
Is the In Vitro Observed NETosis the Favored Physiological Death of Neutrophils or Mainly Induced by an Isolation Bias?

Abstract
:1. Introduction
2. Results
2.1. Chemotactic Migration
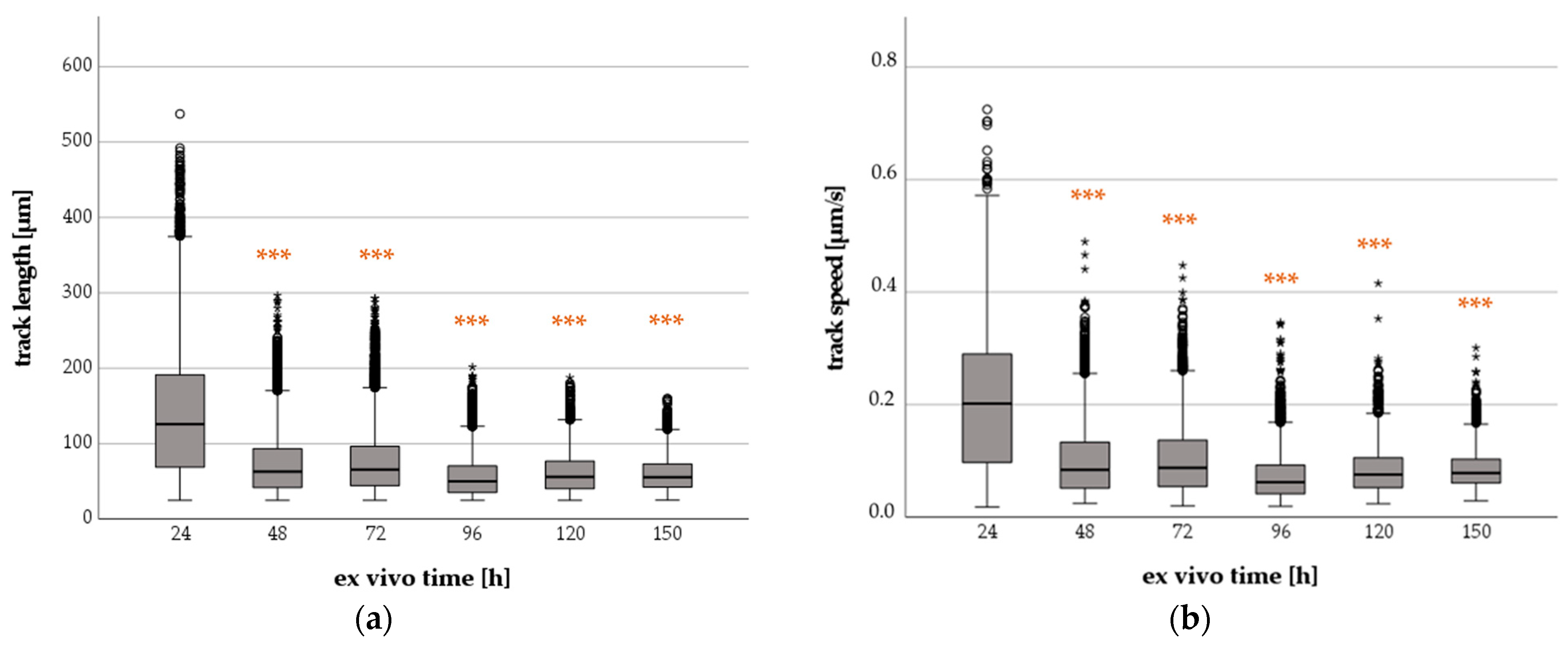
2.1.1. Effect of Increasing Ex Vivo Time on Neutrophil TL and TS
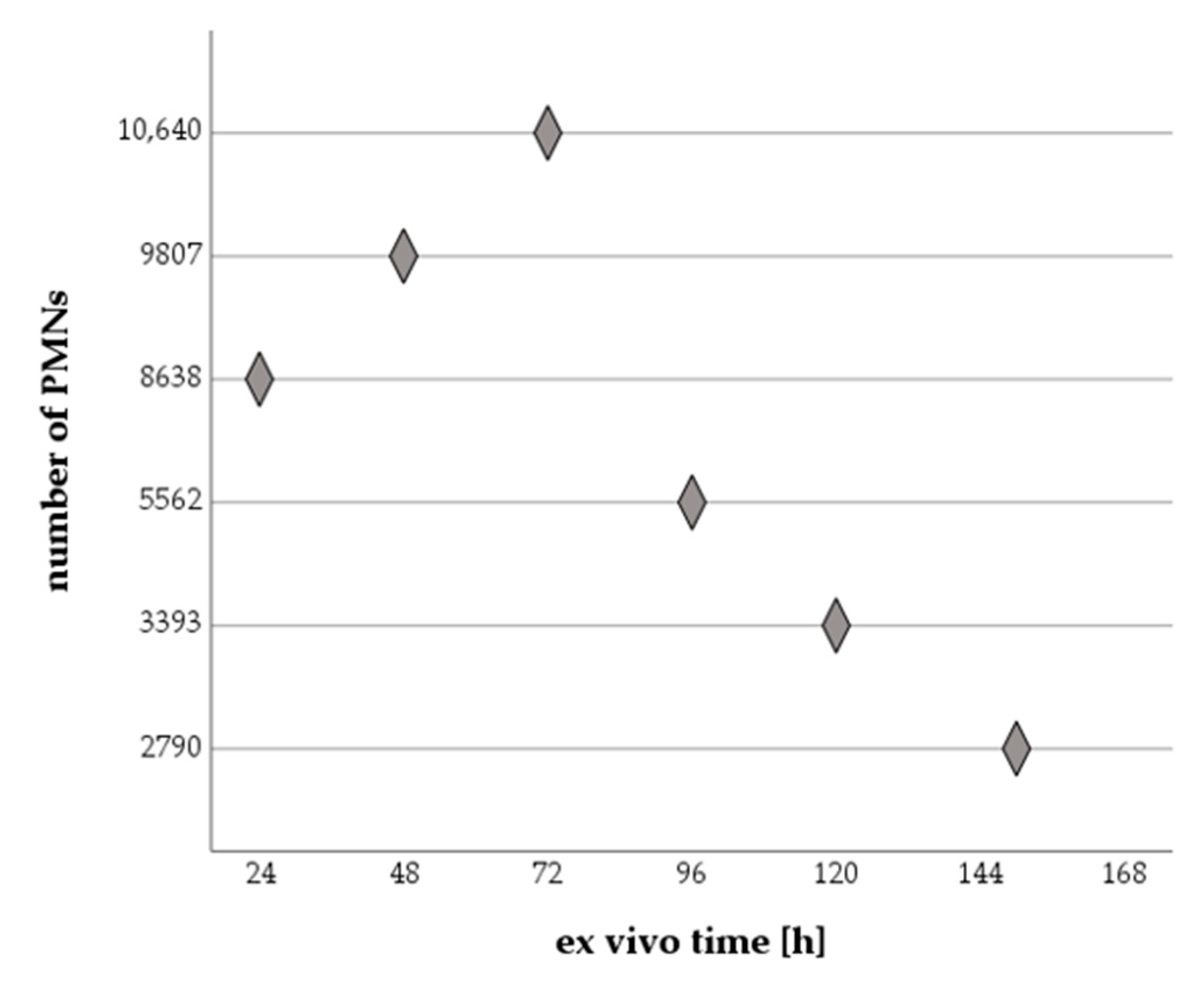
2.1.2. Effect of Increasing Ex Vivo Time on Analyzed PMN Numbers
2.2. Analysis of Fluorescence Staining of Neutrophils
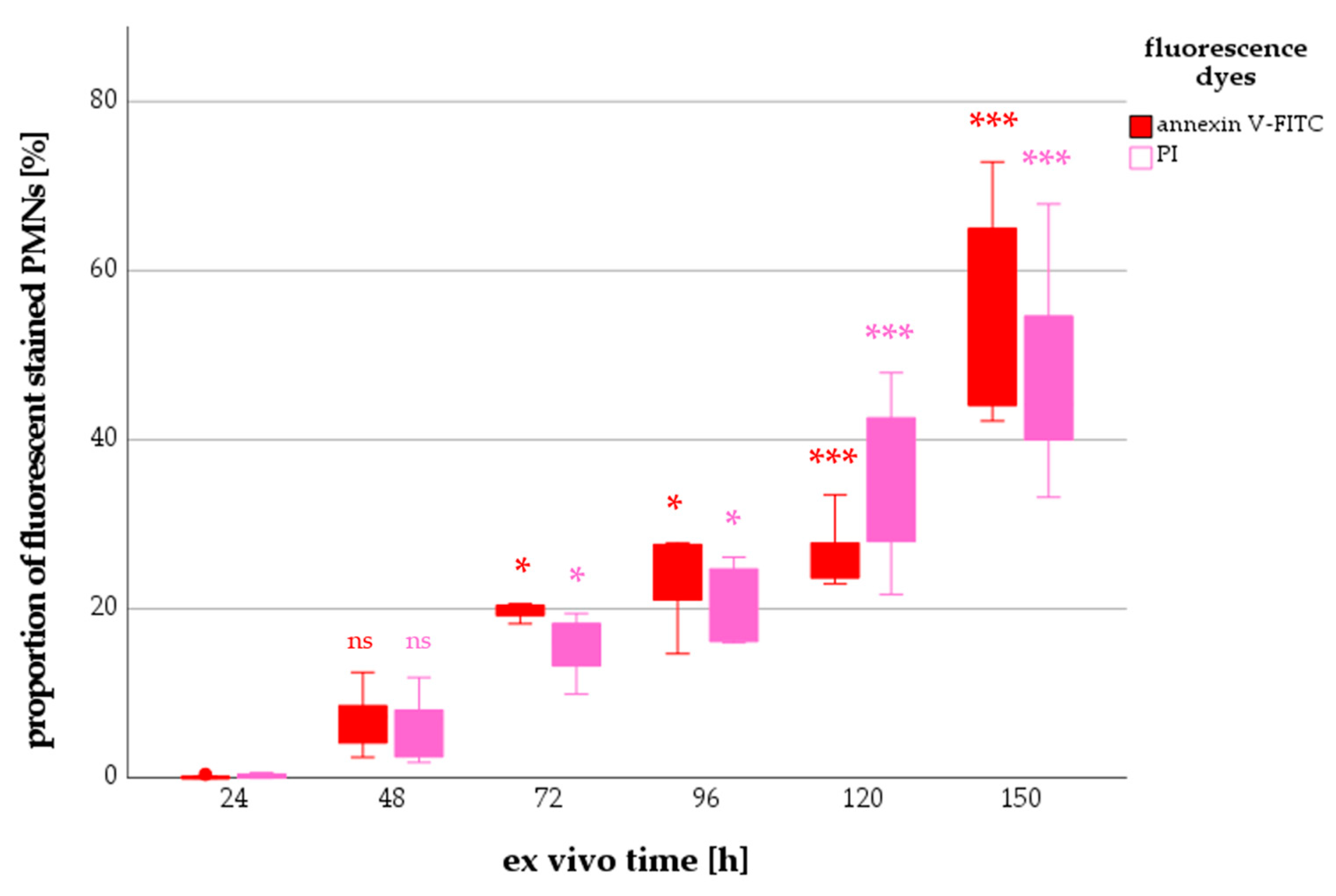
2.2.1. Annexin V and PI Staining of Neutrophils
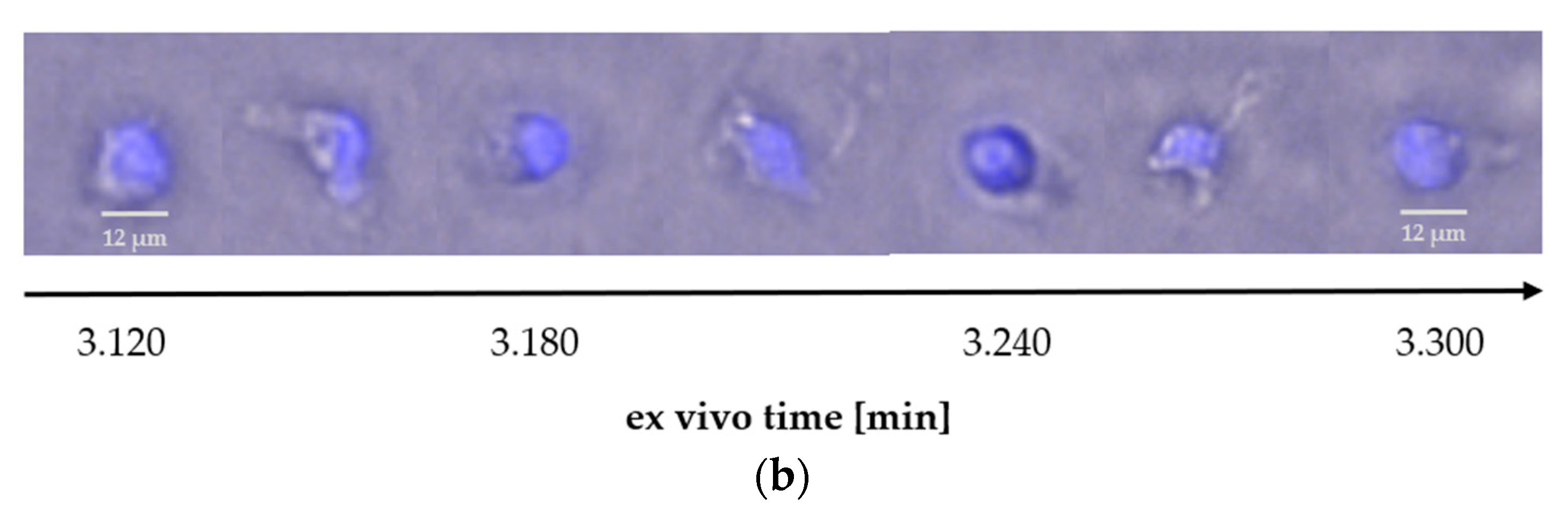
2.2.2. Comparison of DAPI-Stained PMNs from Different Isolation Methods
2.2.3. Progress of Median ROS Intensity
3. Discussion
4. Materials and Methods
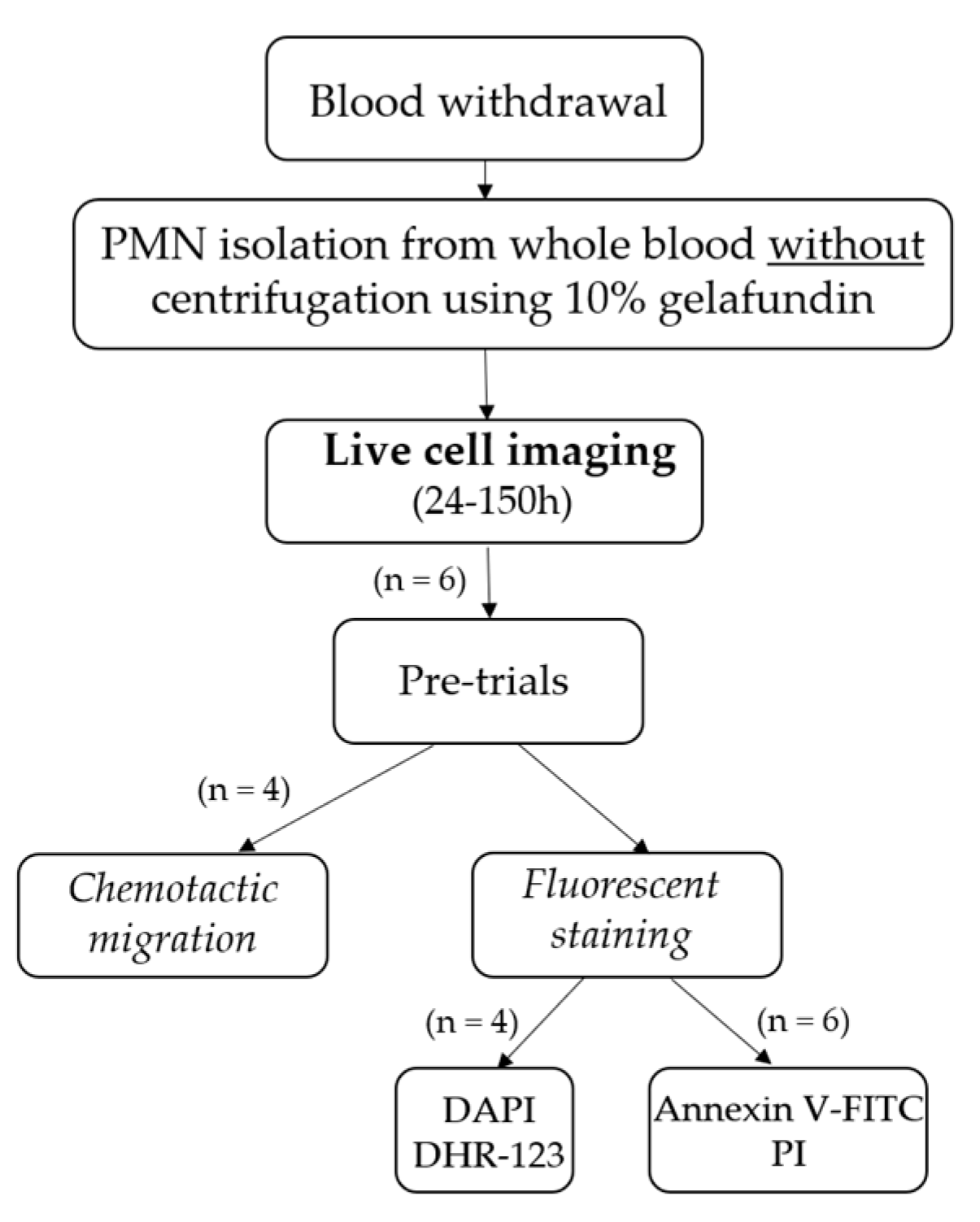
4.1. Study Plan
4.2. Blood Withdrawal and PMN Isolation
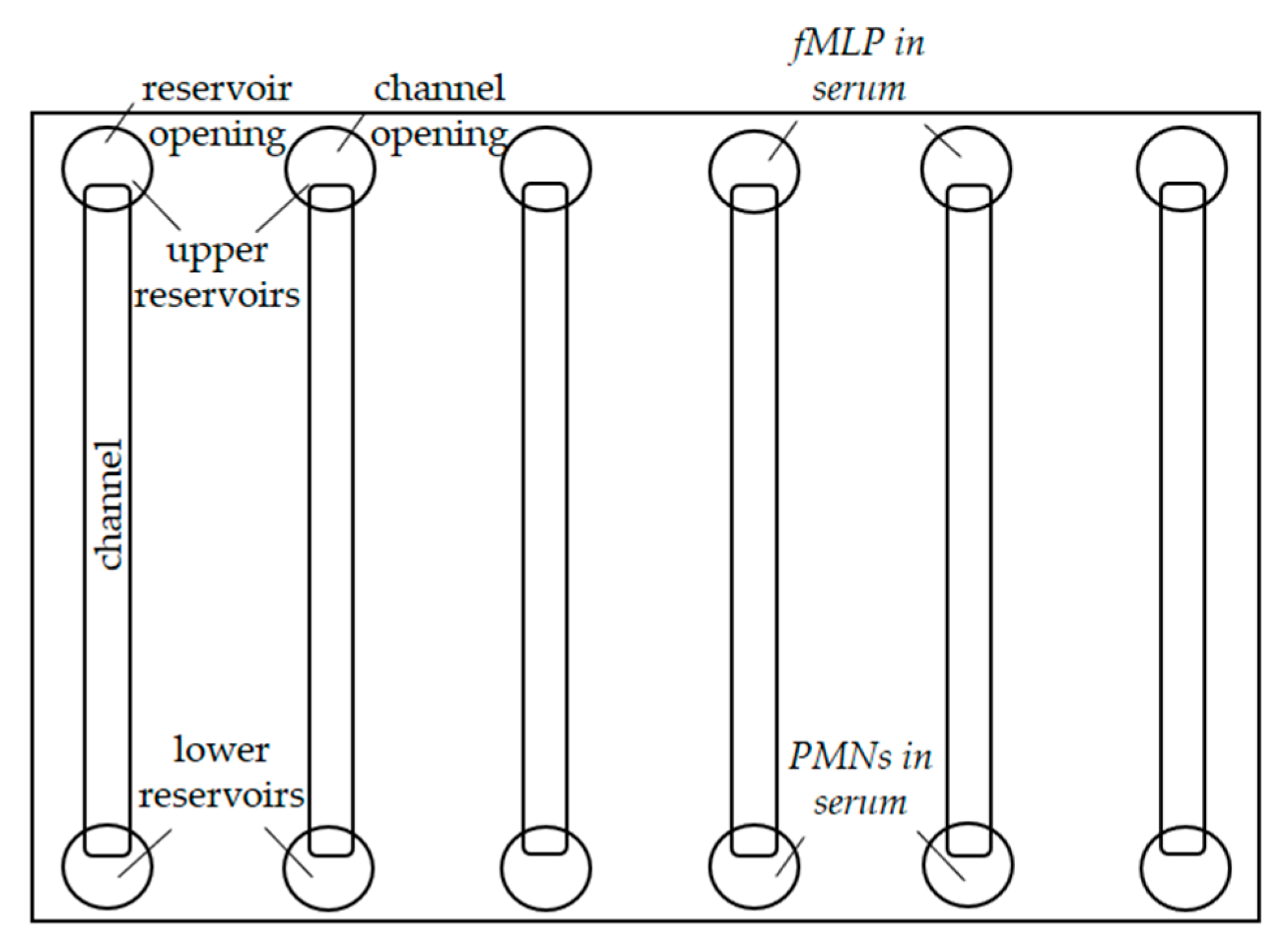
4.3. Experimental Setup
4.4. Data Analysis
4.4.1. Migration Analysis
4.4.2. Fluorescent Staining
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef]
- Denning, N.-L.; Aziz, M.; Gurien, S.D.; Wang, P. DAMPs and NETs in Sepsis. Front. Immunol. 2019, 10, 2536. [Google Scholar] [CrossRef]
- Bashant, K.R.; Aponte, A.M.; Randazzo, D.; Rezvan Sangsari, P.; Wood, A.J.; Bibby, J.A.; West, E.E.; Vassallo, A.; Manna, Z.G.; Playford, M.P.; et al. Proteomic, biomechanical and functional analyses define neutrophil hetero-geneity in systemic lupus erythematosus. Ann. Rheum. Dis. 2021, 80, 209–218. [Google Scholar] [CrossRef]
