
NPAS4 Exacerbates Pyroptosis via Transcriptionally Regulating NLRP6 in the Acute Phase of Intracerebral Hemorrhage in Mice
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
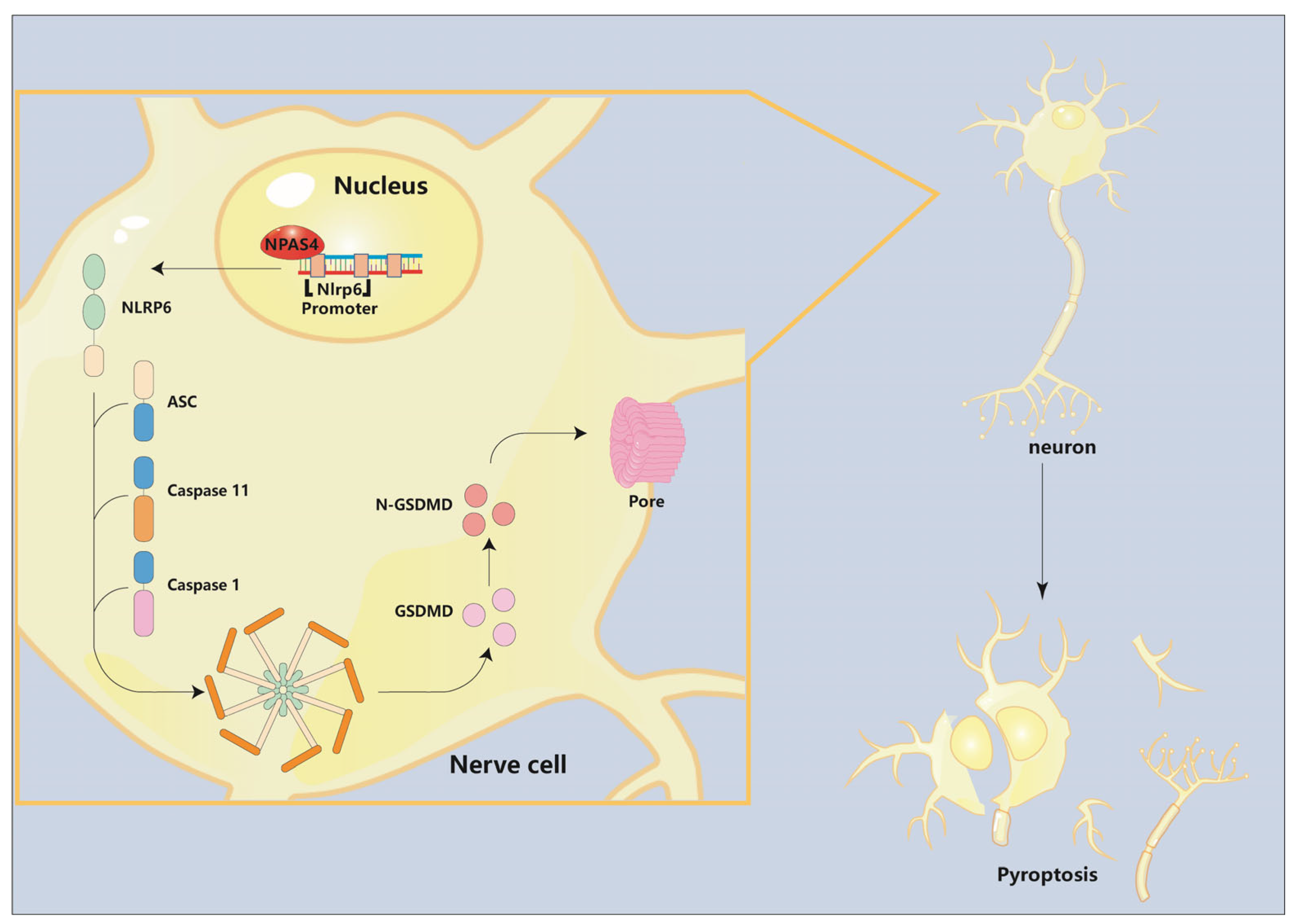
Abstract
:1. Introduction
2. Results
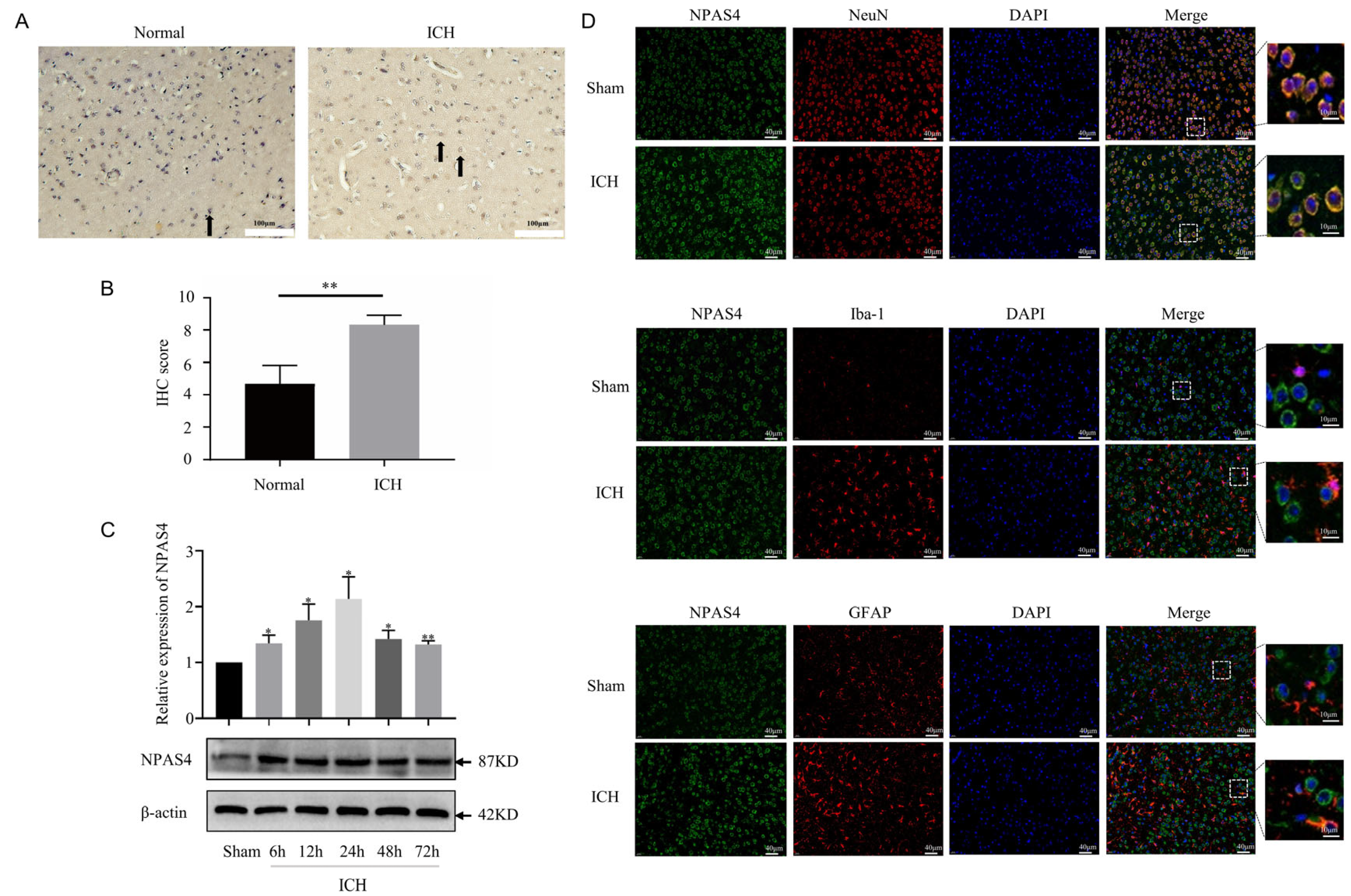
2.1. The Spatiotemporal Expression of NPAS4 after ICH
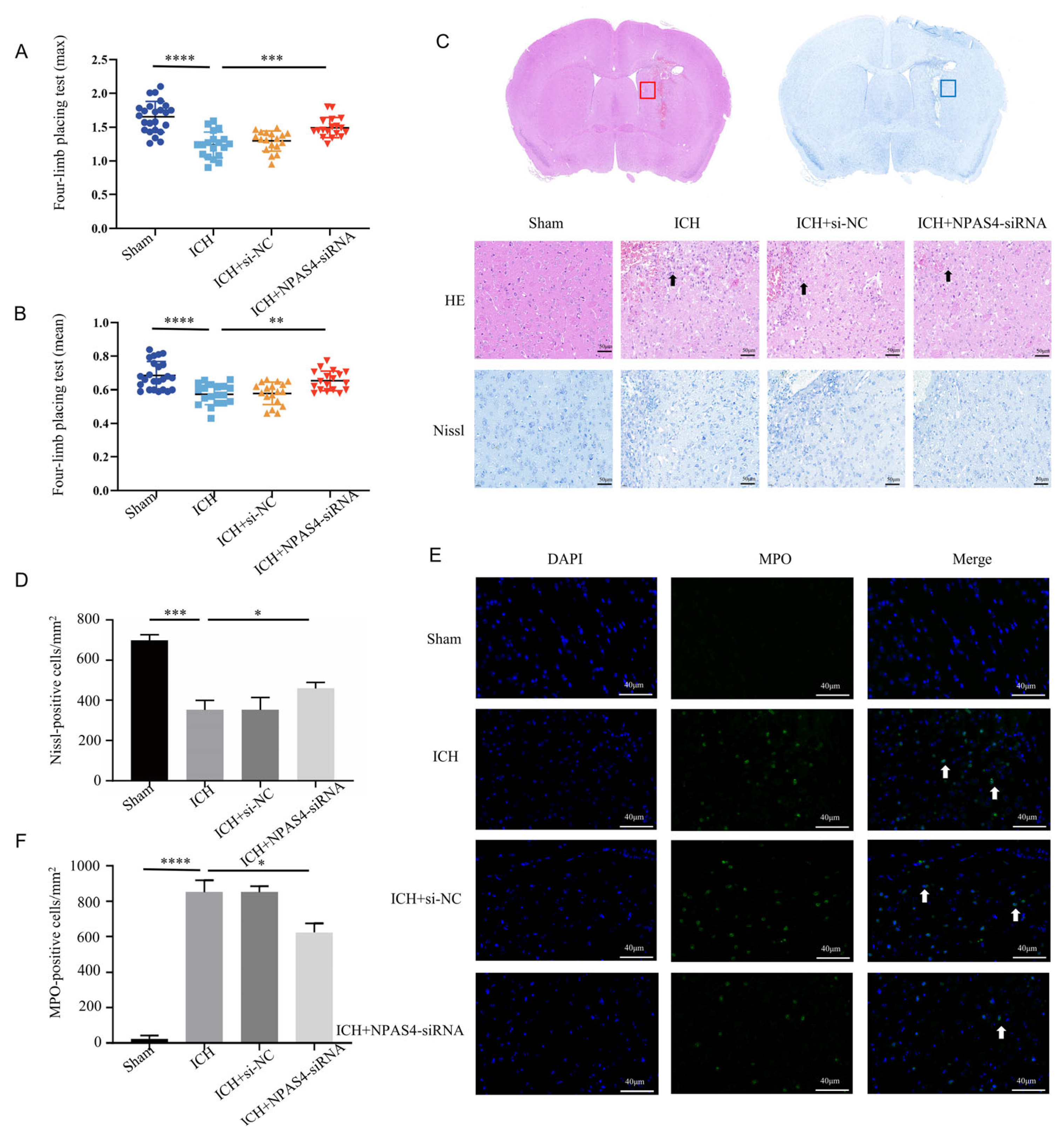
2.2. NAPS4 Deficiency Attenuated Neurological Function Deficits, Morphologic Damage, and Neutrophil Infiltration after ICH in Mice
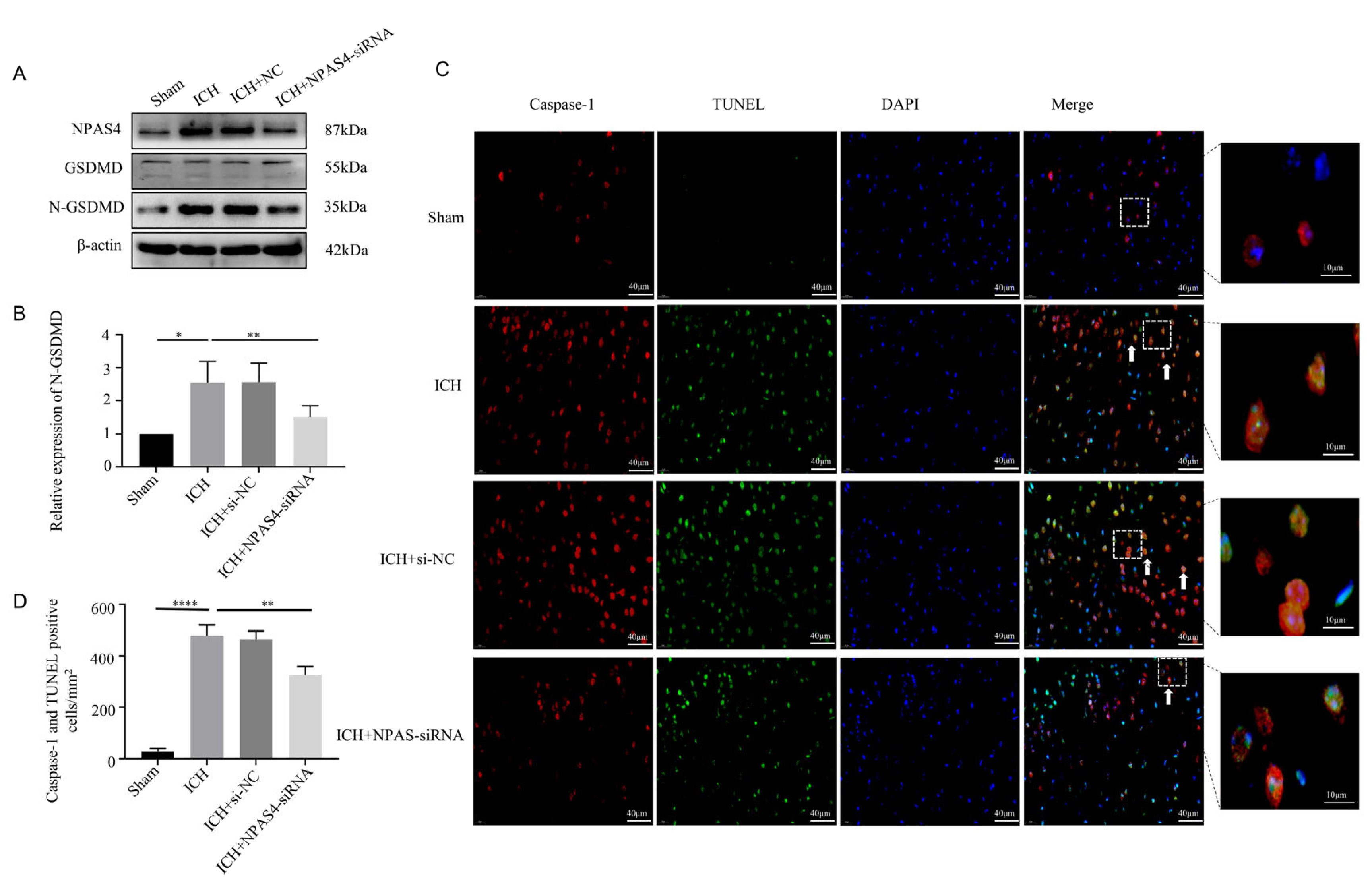
2.3. NAPS4 Deficiency Lessened Pyroptosis after ICH Modeling
2.4. The Interaction of NPAS4 and Nlrp6 Promoted Inflammasome Activation after ICH
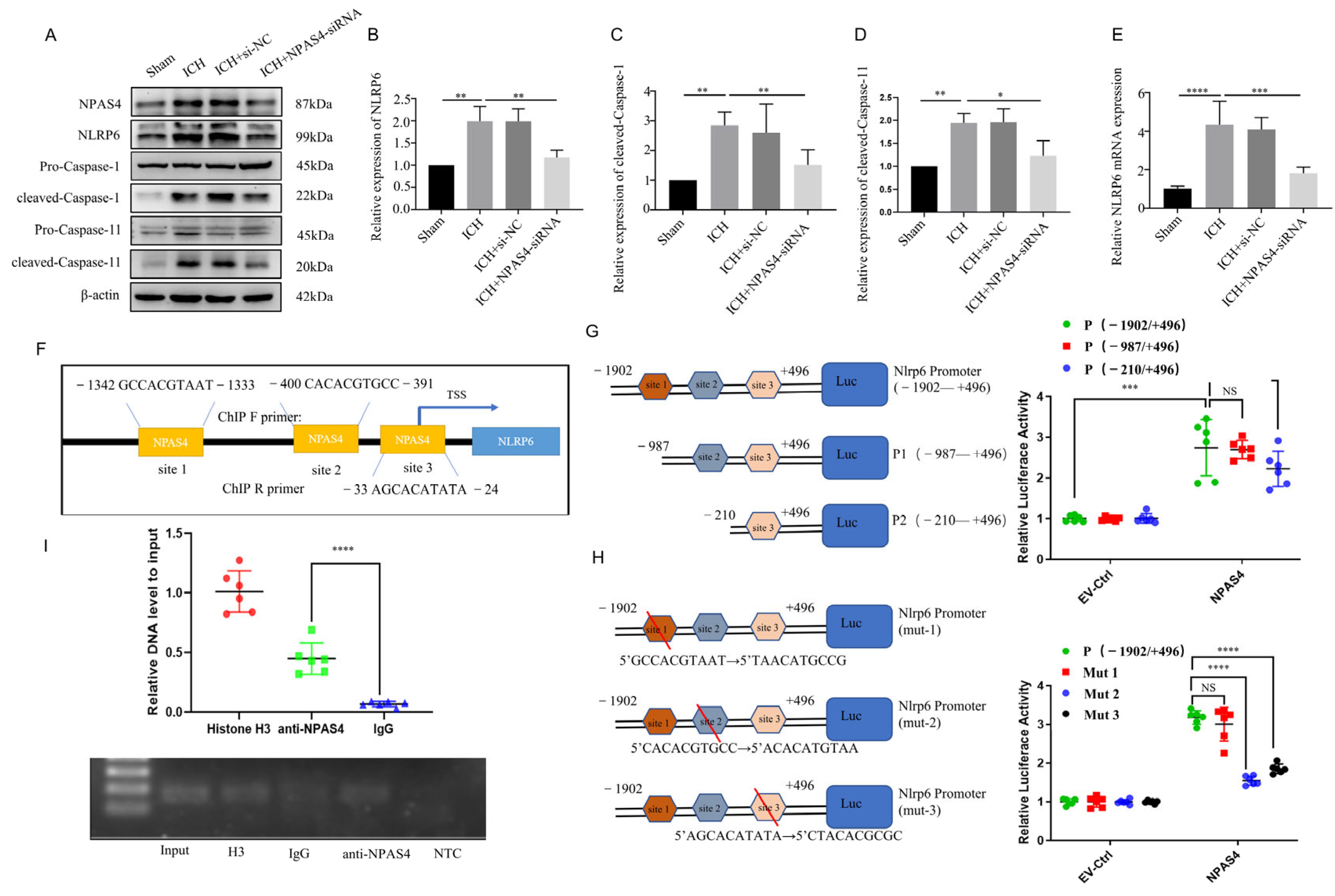
2.4.1. NPAS4 Deficiency Inhibited the Activation of NLRP6 Inflammasome after ICH
2.4.2. The Interaction of NPAS4 and Nlrp6 Promotor
2.5. NPAS4 Overexpression Aggravated Neuron Death, Neurological Deficits, and Cell Pyroptosis following ICH
3. Discussion
4. Materials and Methods
4.1. Human Tissues
4.2. Animals
4.3. ICH Model
4.4. siRNA Transfections
4.5. Adeno-Associated Virus Transfection
4.6. Grip Strength Test
4.7. Hematoxylin and Eosin (H&E) Staining
4.8. Nissl Staining
4.9. Immunofluorescence Staining
4.10. Immunohistochemical Staining
4.11. Western Blotting
4.12. Quantitative Real-Time PCR (qRT-PCR)
4.13. Plasmids Construction
4.14. Dual Luciferase Reporter Gene Assay
4.15. Chromatin Immunoprecipitation (ChIP) Assay
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liddle, L.J.; Ralhan, S.; Ward, D.L.; Colbourne, F. Translational Intracerebral Hemorrhage Research: Has Current Neuroprotection Research ARRIVEd at a Standard for Experimental Design and Reporting? Transl. Stroke Res. 2020, 11, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Gang, X.; Han, Q.; Zhao, X.; Liu, Q.; Wang, Y. Dynamic Changes in Toll-Like Receptor 4 in Human Perihematoma Tissue after Intracerebral Hemorrhage. World Neurosurg. 