
Spatial Attention Modulates Neuronal Interactions between Simple and Complex Cells in V1
 ,
, 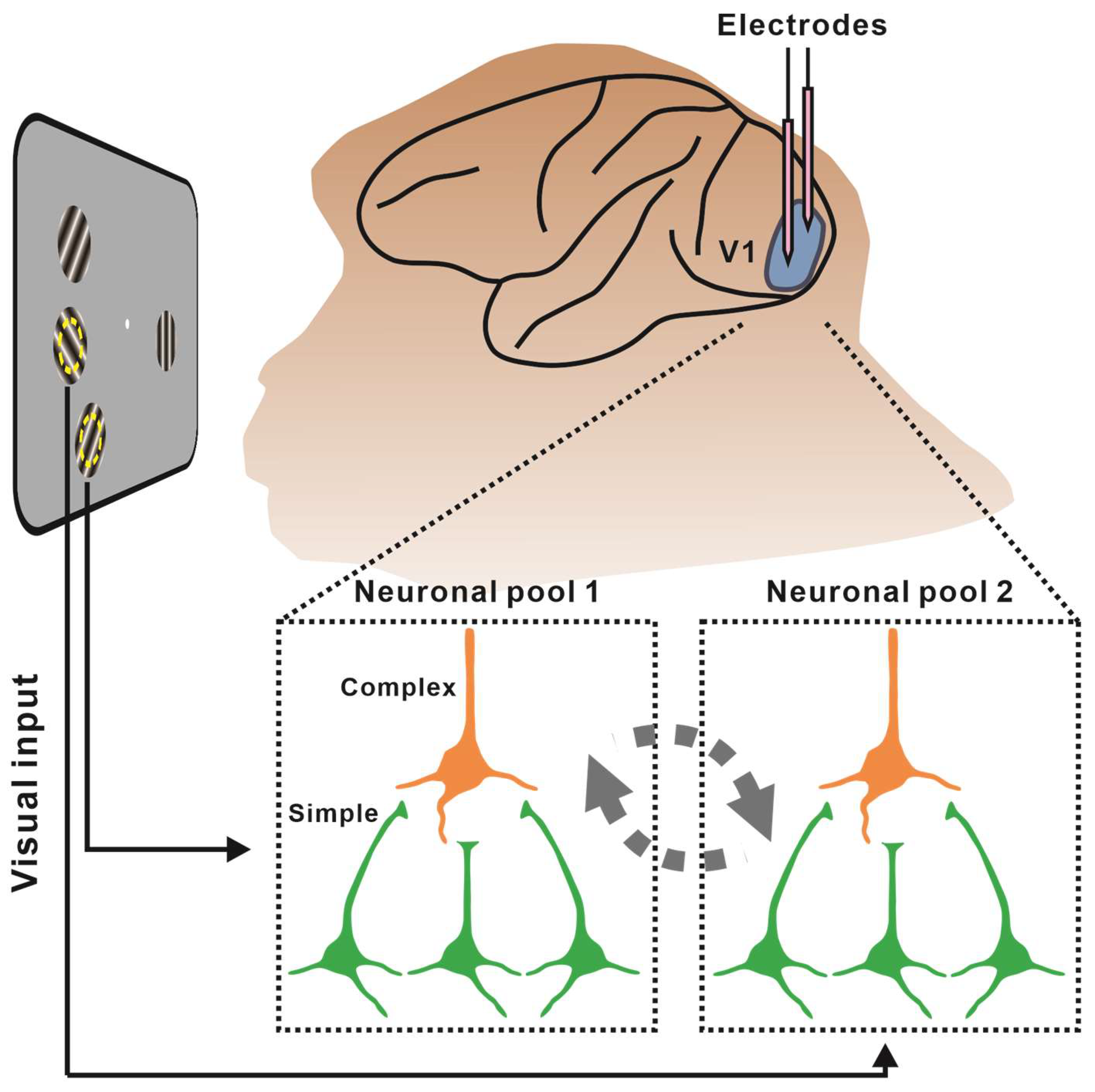{kind=link}
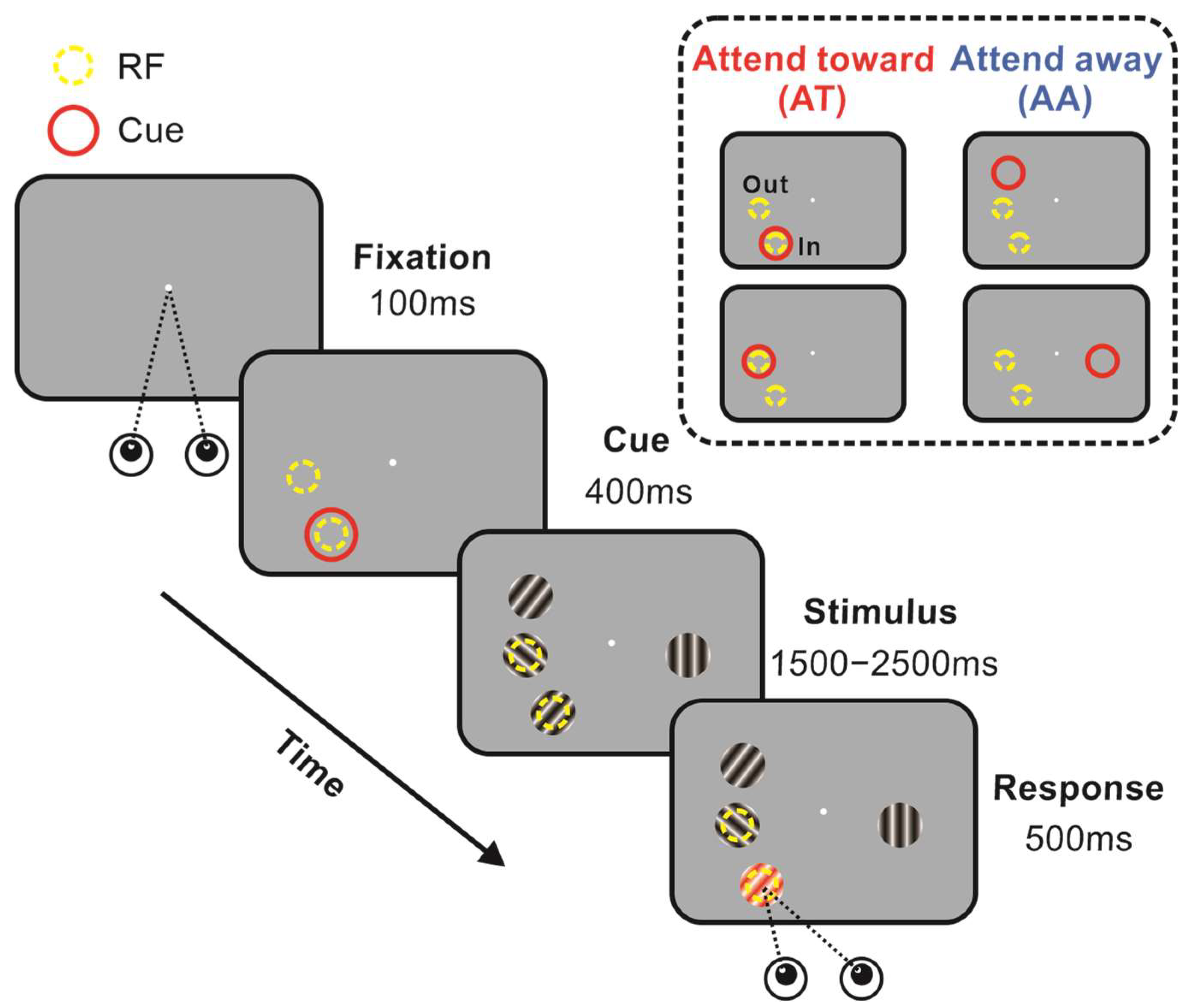{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
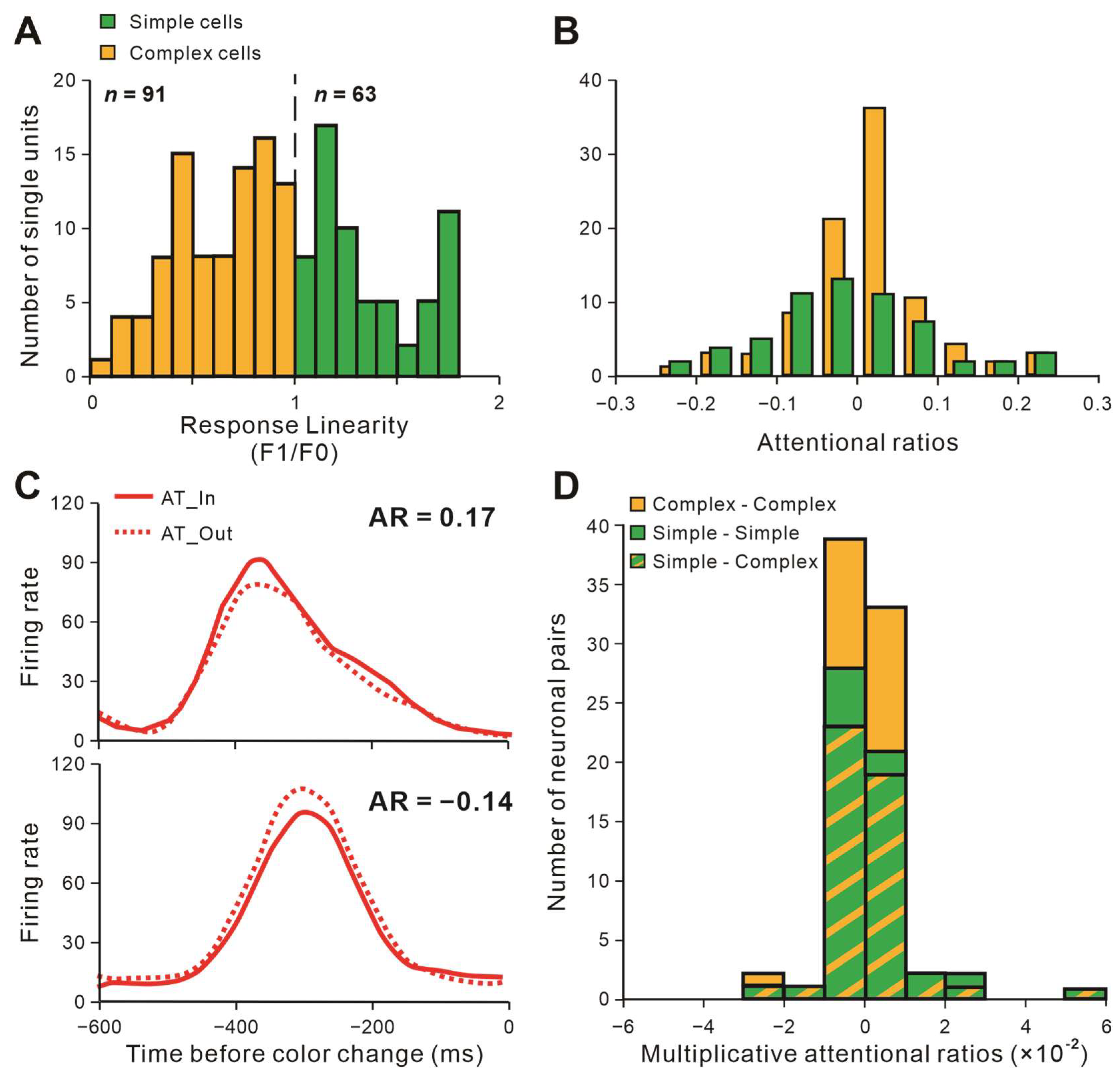
2.1. Neuronal Classification and Attentional Modulation on Firing Rates
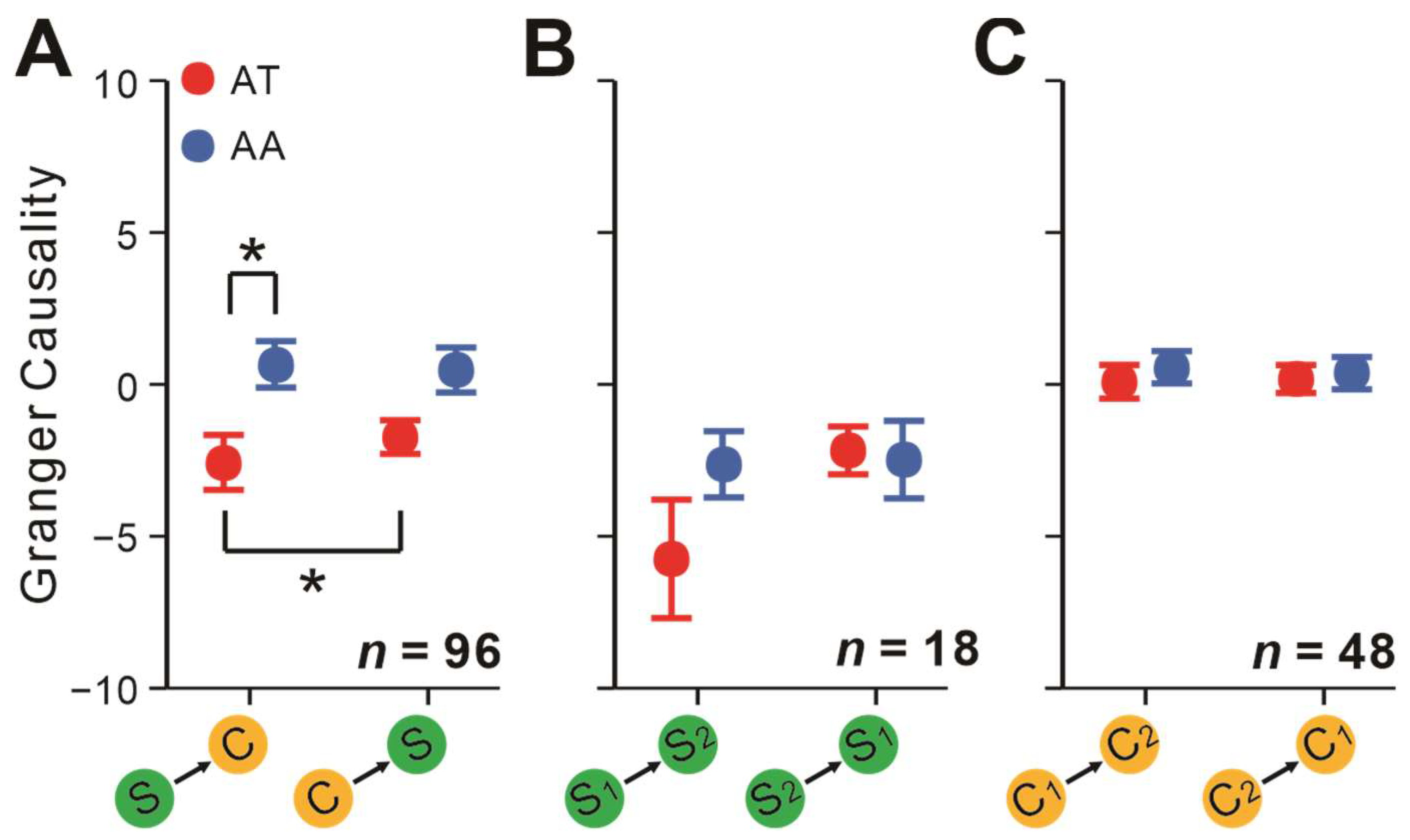
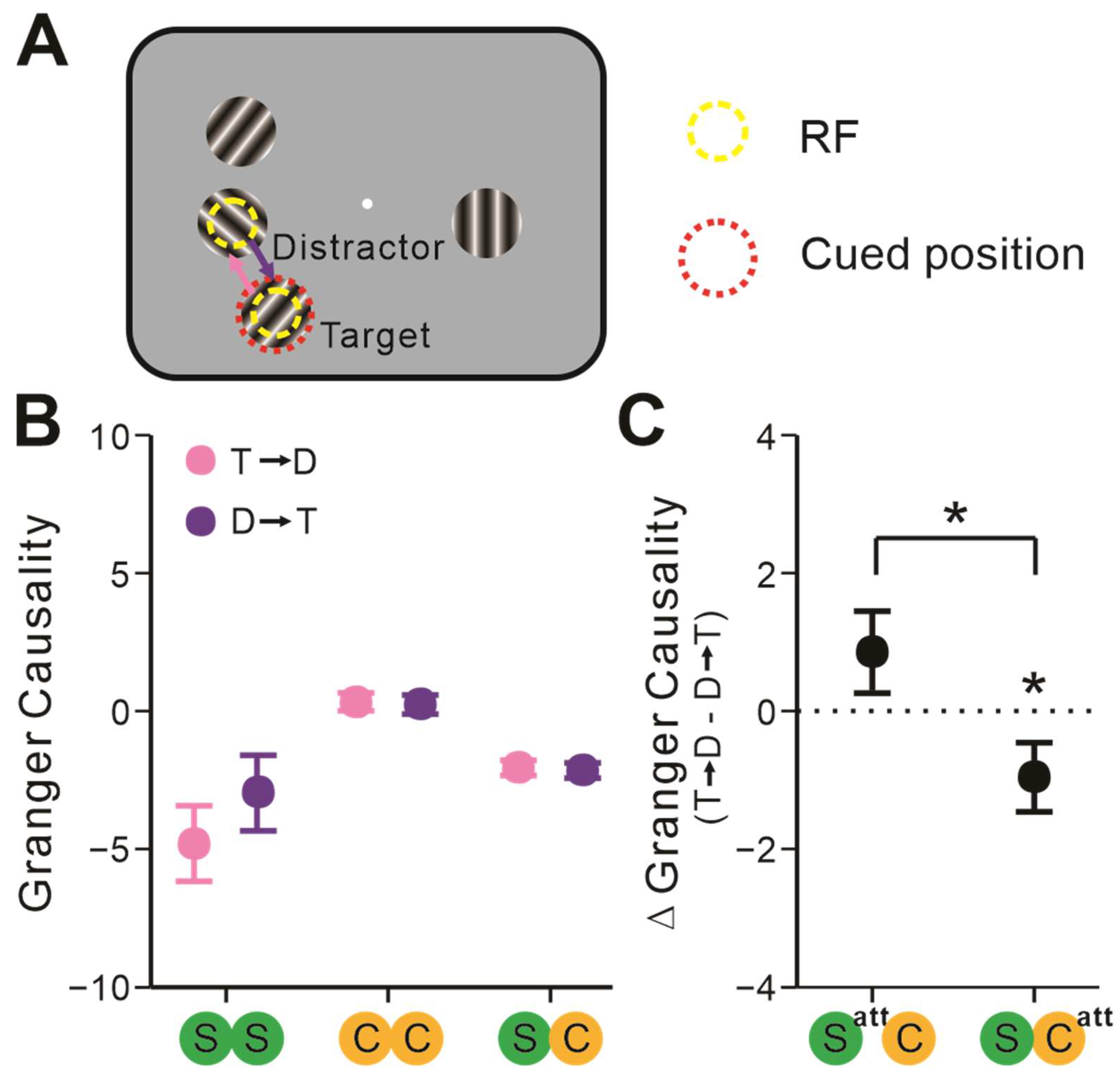
2.2. Attentional Modulation of Neuronal Interactions
2.3. The Attentional Focus Influenced S-C Pairs’ Interactions
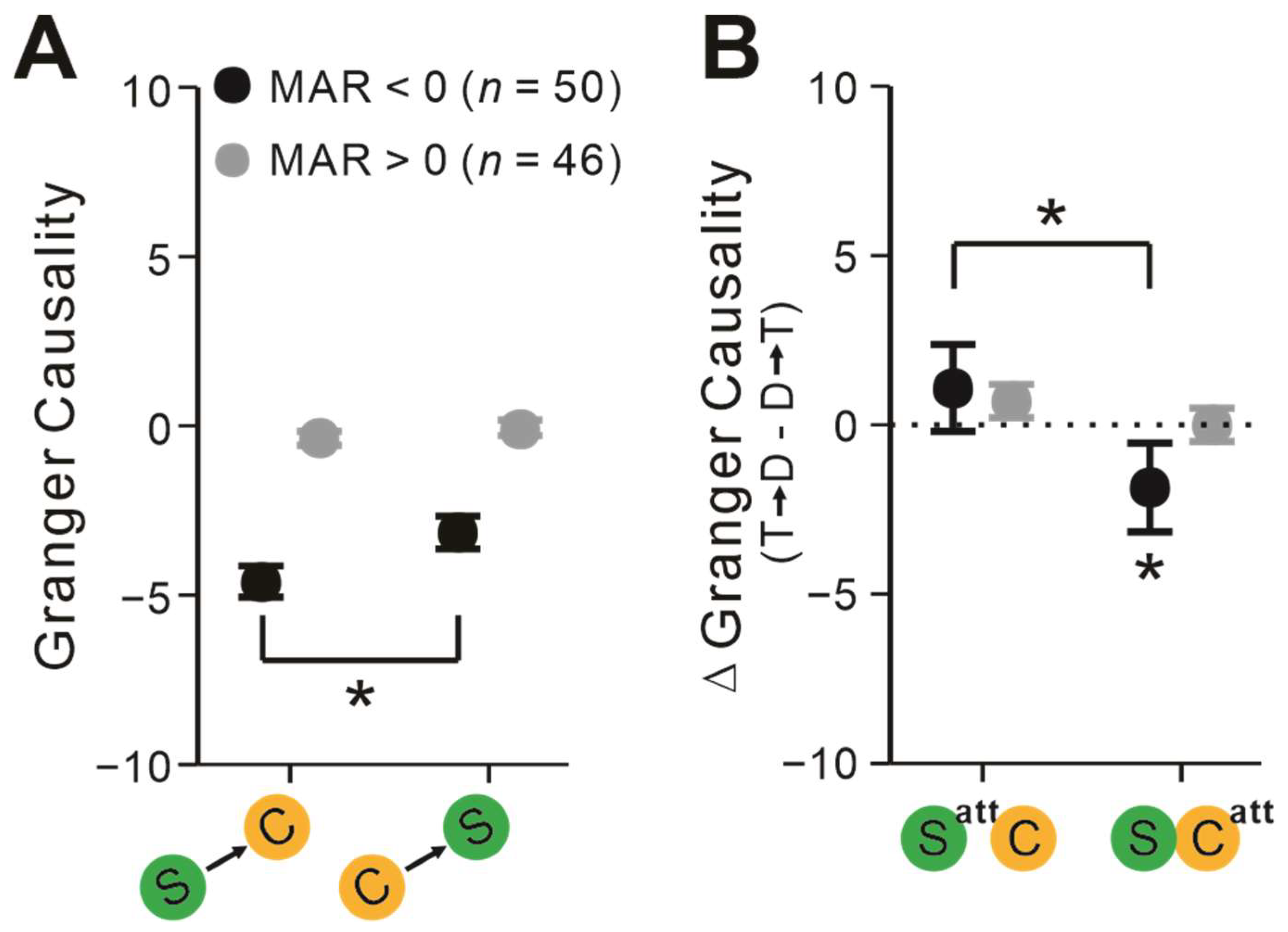
2.4. Pairs’ Interactions Depend on Attentional Modulation of Firing Rates
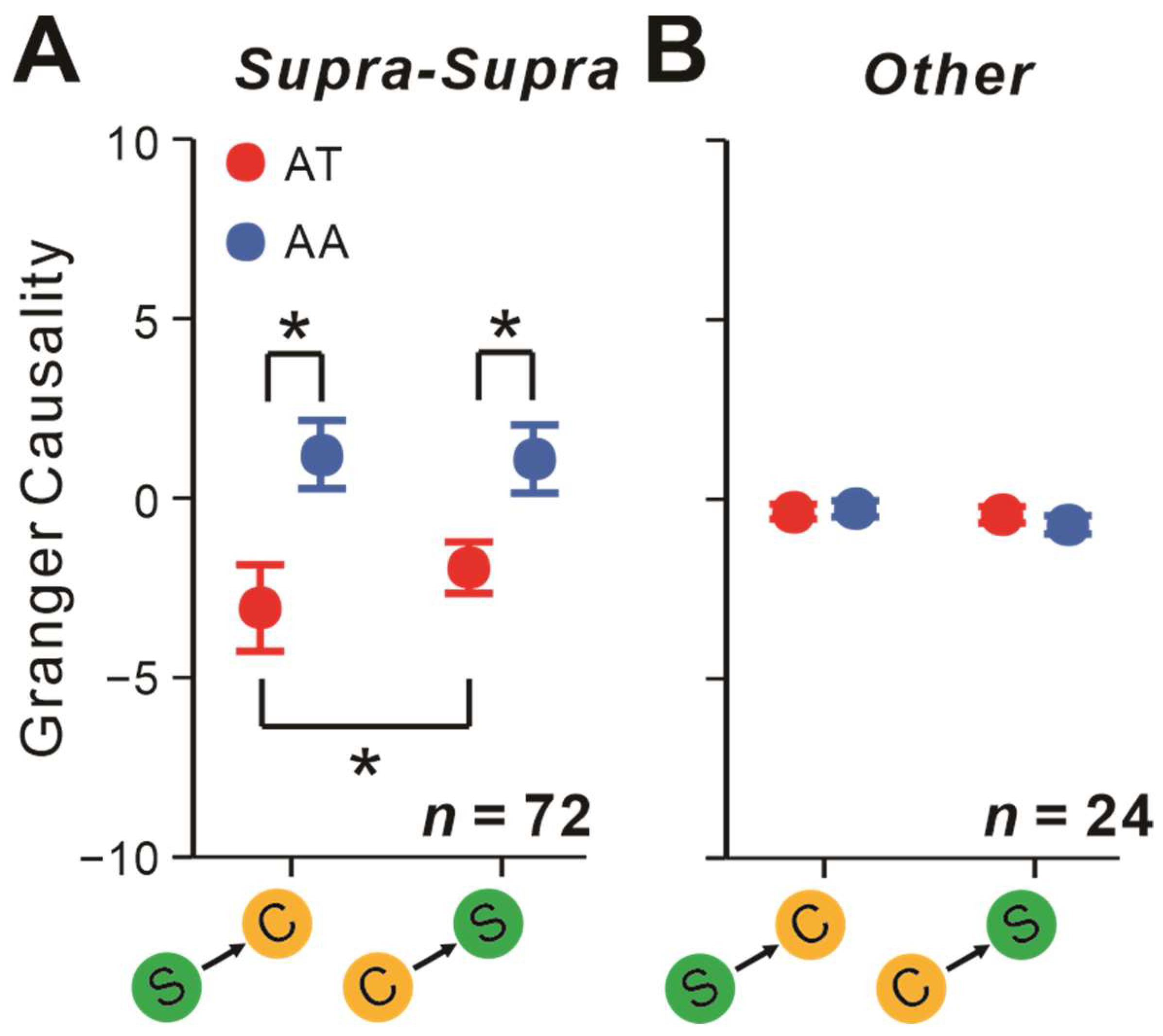
2.5. Attentional Impacts on Interactions Depending on the Layers Neurons Originated from
3. Discussion
3.1. Comparison to Previous Neuronal Communication Studies
3.2. The Influence of Attentional Focus Placement on S-C Pairs
3.3. S-C Pairs’ Communication Depends on Their MAR
3.4. The Layers Neurons Come from Explains Attentional Impact on S-C Pairs’ Interaction
3.5. Other Potential Confounders That Cannot Explain Our Findings
3.6. Limitations
4. Materials and Methods
4.1. Subjects
4.2. Spatial Attention Behavioral Task
4.3. Electrophysiological Recordings
4.4. Data Analysis
4.4.1. Electrophysiological Data Analysis and Classification of Neurons
4.4.2. Receptive Field Mapping
4.4.3. Granger Causality Analysis
4.4.4. Attentional Ratios and Multiplicative Attentional Ratios
4.4.5. Laminar Alignment
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maunsell, J.H. Neuronal mechanisms of visual attention. Annu. Rev. Vis. Sci. 2015, 1, 373–391. [Google Scholar] [CrossRef]
