
Critical Role of the Presynaptic Protein CAST in Maintaining the Photoreceptor Ribbon Synapse Triad
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
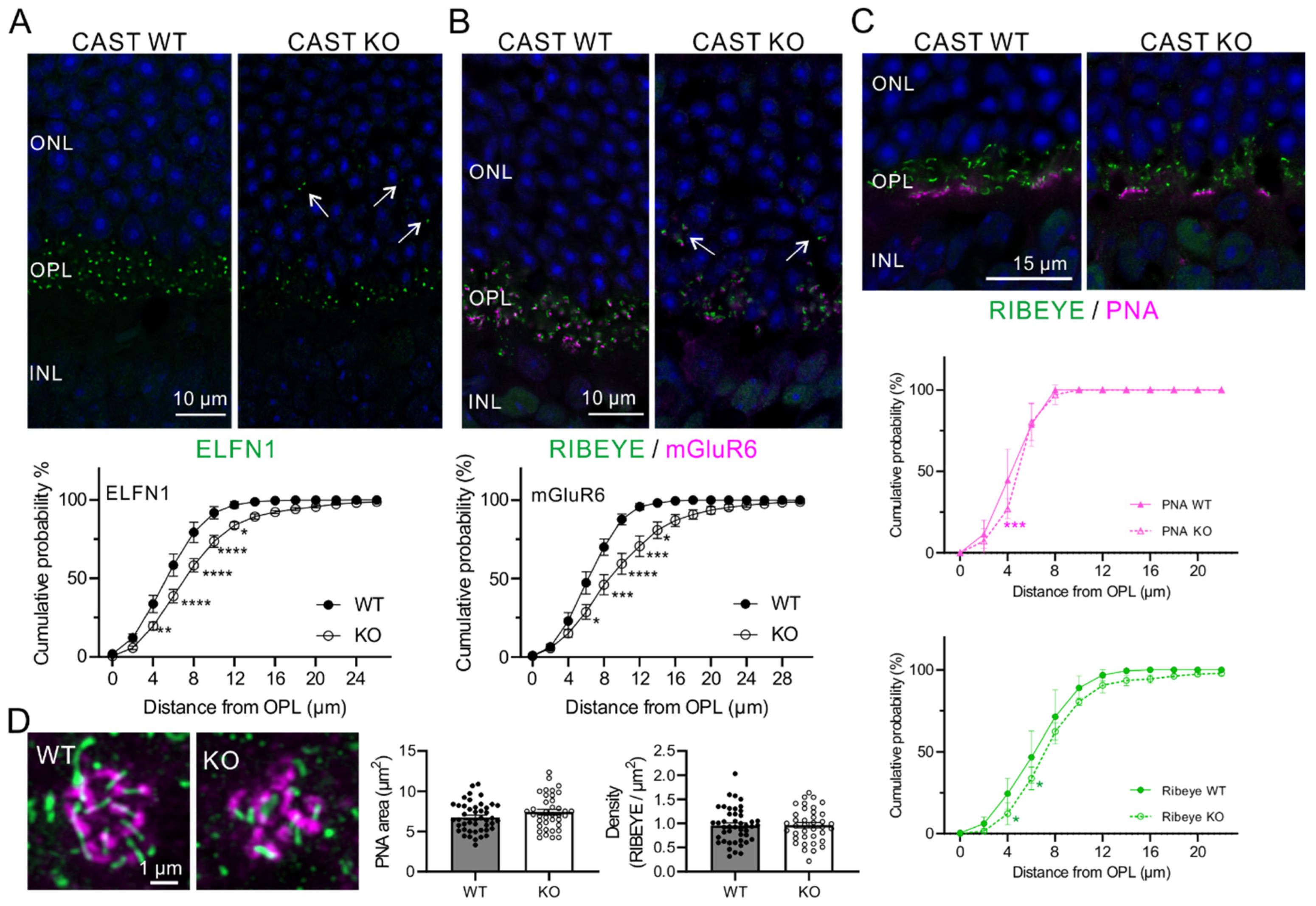
2.1. Ectopic Localization of Rod Terminals in the CAST KO Retina
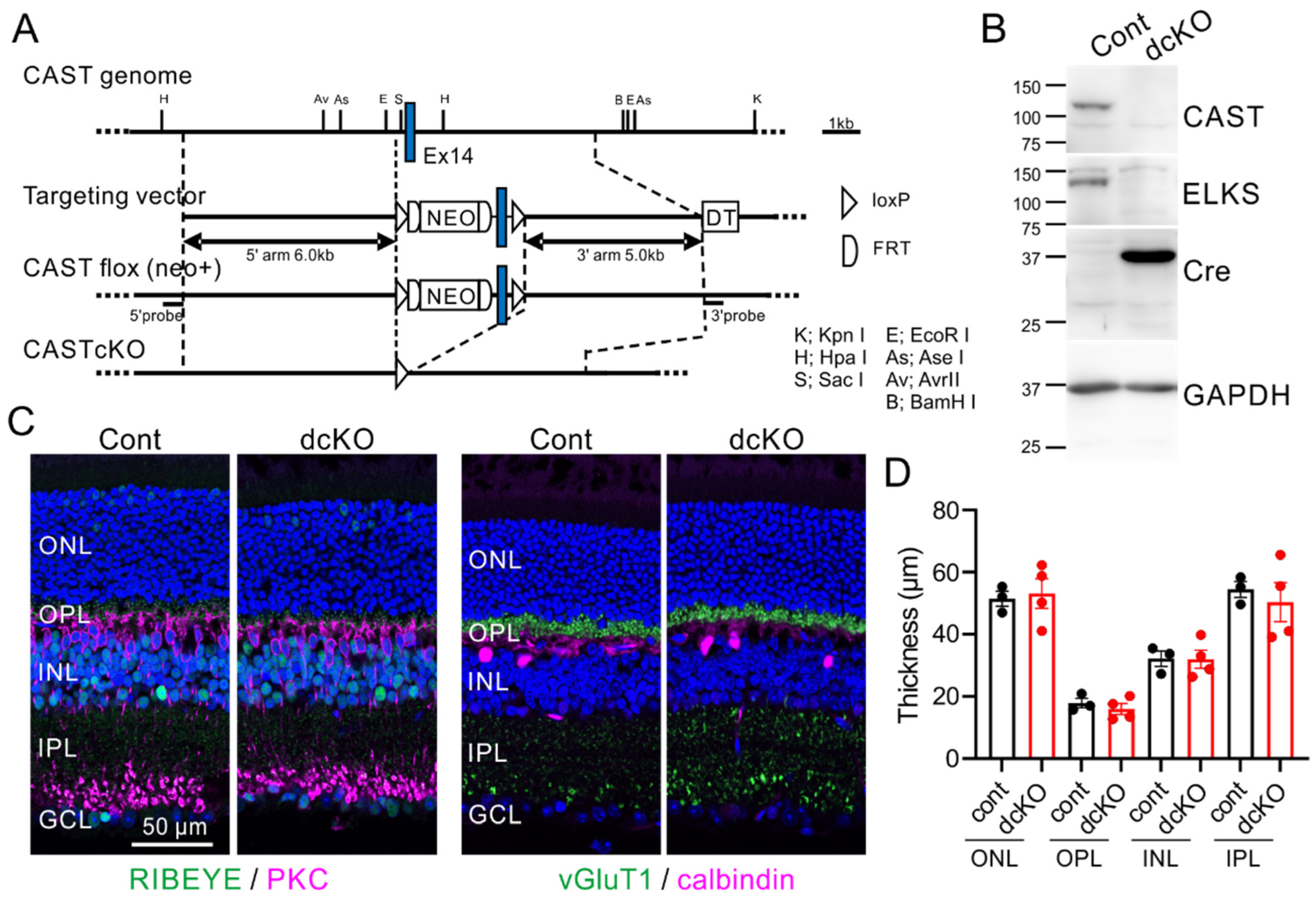
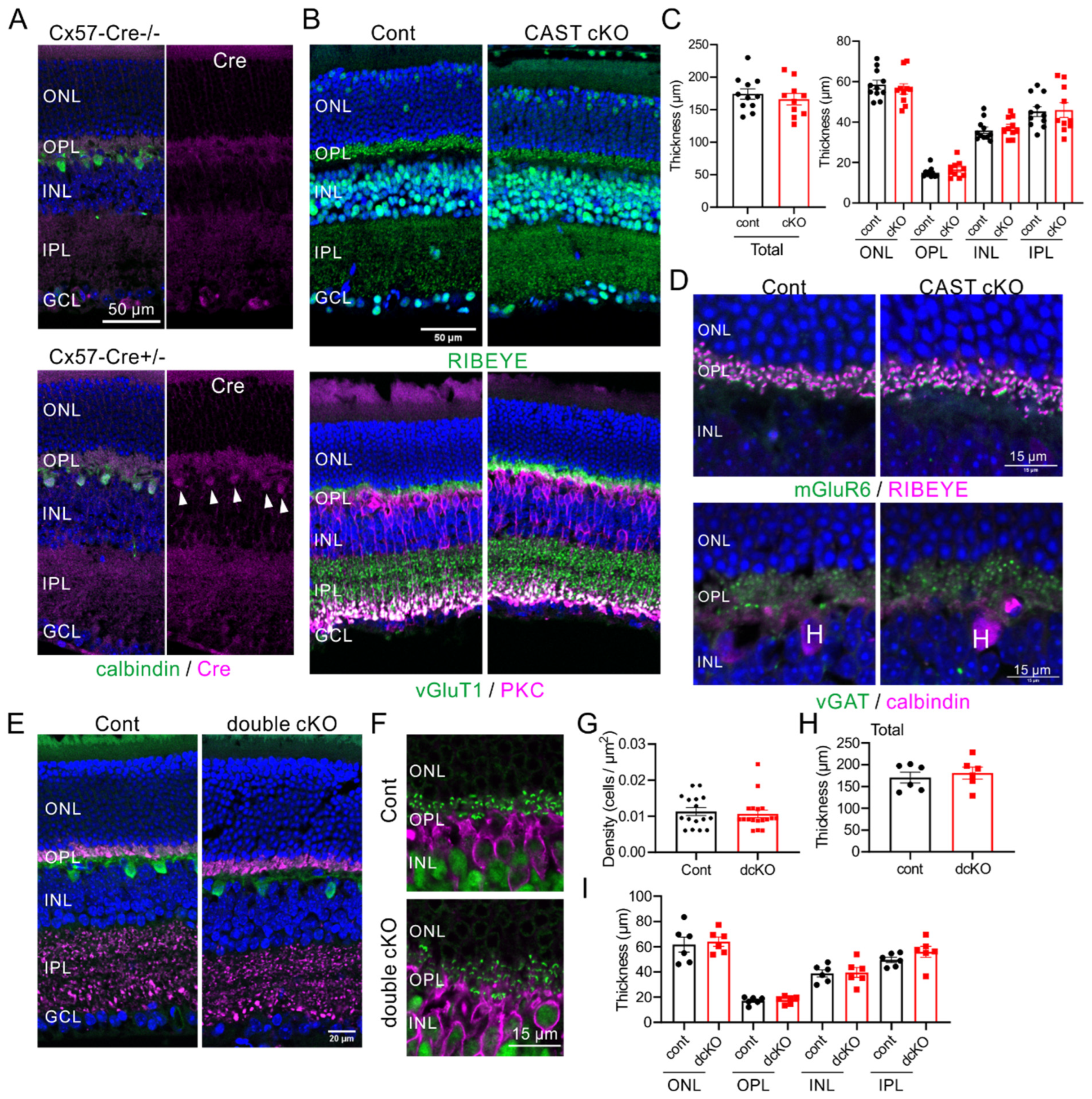
2.2. Depletion of CAST and ELKS in Photoreceptor Neurons Does Not Affect Retina Formation
2.3. CAST and ELKS in Horizontal Cells Do Not Play a Major Role in the Formation of the Retina
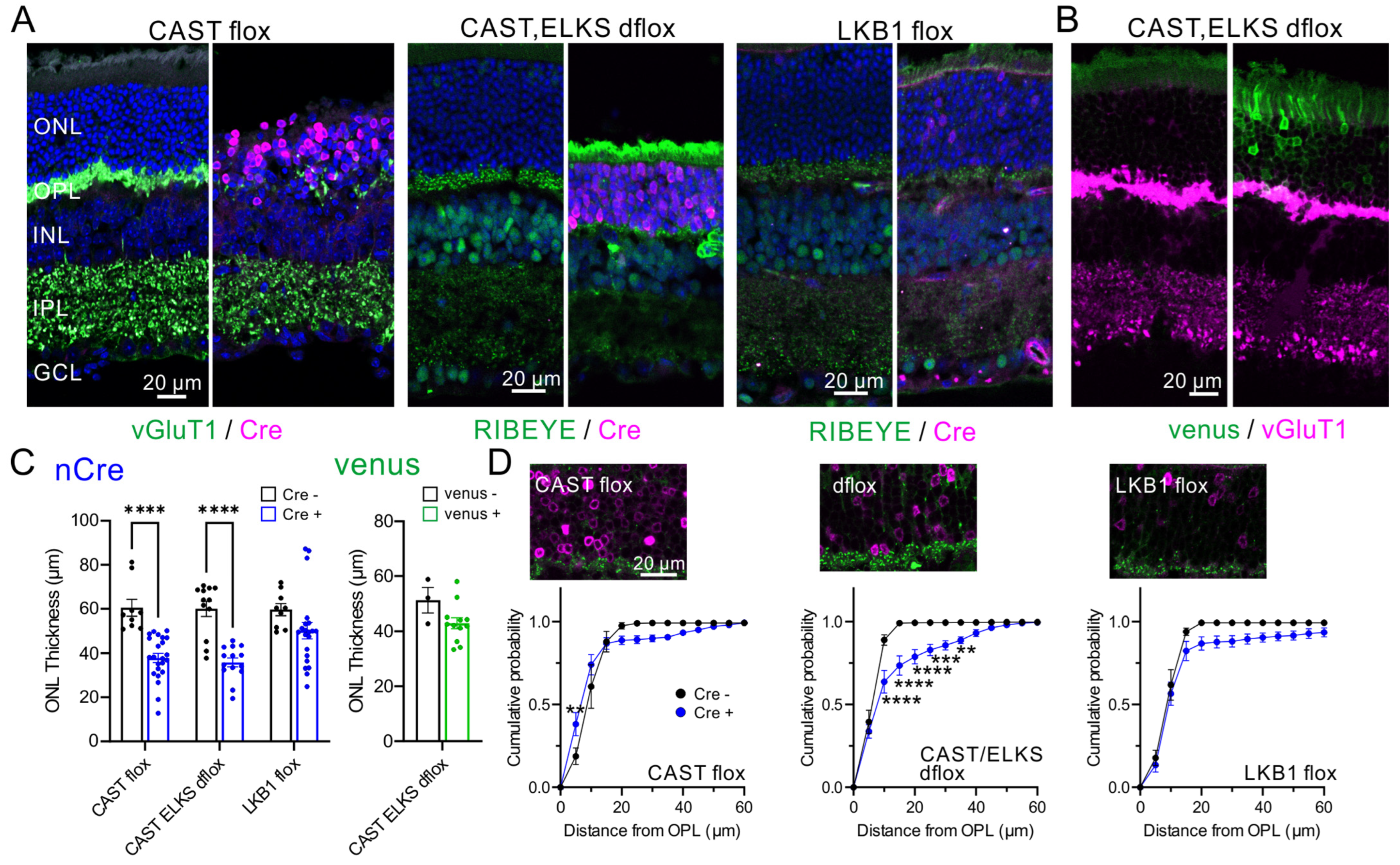
2.4. Acute Ablation of CAST and ELKS Causes Photoreceptor Degeneration
3. Discussion
4. Materials and Methods
4.1. Mouse Lines
4.2. Immunohistochemistry and Image Analysis
4.3. Immunoblotting
4.4. AAV5-Mediated Cre Expression and Image Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hatanaka, Y.; Hirata, T. How Do Cortical Excitatory Neurons Terminate Their Migration at the Right Place? Critical Roles of Environmental Elements. Front. Cell Dev. Biol. 2020, 8, 596708. [Google Scholar] [CrossRef]