2018, 118, e593–e600. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Tu, S.; Shao, A. Pathophysiological Mechanisms and Potential Therapeutic Targets in Intracerebral Hemorrhage. Front. Pharmacol. 2019, 10, 1079. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Aronowski, J. Nrf2 to pre-condition the brain against injury caused by products of hemolysis after ICH. Transl. Stroke Res. 2013, 4, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Khan, S.; Liu, Y.; Zhang, R.; Li, H.; Wu, G.; Tang, Z.; Xue, M.; Yong, V.W. Modes of Brain Cell Death Following Intracerebral Hemorrhage. Front. Cell Neurosci. 2022, 16, 799753. [Google Scholar] [CrossRef]
- Xi, G.; Strahle, J.; Hua, Y.; Keep, R.F. Progress in translational research on intracerebral hemorrhage: Is there an end in sight? Prog. Neurobiol. 2014, 115, 45–63. [Google Scholar] [CrossRef]
- Ren, S.; Chen, Y.; Wang, L.; Wu, G. Neuronal ferroptosis after intracerebral hemorrhage. Front. Mol. Biosci. 2022, 9, 966478. [Google Scholar] [CrossRef]
- Matsushita, K.; Meng, W.; Wang, X.; Asahi, M.; Asahi, K.; Moskowitz, M.A.; Lo, E.H. Evidence for apoptosis after intercerebral hemorrhage in rat striatum. J. Cereb. Blood Flow. Metab. 2000, 20, 396–404. [Google Scholar] [CrossRef]
- Zhu, X.; Tao, L.; Tejima-Mandeville, E.; Qiu, J.; Park, J.; Garber, K.; Ericsson, M.; Lo, E.H.; Whalen, M.J. Plasmalemma permeability and necrotic cell death phenotypes after intracerebral hemorrhage in mice. Stroke 2012, 43, 524–531. [Google Scholar] [CrossRef]