- Moore, T.; Zirnsak, M. Neural mechanisms of selective visual attention. Annu. Rev. Psychol. 2017, 68, 47–72. [Google Scholar] [CrossRef] [PubMed]
- Briggs, F.; Mangun, G.R.; Usrey, W.M. Attention enhances synaptic efficacy and the signal-to-noise ratio in neural circuits. Nature 2013, 499, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Bosman, C.; Schoffelen, J.M.; Brunet, N.; Oostenveld, R.; Bastos, A.; Womelsdorf, T.; Rubehn, B.; Stieglitz, T.; Deweerd, P.; Fries, P. Attentional stimulus selection through selective synchronization between monkey visual areas. Neuron 2012, 75, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Ferro, D.; van Kempen, J.; Boyd, M.; Panzeri, S.; Thiele, A. Directed information exchange between cortical layers in macaque V1 and V4 and its modulation by selective attention. Proc. Natl. Acad. Sci. USA 2021, 118, e2022097118. [Google Scholar] [CrossRef] [PubMed]
- Gregoriou, G.G.; Gotts, S.J.; Zhou, H.; Desimone, R. High-frequency, long-range coupling between prefrontal and visual cortex during attention. Science 2009, 324, 1207–1210. [Google Scholar] [CrossRef]
- Ruff, D.A.; Cohen, M.R. Attention increases spike count correlations between visual cortical areas. J. Neurosci. 2016, 36, 7523–7534. [Google Scholar] [CrossRef]
- Srinath, R.; Ruff, D.A.; Cohen, M.R. Attention improves information flow between neuronal populations without changing the communication subspace. Curr. Biol. 2021, 31, 5299–5313. [Google Scholar] [CrossRef]
- Hubel, D.H.; Wiesel, T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol. 1962, 160, 106–154. [Google Scholar] [CrossRef]