- Stouffer, M.A.; Golden, J.A.; Francis, F. Neuronal Migration Disorders: Focus on the Cytoskeleton and Epilepsy. Neurobiol. Dis. 2016, 92, 18–45. [Google Scholar] [CrossRef]
- Sarin, S.; Zuniga-Sanchez, E.; Kurmangaliyev, Y.Z.; Cousins, H.; Patel, M.; Hernandez, J.; Zhang, K.X.; Samuel, M.A.; Morey, M.; Sanes, J.R.; et al. Role for Wnt Signaling in Retinal Neuropil Development: Analysis via RNA-Seq and In Vivo Somatic CRISPR Mutagenesis. Neuron 2018, 98, 109–126.e8. [Google Scholar] [CrossRef] [Green Version]
- Dellett, M.; Sasai, N.; Nishide, K.; Becker, S.; Papadaki, V.; Astrid Limb, G.; Moore, A.T.; Kondo, T.; Ohnuma, S.I. Genetic Background and Light-Dependent Progression of Photoreceptor Cell Degeneration in Prominin-1 Knockout Mice. Investig. Ophthalmol. Vis. Sci. 2015, 56, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Fernández, M.; Uribe, M.L.; Vicente-Tejedor, J.; Germain, F.; Susín-Lara, C.; Quereda, C.; Montoliu, L.; De La Villa, P.; Martín-Nieto, J.; Cruces, J. Impairment of Photoreceptor Ribbon Synapses in a Novel Pomt1 Conditional Knockout Mouse Model of Dystroglycanopathy. Sci. Rep. 