- Zille, M.; Karuppagounder, S.S.; Chen, Y.; Gough, P.J.; Bertin, J.; Finger, J.; Milner, T.A.; Jonas, E.A.; Ratan, R.R. Neuronal Death After Hemorrhagic Stroke In Vitro and In Vivo Shares Features of Ferroptosis and Necroptosis. Stroke 2017, 48, 1033–1043. [Google Scholar] [CrossRef]
- Li, Q.; Han, X.; Lan, X.; Gao, Y.; Wan, J.; Durham, F.; Cheng, T.; Yang, J.; Wang, Z.; Jiang, C.; et al. Inhibition of neuronal ferroptosis protects hemorrhagic brain. J. Clin. Investig. 2017, 2, e90777. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal. Transduct. Target. Ther. 2021, 6, 128. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Galloway, N.L.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Muñoz-Arias, I.; et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kanneganti, T.D. Regulation of inflammasome activation. Immunol. Rev. 2015, 265, 6–21. [Google Scholar] [CrossRef]
- Ruan, J.; Wang, S.; Wang, J. Mechanism and regulation of pyroptosis-mediated in cancer cell death. Chem. Biol. Interact. 2020, 323, 109052. [Google Scholar] [CrossRef]
- Man, S.M.; Karki, R.; Kanneganti, T.D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol. Rev. 2017, 277, 61–75. [Google Scholar] [CrossRef]
- Levy, M.; Shapiro, H.; Thaiss, C.A.; Elinav, E. NLRP6: A Multifaceted Innate Immune Sensor. Trends Immunol. 2017, 38, 248–260. [Google Scholar] [CrossRef]
- Henao-Mejia, J.; Elinav, E.; Thaiss, C.A.; Flavell, R.A. Inflammasomes and metabolic disease. Annu. Rev. Physiol. 2014, 76, 57–78. [Google Scholar] [CrossRef]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 278–286. [Google Scholar] [CrossRef]