- Martinez, L.M.; Wang, Q.; Reid, R.C.; Pillai, C.; Alonso, J.-M.; Sommer, F.T.; Hirsch, J.A. Receptive field structure varies with layer in the primary visual cortex. Nat. Neurosci. 2005, 8, 372–379. [Google Scholar] [CrossRef]
- Antolik, J.; Bednar, J. Development of maps of simple and complex cells in the primary visual cortex. Front. Comput. Neurosci. 2011, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Jang, J.; Paik, S.B. Periodic clustering of simple and complex cells in visual cortex. Neural Netw. 2021, 143, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ferster, D. Functional Coupling from Simple to Complex Cells in the Visually Driven Cortical Circuit. J. Neurosci. 2013, 33, 18855–18866. [Google Scholar] [CrossRef]
- Hembrook-Short, J.R.; Mock, V.L.; Usrey, W.M.; Briggs, F. Attention enhances the efficacy of communication in V1 local circuits. J. Neurosci. 2019, 39, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Granger, C.W.J. Investigating causal relations by econometric models and cross-spectral methods. Econometrica 1969, 37, 424–438. [Google Scholar] [CrossRef]
- Brovelli, A.; Ding, M.; Ledberg, A.; Chen, Y.; Nakamura, R.; Bressler, S.L. Beta oscillations in a large-scale sensorimotor cortical network: Directional influences revealed by Granger causality. Proc. Natl. Acad. Sci. USA 2004, 101, 9849–9854. [Google Scholar] [CrossRef]
- Kim, S.; Putrino, D.; Ghosh, S.; Brown, E.N. A Granger causality measure for point process models of ensemble neural spiking activity. PLoS Comput. Biol. 2011, 7, e1001110. [Google Scholar] [CrossRef]
- Liang, H.; Gong, X.; Chen, M.; Yan, Y.; Li, W.; Gilbert, C.D. Interactions between feedback and lateral connections in the primary visual cortex. Proc. Natl. Acad. Sci. USA 2017, 114, 8637–8642. [Google Scholar] [CrossRef]
