodegeneration. Front. Mol. Neurosci. 2022, 15, 1071731. [Google Scholar] [CrossRef]
- Yang, D.; Peng, C.; Li, X.; Fan, X.; Li, L.; Ming, M.; Chen, S.; Le, W. Pitx3-transfected astrocytes secrete brain-derived neurotrophic factor and glial cell line-derived neurotrophic factor and protect dopamine neurons in mesencephalon cultures. J. Neurosci. Res. 2008, 86, 3393–3400. [Google Scholar] [CrossRef]
- Håkansson, A.; Westberg, L.; Nilsson, S.; Buervenich, S.; Carmine, A.; Holmberg, B.; Sydow, O.; Olson, L.; Johnels, B.; Eriksson, E.; et al. Interaction of polymorphisms in the genes encoding interleukin-6 and estrogen receptor beta on the susceptibility to Parkinson’s disease. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 133, 88–92. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Transcription Factor | The Expression Time Embryonic Day (E) | The Cells It Acts on | The Distribution Region | Reference |
|---|---|---|---|---|
| Pitx3 | E11.5 | Mature mDA neurons | Substantia nigra and ventral tegmental region | [23,61] |
| Lmx1a | E9 | Mitotic ventral mesencephalic precursor cells | Ventral midbrain and dorsally | [46,47] |
| Lmx1b | E7.5 | Neural progenitor cells, Mitotic ventral mesencephalic precursor cells | Derive from the midbrain, then restricted to the ventral midbrain, the floor plate, mid-hindbrain boundary, and roof plate | [46,47] |
| Nurr1 | E10.5 | Immature/ mature mDA neurons | Cerebral cortex, hippocampus, thalamus, amygdala, and midbrain | [61,83] |
| En1/2 | E8 | Ventral mDA precursor | Midbrain and hindbrain | [65,66,67] |
| Foxa 1 | E7.5 | mDA progenitors, immature/ mature neurons | Floorplate, notochord, and endoderm | [76,77,79] |
| Foxa 2 | E6.5 | mDA progenitors, immature/ mature neurons | Posterior epiblast, anterior definitive endoderm, axial mesoderm | [76,77,79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chen, X.; Liu, G.; Cai, H.; Le, W. The Crucial Roles of Pitx3 in Midbrain Dopaminergic Neuron Development and Parkinson’s Disease-Associated Neurodegeneration. Int. J. Mol. Sci. 2023, 24, 8614. https://doi.org/10.3390/ijms24108614
Wang X, Chen X, Liu G, Cai H, Le W. The Crucial Roles of Pitx3 in Midbrain Dopaminergic Neuron Development and Parkinson’s Disease-Associated Neurodegeneration. International Journal of Molecular Sciences. 2023; 24(10):8614. https://doi.org/10.3390/ijms24108614
Chicago/Turabian StyleWang, Xin, Xi Chen, Guangdong Liu, Huaibin Cai, and Weidong Le. 2023. "The Crucial Roles of Pitx3 in Midbrain Dopaminergic Neuron Development and Parkinson’s Disease-Associated Neurodegeneration" International Journal of Molecular Sciences 24, no. 10: 8614. https://doi.org/10.3390/ijms24108614