- Lima, C.; Falcao MA, P.; Andrade-Barros, A.I.; Seni-Silva, A.C.; Grund, L.Z.; Balogh, E.; Conceiçao, K.; Queniaux, V.F.; Ryffel, B.; Lopes-Ferreira, M. Natterin an aerolysin-like fish toxin drives IL-1β-dependent neutrophilic inflammation mediated by caspase-1 and caspase-11 activated by the inflammasome sensor NLRP6. Int. Immunopharmacol. 2021, 91, 107287. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Seregin, S.S.; Yang, D.; Fukase, K.; Chamaillard, M.; Alnemri, E.S.; Inohara, N.; Chen, G.Y.; Núñez, G. The NLRP6 Inflammasome Recognizes Lipoteichoic Acid and Regulates Gram-Positive Pathogen Infection. Cell 2018, 175, 1651–1664.e14. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Chen, H.; Jiang, R.; Zhang, L.; Wang, L.; Gan, H.; Jiang, N.; Zhao, J.; Zhai, X.; Liang, P. NLRP6 contributes to inflammation and brain injury following intracerebral haemorrhage by activating autophagy. J. Mol. Med. 2020, 98, 1319–1331. [Google Scholar] [CrossRef]
- Flood, W.D.; Moyer, R.W.; Tsykin, A.; Sutherland, G.R.; Koblar, S.A. Nxf and Fbxo33: Novel seizure-responsive genes in mice. Eur. J. Neurosci. 2004, 20, 1819–1826. [Google Scholar] [CrossRef]
- Ooe, N.; Saito, K.; Mikami, N.; Nakatuka, I.; Kaneko, H. Identification of a novel basic helix-loop-helix-PAS factor, NXF, reveals a Sim2 competitive, positive regulatory role in dendritic-cytoskeleton modulator drebrin gene expression. Mol. Cell Biol. 2004, 24, 608–616. [Google Scholar] [CrossRef]
- Crews, S.T.; Fan, C.M. Remembrance of things PAS: Regulation of development by bHLH-PAS proteins. Curr. Opin. Genet. Dev. 1999, 9, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Kewley, R.J.; Whitelaw, M.L.; Chapman-Smith, A. The mammalian basic helix-loop-helix/PAS family of transcriptional regulators. Int. J. Biochem. Cell Biol. 2004, 36, 189–204. [Google Scholar] [CrossRef]
- Choy, F.C.; Klarić, T.S.; Koblar, S.A.; Lewis, M.D. The Role of the Neuroprotective Factor Npas4 in Cerebral Ischemia. Int. J. Mol. Sci. 2015, 16, 29011–29028. [Google Scholar] [CrossRef]