- Schmitt, L.I.; Wimmer, R.D.; Nakajima, M.; Happ, M.; Mofakham, S.; Halassa, M.M. Thalamic amplification of cortical connectivity sustains attentional control. Nature 2017, 545, 219–223. [Google Scholar] [CrossRef]
- Hu, Q.; Hu, W.; Liu, K.; Bu, X.; Hu, L.; Li, L.; Chai, X.; Chen, Y. Modulation of spike count correlations between macaque primary visual cortex neurons by difficulty of attentional task. Neurosci. Bull. 2021, 38, 489–504. [Google Scholar] [CrossRef]
- Hu, Q.; Zheng, Z.; Sui, X.; Li, L.; Chai, X.; Chen, Y. Spatial Attention Modulates Spike Count Correlations and Granger Causality in the Primary Visual Cortex. Front. Cell. Neurosci. 2022, 16, 838049. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.H.; Chelazzi, L.; Desimone, R. Competitive mechanisms subserve attention in macaque areas V2 and V4. J. Neurosci. 1999, 19, 1736–1753. [Google Scholar] [CrossRef] [PubMed]
- Buschman, T.J.; Kastner, S. From behavior to neural dynamics: An integrated theory of attention. Neuron 2015, 88, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Esghaei, M.; Daliri, M.R.; Treue, S. Attention Decreases Phase-Amplitude Coupling, Enhancing Stimulus Discriminability in Cortical Area MT. Front. Neural Circuits 2015, 9, 82. [Google Scholar] [CrossRef]
- Esghaei, M.; Daliri, M.R.; Treue, S. Attention decouples action potentials from the phase of local field potentials in macaque visual cortical area MT. BMC Biol. 2018, 16, 86. [Google Scholar] [CrossRef]
- Zareian, B.; Maboudi, K.; Daliri, M.R.; Abrishami Moghaddam, H.; Treue, S.; Esghaei, M. Attention strengthens across-trial pre-stimulus phase coherence in visual cortex, enhancing stimulus processing. Sci. Rep. 2020, 10, 4837. [Google Scholar] [CrossRef]