2018, 8, 8543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claes, E.; Seeliger, M.; Michalakis, S.; Biel, M.; Humphries, P.; Haverkamp, S. Morphological Characterization of the Retina of the CNGA3-/- Rho-/- Mutant Mouse Lacking Functional Cones and Rods. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2039–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- tom Dieck, S.; Specht, D.; Strenzke, N.; Hida, Y.; Krishnamoorthy, V.; Schmidt, K.-F.; Inoue, E.; Ishizaki, H.; Tanaka-Okamoto, M.; Miyoshi, J.; et al. Deletion of the Presynaptic Scaffold CAST Reduces Active Zone Size in Rod Photoreceptors and Impairs Visual Processing. J. Neurosci. 2012, 32, 12192–12203. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, A.; Kitahara, Y.; Grabner, C.P.; Vogl, C.; Abe, M.; Kitta, R.; Ohta, K.; Nakamura, K.; Sakimura, K.; Moser, T.; et al. Cytomatrix Proteins CAST and ELKS Regulate Retinal Photoreceptor Development and Maintenance. J. Cell Biol. 2018, 217, 3993–4006. [Google Scholar] [CrossRef]
- Chang, B.; Heckenlively, J.R.; Bayley, P.R.; Brecha, N.C.; Davisson, M.T.; Hawes, N.L.; Hirano, A.A.; Hurd, R.E.; Ikeda, A.; Johnson, B.A.; et al. The Nob2 Mouse, a Null Mutation in Cacna1f: Anatomical and Functional Abnormalities in the Outer Retina and Their Consequences on Ganglion Cell Visual Responses. Vis. Neurosci. 2006, 23, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Specht, D.; Wu, S.B.; Turner, P.; Dearden, P.; Koentgen, F.; Wolfrum, U.; Maw, M.; Brandstätter, J.H.; Dieck, S.T. Effects of Presynaptic Mutations on a Postsynaptic Cacna1s Calcium Channel Colocalized with MGluR6 at Mouse Photoreceptor Ribbon Synapses. Investig. Ophthalmol. Vis. Sci. 2009, 50, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Boycott, K.M.; Maybaum, T.A.; Naylor, M.J.; Weleber, R.G.; Robitaille, J.; Miyake, Y.; Bergen, A.A.B.; Pierpont, M.E.; Pearce, W.G.; Bech-Hansen, N.T. A Summary of 20 CACNA1F Mutations Identified in 36 Families with Incomplete X-Linked Congenital Stationary Night Blindness, and Characterization of Splice Variants. Hum. Genet. 2001, 108, 91–97. [Google Scholar] [CrossRef]