- Apel, F.; Zychlinsky, A.; Kenny, E.F. The role of neutrophil extracellular traps in rheumatic diseases. Nat. Rev. Rheumatol. 2018, 14, 467–475. [Google Scholar] [CrossRef]
- Gray, R.D.; Hardisty, G.; Regan, K.H.; Smith, M.; Robb, C.T.; Duffin, R.; Mackellar, A.; Felton, J.M.; Paemka, L.; McCullagh, B.N.; et al. Delayed neutrophil apoptosis enhances NET formation in cystic fibrosis. Thorax 2018, 73, 134–144. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef]
- Adrover, J.M.; Del Fresno, C.; Crainiciuc, G.; Cuartero, M.I.; Casanova-Acebes, M.; Weiss, L.A.; Huerga-Encabo, H.; Silvestre-Roig, C.; Rossaint, J.; Cossío, I.; et al. A Neutrophil Timer Coordinates Immune Defense and Vas-cular Protection. Immunity 2019, 50, 390–402. [Google Scholar] [CrossRef]
- Pillay, J.; Den Braber, I.; Vrisekoop, N.; Kwast, L.M.; de Boer, R.J.; Borghans, J.A.M.; Tesselaar, K.; Koenderman, L. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Blood 2010, 116, 625–627. [Google Scholar] [CrossRef]
- Tofts, P.S.; Chevassut, T.; Cutajar, M.; Dowell, N.G.; Peters, A.M. Doubts concerning the recently reported hu-man neutrophil lifespan of 5.4 days. Blood 2011, 117, 6050–6052. [Google Scholar] [CrossRef]