- Motter, B.C. Focal attention produces spatially selective processing in visual cortical areas V1, V2, and V4 in the presence of competing stimuli. J. Neurophysiol. 1993, 70, 909–919. [Google Scholar] [CrossRef]
- Luck, S.J.; Chelazzi, L.; Hillyard, S.A.; Desimone, R. Neural mechanisms of spatial selective attention in areas V1, V2, and V4 of macaque visual cortex. J. Neurophysiol. 1997, 77, 24–42. [Google Scholar] [CrossRef]
- Buffalo, E.A.; Fries, P.; Landman, R.; Liang, H.; Desimone, R. A backward progression of attentional effects in the ventral stream. Proc. Natl. Acad. Sci. USA 2010, 107, 361–365. [Google Scholar] [CrossRef]
- Zhou, H.; Desimone, R. Feature-based attention in the frontal eye field and area V4 during visual search. Neuron 2011, 70, 1205–1217. [Google Scholar] [CrossRef]
- Denfield, G.H.; Ecker, A.S.; Shinn, T.J.; Bethge, M.; Tolias, A.S. Attentional fluctuations induce shared variability in macaque primary visual cortex. Nat. Commun. 2018, 9, 2654. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.S.; Lund, R.D.; Hendrickson, A.E.; Bunt, A.H.; Fuchs, A.F. The origin of efferent pathways from the primary visual cortex, area 17, of the macaque monkey as shown by retrograde transport of horseradish peroxidase. J. Comp. Neurol 1975, 164, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E.M. Local circuits in primary visual cortex of the macaque monkey. Annu. Rev. Neurosci. 1998, 21, 47–74. [Google Scholar] [CrossRef] [PubMed]