- Jaehne, E.J.; Klarić, T.S.; Koblar, S.A.; Baune, B.T.; Lewis, M.D. Effects of Npas4 deficiency on anxiety, depression-like, cognition and sociability behaviour. Behav. Brain Res. 2015, 281, 276–282. [Google Scholar] [CrossRef]
- Zhang, Z.; Fei, P.; Mu, J.; Li, W.; Song, J. Hippocampal expression of aryl hydrocarbon receptor nuclear translocator 2 and neuronal PAS domain protein 4 in a rat model of depression. Neurol. Sci. 2014, 35, 277–282. [Google Scholar] [CrossRef]
- Xie, J.; Wu, E.Q.; Zheng, Z.-J.; Croft, J.B.; Greenlund, K.J.; Mensah, G.A.; Labarthe, D.R. Impact of stroke on health-related quality of life in the noninstitutionalized population in the United States. Stroke 2006, 37, 2567–2572. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Preclinical and clinical research on inflammation after intracerebral hemorrhage. Prog. Neurobiol. 2010, 92, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, S.; Chang, J.; Wei, J.; Feng, M.; Wang, R. Perihematomal Edema After Intracerebral Hemorrhage: An Update on Pathogenesis, Risk Factors, and Therapeutic Advances. Front. Immunol. 2021, 12, 740632. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.H.; Chen, Q.X.; Cheng, B.L.; Fang, X.M. Defensins and sepsis. BioMed Res. Int. 2014, 2014, 180109. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.A.; Cookson, B.T. Salmonella induces macrophage death by caspase-1-dependent necrosis. Mol. Microbiol. 2000, 38, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Qian, J.; Zhang, P.; Li, H.; Shen, H.; Li, X.; Chen, G. Gasdermin D serves as a key executioner of pyroptosis in experimental cerebral ischemia and reperfusion model both in vivo and in vitro. J. Neurosci. Res. 2019, 97, 645–660. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, T.; Chen, X.-F.; Xu, Z.-X.; Cao, J.-B.; Sun, H. TMEM59 protects against cerebral ischemic stroke by suppressing pyroptosis and microglial activation. Biochem. Biophys. Res. Commun. 2021, 543, 72–79. [Google Scholar] [CrossRef]
- Lin, X.; Ye, H.; Siaw-Debrah, F.; Pan, S.; He, Z.; Ni, H.; Xu, Z.; Jin, K.; Zhuge, Q.; Huang, L. AC-YVAD-CMK Inhibits Pyroptosis and Improves Functional Outcome after Intracerebral Hemorrhage. BioMed Res. Int. 2018, 2018, 3706047. [Google Scholar] [CrossRef]
- Liu, W.; Liu, J.; Wang, W.; Wang, Y.; Ouyang, X. NLRP6 Induces Pyroptosis by Activation of Caspase-1 in Gingival Fibroblasts. J. Dent. Res. 2018, 97, 1391–1398. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.; Zhao, W.; Xiao, X.; Liu, S.; Peng, Q.; Jiang, N.; Zhou, B. NLRP6-Dependent Pyroptosis-Related lncRNAs Predict the Prognosis of Hepatocellular Carcinoma. Front. Med. 2022, 9, 760722. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, N.; Zhang, L.; Meng, C.; Zhao, J.; Wu, J. NLRP6 expressed in astrocytes aggravates neurons injury after OGD/R through activating the inflammasome and inducing pyroptosis. Int. Immunopharmacol. 2020, 80, 106183. [Google Scholar] [CrossRef] [PubMed]
- Chapman-Smith, A.; Whitelaw, M.L. Novel DNA binding by a basic helix-loop-helix protein. Role Dioxin Recept. PAS Domain. J. Biol. Chem. 2006, 281, 12535–12545. [Google Scholar] [CrossRef] [PubMed]
- Leong, W.K.; Klaric, T.S.; Lin, Y.; Lewis, M.D.; Koblar, S.A. Upregulation of the neuronal Per-Arnt-Sim domain protein 4 (Npas4) in the rat corticolimbic system following focal cerebral ischemia. Eur. J. Neurosci. 2013, 37, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Shamloo, M.; Soriano, L.; Von Schack, D.; Rickhag, M.; Chin, D.J.; Gonzalez-Zulueta, M.; Gidö, G.; Urfer, R.; Wieloch, T.; Nikolich, K. Npas4, a novel helix-loop-helix PAS domain protein, is regulated in response to cerebral ischemia. Eur. J. Neurosci. 2006, 24, 2705–2720. [Google Scholar] [CrossRef]