- Lahoz-Beneytez, J.; Elemans, M.; Zhang, Y.; Ahmed, R.; Salam, A.; Block, M.; Niederalt, C.; Asquith, B.; Macallan, D. Human neutrophil kinetics: Modeling of stable isotope labeling data supports short blood neutrophil half-lives. Blood 2016, 127, 3431–3438. [Google Scholar] [CrossRef]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12, 631821. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chan, D.W.; Zaal, K.J.; Kaplan, M.J. A High-Throughput Real-Time Imaging Technique To Quantify NETosis and Distinguish Mechanisms of Cell Death in Human Neutrophils. J. Immunol. 2018, 200, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Mutua, V.; Gershwin, L.J. A Review of Neutrophil Extracellular Traps (NETs) in Disease: Potential Anti-NETs Therapeutics. Clin. Rev. Allergy Immunol. 2021, 61, 194–211. [Google Scholar] [CrossRef]
- Pai, D.; Gruber, M.; Pfaehler, S.-M.; Bredthauer, A.; Lehle, K.; Trabold, B. Polymorphonuclear Cell Chemotaxis and Suicidal NETosis: Simultaneous Observation Using fMLP, PMA, H7, and Live Cell Imaging. J. Immunol. Res. 2020, 2020, 1415947. [Google Scholar] [CrossRef]
- Bredthauer, A.; Kopfmueller, M.; Gruber, M.; Pfaehler, S.-M.; Lehle, K.; Petermichl, W.; Seyfried, T.; Bitzinger, D.; Redel, A. Therapeutic Anticoagulation with Argatroban and Heparins Reduces Granulocyte Migration: Possible Impact on ECLS-Therapy? Cardiovasc. Ther. 2020, 2020, 9783630. [Google Scholar] [CrossRef]