- Douglas, R.J.; Martin, K.A. Neuronal circuits of the neocortex. Annu. Rev. Neurosci. 2004, 27, 419–451. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, J.A.; Gallagher, C.A.; Alonso, J.M.; Martinez, L.M. Ascending projections of simple and complex cells in layer 6 of the cat striate cortex. J. Neurosci. 1998, 18, 8086–8094. [Google Scholar] [CrossRef]
- Martinez, L.M.; Alonso, J.M.; Reid, R.C.; Hirsch, J.A. Laminar processing of stimulus orientation in cat visual cortex. J. Physiol. 2002, 540, 321–333. [Google Scholar] [CrossRef]
- Alonso, J.M.; Martinez, L.M. Functional connectivity between simple cells and complex cells in cat striate cortex. Nat. Neurosci. 1998, 1, 395–403. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Academic Press: New York, NY, USA, 2013. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Ringach, D.L.; Shapley, R.M.; Hawken, M.J. Orientation selectivity in macaque V1: Diversity and laminar dependence. J. Neurosci. 2002, 22, 5639–5651. [Google Scholar] [CrossRef]
- Nandy, A.S.; Nassi, J.J.; Reynolds, J.H. Laminar Organization of Attentional Modulation in Macaque Visual Area V4. Neuron 2017, 93, 235–246. [Google Scholar] [CrossRef]
- Chen, Y.; Martinez-Conde, S.; Macknik, S.L.; Bereshpolova, Y.; Swadlow, H.A.; Alonso, J.-M. Task difficulty modulates the activity of specific neuronal populations in primary visual cortex. Nat. Neurosci. 2008, 11, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, K.A.; Mitchell, J.F.; Reynolds, J.H. Spatial attention modulates center-surround interactions in macaque visual area v4. Neuron 2009, 61, 952–963. [Google Scholar] [CrossRef]