- Sun, X.; Lin, Y. Npas4: Linking Neuronal Activity to Memory. Trends Neurosci. 2016, 39, 264–275. [Google Scholar] [CrossRef]
- Choy, F.C.; Klarić, T.; Leong, W.K.; Koblar, S.; Lewis, M.D. Reduction of the neuroprotective transcription factor Npas4 results in increased neuronal necrosis, inflammation and brain lesion size following ischaemia. J. Cereb. Blood Flow. Metab. 2016, 36, 1449–1463. [Google Scholar] [CrossRef]
- Kim, S.; Park, D.; Kim, J.; Kim, D.; Kim, H.; Mori, T.; Jung, H.; Lee, D.; Hong, S.; Jeon, J.; et al. Npas4 regulates IQSEC3 expression in hippocampal somatostatin interneurons to mediate anxiety-like behavior. Cell Rep. 2021, 36, 109417. [Google Scholar] [CrossRef]
- Kersse, K.; Bertrand, M.J.; Lamkanfi, M.; Vandenabeele, P. NOD-like receptors and the innate immune system: Coping with danger, damage and death. Cytokine Growth Factor Rev. 2011, 22, 257–276. [Google Scholar] [CrossRef]
- Wang, C.; Ruan, J. Mechanistic Insights into Gasdermin Pore Formation and Regulation in Pyroptosis. J. Mol. Biol. 2022, 434, 167297. [Google Scholar] [CrossRef]
- Greb-Markiewicz, B.; Zarębski, M.; Ożyhar, A. Multiple sequences orchestrate subcellular trafficking of neuronal PAS domain-containing protein 4 (NPAS4). J. Biol. Chem. 2018, 293, 11255–11270. [Google Scholar] [CrossRef]