- Bredthauer, A.; Geiger, A.; Gruber, M.; Pfaehler, S.-M.; Petermichl, W.; Bitzinger, D.; Metterlein, T.; Seyfried, T. Propofol Ameliorates Exaggerated Human Neutrophil Activation in a LPS Sepsis Model. J. Inflamm. Res. 2021, 14, 3849–3862. [Google Scholar] [CrossRef]
- Segawa, K.; Kurata, S.; Yanagihashi, Y.; Brummelkamp, T.R.; Matsuda, F.; Nagata, S. Caspase-mediated cleavage of phospholipid flippase for apoptotic phosphatidylserine exposure. Science 2014, 344, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087288. [Google Scholar] [CrossRef]
- Crowley, L.C.; Scott, A.P.; Marfell, B.J.; Boughaba, J.A.; Chojnowski, G.; Waterhouse, N.J. Measuring Cell Death by Propidium Iodide Uptake and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087163. [Google Scholar] [CrossRef] [PubMed]
- Scannell, M.; Flanagan, M.B.; deStefani, A.; Wynne, K.J.; Cagney, G.; Godson, C.; Maderna, P. Annexin-1 and Peptide Derivatives Are Released by Apoptotic Cells and Stimulate Phagocytosis of Apoptotic Neutrophils by Macrophages. J. Immunol. 2007, 178, 4595–4605. [Google Scholar] [CrossRef]
- Li, B.; Zeng, M.; Zheng, H.; Huang, C.; He, W.; Lu, G.; Li, X.; Chen, Y.; Xie, R. Effects of ghrelin on the apoptosis of human neutrophils in vitro. Int. J. Mol. Med. 2016, 38, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Sulowska, Z.; Majewska, E.; Klink, M.; Banasik, M.; Tchórzewski, H. Flow Cytometric Evaluation of Human Neutrophil Apoptosis During Nitric Oxide Generation In Vitro: The Role of Exogenous Antioxidants. Mediat. Inflamm. 