- Awh, E.; Matsukura, M.; Serences, J.T. Top-down control over biased competition during covert spatial orienting. J. Exp. Psychol. Hum. Percept. Perform. 2003, 29, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Serences, J.T.; Yantis, S.; Culberson, A.; Awh, E. Preparatory activity in visual cortex indexes distractor suppression during covert spatial orienting. J. Neurophysiol. 2004, 92, 3538–3545. [Google Scholar] [CrossRef]
- Bonifazi, P.; Goldin, M.; Picardo, M.A.; Jorquera, I.; Cattani, A.; Bianconi, G.; Represa, A.; Ben-Ari, Y.; Cossart, R. GABAergic hub neurons orchestrate synchrony in developing hippocampal networks. Science 2009, 326, 1419–1424. [Google Scholar] [CrossRef]
- Kwan, A.C.; Dan, Y. Dissection of cortical microcircuits by single-neuron stimulation in vivo. Curr. Biol. 2012, 22, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Womelsdorf, T.; Ardid, S.; Everling, S.; Valiante, T.A. Burst firing synchronizes prefrontal and anterior cingulate cortex during attentional control. Curr. Biol. 2014, 24, 2613–2621. [Google Scholar] [CrossRef]
- Buffalo, E.A.; Fries, P.; Landman, R.; Buschman, T.J.; Desimone, R. Laminar differences in gamma and alpha coherence in the ventral stream. Proc. Natl. Acad. Sci. USA 2011, 108, 11262–11267. [Google Scholar] [CrossRef]
- Barone, P.; Batardiere, A.; Knoblauch, K.; Kennedy, H. Laminar distribution of neurons in extrastriate areas projecting to visual areas V1 and V4 correlates with the hierarchical rank and indicates the operation of a distance rule. J. Neurosci. 2000, 20, 3263–3281. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Ignashchenkova, A.; Thier, P.; Hafed, Z.M. Neuronal response gain enhancement prior to microsaccades. Curr. Biol. 2015, 25, 2065–2074. [Google Scholar] [CrossRef]