- Spiegel, I.; Mardinly, A.R.; Gabel, H.W.; Bazinet, J.E.; Couch, C.H.; Tzeng, C.P.; Harmin, D.A.; Greenberg, M.E. Npas4 regulates excitatory-inhibitory balance within neural circuits through cell-type-specific gene programs. Cell 2014, 157, 1216–1229. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Guo, O.; Zhen, Z.; Zhen, J. Essential Functions of the Transcription Factor Npas4 in Neural Circuit Development, Plasticity, and Diseases. Front. Neurosci. 2020, 14, 603373. [Google Scholar] [CrossRef] [PubMed]
- Reisz-Porszasz, S.; Probst, M.R.; Fukunaga, B.N.; Hankinson, O. Identification of functional domains of the aryl hydrocarbon receptor nuclear translocator protein (ARNT). Mol. Cell Biol. 1994, 14, 6075–6086. [Google Scholar]
- Fairman, R.; Beran-Steed, R.K.; Handel, T.M. Heteronuclear (1H, 13C, 15N) NMR assignments and secondary structure of the basic region-helix-loop-helix domain of E47. Protein Sci. A Publ. Protein Soc. 1997, 6, 175–184. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jian, D.; Qin, L.; Gan, H.; Zheng, S.; Xiao, H.; Duan, Y.; Zhang, M.; Liang, P.; Zhao, J.; Zhai, X. NPAS4 Exacerbates Pyroptosis via Transcriptionally Regulating NLRP6 in the Acute Phase of Intracerebral Hemorrhage in Mice. Int. J. Mol. Sci. 2023, 24, 8320. https://doi.org/10.3390/ijms24098320
Jian D, Qin L, Gan H, Zheng S, Xiao H, Duan Y, Zhang M, Liang P, Zhao J, Zhai X. NPAS4 Exacerbates Pyroptosis via Transcriptionally Regulating NLRP6 in the Acute Phase of Intracerebral Hemorrhage in Mice. International Journal of Molecular Sciences. 2023; 24(9):8320. https://doi.org/10.3390/ijms24098320
Chicago/Turabian StyleJian, Dan, Le Qin, Hui Gan, Shuyue Zheng, Han Xiao, Yuhao Duan, Mi Zhang, Ping Liang, Jing Zhao, and Xuan Zhai. 2023. "NPAS4 Exacerbates Pyroptosis via Transcriptionally Regulating NLRP6 in the Acute Phase of Intracerebral Hemorrhage in Mice" International Journal of Molecular Sciences 24, no. 9: 8320. https://doi.org/10.3390/ijms24098320