2005, 2005, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Zahler, S.; Kowalski, C.; Brosig, A.; Kupatt, C.; Becker, B.F.; Gerlach, E. The function of neutrophils isolated by a magnetic antibody cell separation technique is not altered in comparison to a density gradient centrifugation method. J. Immunol. Methods 1997, 200, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Swierczak, A.; Pollard, J.W. FACS isolation and analysis of human circulating and tumor neutrophils. Meth. Enzymol. 2020, 632, 229–257. [Google Scholar] [CrossRef]
- Hundhammer, T.; Gruber, M.; Wittmann, S. Paralytic Impact of Centrifugation on Human Neutrophils. Biomedicines 2022, 10, 2896. [Google Scholar] [CrossRef]
- Kraus, R.F.; Gruber, M.A. Neutrophils-From Bone Marrow to First-Line Defense of the Innate Immune System. Front. Immunol. 2021, 12, 767175. [Google Scholar] [CrossRef]
- Doblinger, N.; Bredthauer, A.; Mohrez, M.; Hähnel, V.; Graf, B.; Gruber, M.; Ahrens, N. Impact of hydroxyethyl starch and modified fluid gelatin on granulocyte phenotype and function. Transfusion 2019, 59, 2121–2130. [Google Scholar] [CrossRef]
- Koo, S.; Offner, R.; Haile, S.-M.; Brosig, A.; Hähnel, V.; Gruber, M.; Burkhardt, R.; Ahrens, N. Granulocyte con-centrate splitting does not affect phenotype and function. Transfusion 2023, 63, 393–401. [Google Scholar] [CrossRef]