- McFarland, J.M.; Cumming, B.G.; Butts, D.A. Variability and correlations in primary visual cortical neurons driven by fixational eye movements. J. Neurosci. 2016, 36, 6225–6241. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.H.; Gaska, J.P.; Nagler, M.; Pollen, D.A. Spatial and temporal frequency selectivity of neurones in visual cortical areas V1 and V2 of the macaque monkey. J. Physiol. 1985, 365, 331–363. [Google Scholar] [CrossRef]
- Hembrook-Short, J.R.; Mock, V.L.; Briggs, F. Attentional modulation of neuronal activity depends on neuronal feature selectivity. Curr. Biol. 2017, 27, 1878–1887 e1875. [Google Scholar] [CrossRef]
- Skottun, B.C.; De Valois, R.L.; Grosof, D.H.; Movshon, J.A.; Albrecht, D.G.; Bonds, A. Classifying simple and complex cells on the basis of response modulation. Vis. Res. 1991, 31, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Ringach, D.; Shapley, R. Reverse correlation in neurophysiology. Cogn. Sci. 2004, 28, 147–166. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Z.; Hu, Q.; Bu, X.; Jiang, H.; Sui, X.; Li, L.; Chai, X.; Chen, Y. Spatial Attention Modulates Neuronal Interactions between Simple and Complex Cells in V1. Int. J. Mol. Sci. 2023, 24, 8229. https://doi.org/10.3390/ijms24098229
Zheng Z, Hu Q, Bu X, Jiang H, Sui X, Li L, Chai X, Chen Y. Spatial Attention Modulates Neuronal Interactions between Simple and Complex Cells in V1. International Journal of Molecular Sciences. 2023; 24(9):8229. https://doi.org/10.3390/ijms24098229
Chicago/Turabian StyleZheng, Zhiyan, Qiyi Hu, Xiangdong Bu, Hongru Jiang, Xiaohong Sui, Liming Li, Xinyu Chai, and Yao Chen. 2023. "Spatial Attention Modulates Neuronal Interactions between Simple and Complex Cells in V1" International Journal of Molecular Sciences 24, no. 9: 8229. https://doi.org/10.3390/ijms24098229