- Keating, F.K.; Butenas, S.; Fung, M.K.; Schneider, D.J. Platelet-white blood cell (WBC) interaction, WBC apop-tosis, and procoagulant activity in stored red blood cells. Transfusion 2011, 51, 1086–1095. [Google Scholar] [CrossRef]
- Patel, A.A.; Zhang, Y.; Fullerton, J.N.; Boelen, L.; Rongvaux, A.; Maini, A.A.; Bigley, V.; Flavell, R.A.; Gilroy, D.W.; Asquith, B.; et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J. Exp. Med. 2017, 214, 1913–1923. [Google Scholar] [CrossRef]
- Pfirschke, C.; Engblom, C.; Gungabeesoon, J.; Lin, Y.; Rickelt, S.; Zilionis, R.; Messemaker, M.; Siwicki, M.; Ger-hard, G.M.; Kohl, A.; et al. Tumor-Promoting Ly-6G+ SiglecFhigh Cells Are Mature and Long-Lived Neutrophils. Cell Rep. 2020, 32, 108164. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Yan, J.; Liu, Z. Methodological advances in necroptosis research: From challenges to solutions. J. Natl. Cancer Cent. 2022, 2, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Sawai, H.; Domae, N. Discrimination between primary necrosis and apoptosis by necrostatin-1 in Annexin V-positive/propidium iodide-negative cells. Biochem. Biophys. Res. Commun. 2011, 411, 569–573. [Google Scholar] [CrossRef]
- Gardai, S.J.; McPhillips, K.A.; Frasch, S.C.; Janssen, W.J.; Starefeldt, A.; Murphy-Ullrich, J.E.; Bratton, D.L.; Oldenborg, P.-A.; Michalak, M.; Henson, P.M. Cell-Surface Careticulin Initiates Clearance of Viable or Apoptotic Cell through trans-Activation of LRP on the Phagocyte. Cell 2005, 123, 321–334. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Values (Range) |
|---|---|
| Number of Subjects | 6 |
| Gender (female/male) | 4/2 |
| Median Age (years) | 21 (20–58) |
| Ex Vivo Time [h] | Mean of Median ROS Intensity [afu] |
|---|---|
| 24 | 13.8 ± 1.9 |
| 48 | 13.1 ± 1.0 |
| 72 | 14.2 ± 3.2 |
| 96 | 14.1 ± 2.1 |
| 120 | 12.9 ± 0.8 |
| 150 | 13.8 ± 0.9 |
| Samples | Time [h] | TL [µm] (15 min) | TL [µm] (30 min) | Apoptotic [%] | Necrotic/Dead [%] |
|---|---|---|---|---|---|
| GC | 24 | 57 (calculated) | 114 | 0.42 | 9.84 |
| <1 kgs | 24 | 126 | 252 (calculated) | <0.5 | <0.5 |
| GC | 48 | 31.7 (calculated) | 63.5 | 0.79 | 17.4 |
| <1 kgs | 48 | 63 | 126 (calculated) | 7.2 | 7.0 |
| Cell Type (Human) | Mean Lifespan (Days) | Test Method |
|---|---|---|
| Monocytes (classical) | 1.0 ± 0.26 | deuterium-labeled glucose |
| Monocytes (intermediate) | 4.3 ± 0.36 | deuterium-labeled glucose |
| Monocytes (nonclassical) | 7.4 ± 0.53 | deuterium-labeled glucose |
| Cell State | Annexin V | PI |
|---|---|---|
| Vital | − | − |
| Apoptotic | + | − |
| Late apoptotic/necroptotic/necrotic | + | + |
| Dead | − | + |
| Cell State | Annexin V | PI |
|---|---|---|
| Vital | − | − |
| Early apoptotic | + | − |
| Apoptotic | + | + |
| Late apoptotic | + (weak) | + |
| Late apoptotic/necrotic | − | + |
| Upper Reservoir (60 µL) | Channel (5 µL) | Lower Reservoir (60 µL) | |
|---|---|---|---|
| fMLP [10 µM] in autologous serum | Cell medium (33%) DAPI [2 µg/mL] DHR-123 [6.6 µM] Collagen type I (50%) | Cell medium (38%) Annexin V-FITC [7.6 µg/mL] PI [4.6 µg/mL] Collagen type I (50%) | PMNs supernatant and autologous serum (each 50%) |
| Parameter (Abbreviation) | Unit | Description |
|---|---|---|
| Track length (TL) | [µm] | Total track of a PMN within 15 min |
| Mean track speed (TS) | [µm/s] | Migration speed of the tracked PMN |
| Number of PMNs | 1 | Total number of analyzed PMNs meeting inclusion criteria: TL ≥ 25 µm + track duration ≥ 450 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rimboeck, J.; Gruber, M.; Wittmann, S. Is the In Vitro Observed NETosis the Favored Physiological Death of Neutrophils or Mainly Induced by an Isolation Bias? Int. J. Mol. Sci. 2023, 24, 7368. https://doi.org/10.3390/ijms24087368
Rimboeck J, Gruber M, Wittmann S. Is the In Vitro Observed NETosis the Favored Physiological Death of Neutrophils or Mainly Induced by an Isolation Bias? International Journal of Molecular Sciences. 2023; 24(8):7368. https://doi.org/10.3390/ijms24087368
Chicago/Turabian StyleRimboeck, Julia, Michael Gruber, and Sigrid Wittmann. 2023. "Is the In Vitro Observed NETosis the Favored Physiological Death of Neutrophils or Mainly Induced by an Isolation Bias?" International Journal of Molecular Sciences 24, no. 8: 7368. https://doi.org/10.3390/ijms24087368