- Samuel, M.A.; Voinescu, P.E.; Lilley, B.N.; De Cabo, R.; Foretz, M.; Viollet, B.; Pawlyk, B.; Sandberg, M.A.; Vavvas, D.G.; Sanes, J.R. LKB1 and AMPK Regulate Synaptic Remodeling in Old Age. Nat. Neurosci. 2014, 17, 1190–1197. [Google Scholar] [CrossRef] [Green Version]
- Burger, C.A.; Alevy, J.; Casasent, A.K.; Jiang, D.; Albrecht, N.E.; Liang, J.H.; Hirano, A.A.; Brecha, N.C.; Samuel, M.A. Lkb1 Coordinates Neurite Remodeling to Drive Synapse Layer Emergence in the Outer Retina. eLife 2020, 9, e56931. [Google Scholar] [CrossRef]
- Dick, O.; tom Dieck, S.; Altrock, W.D.; Ammermüller, J.; Weiler, R.; Garner, C.C.; Gundelfinger, E.D.; Brandstätter, J.H. The Presynaptic Active Zone Protein Bassoon Is Essential for Photoreceptor Ribbon Synapse Formation in the Retina. Neuron 2003, 37, 775–786. [Google Scholar] [CrossRef] [Green Version]
- D’Orazi, F.D.; Suzuki, S.C.; Wong, R.O. Neuronal Remodeling in Retinal Circuit Assembly, Disassembly, and Reassembly. Trends Neurosci. 2014, 37, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Maxeiner, S.; Luo, F.; Tan, A.; Schmitz, F.; Südhof, T.C. How to Make a Synaptic Ribbon: RIBEYE Deletion Abolishes Ribbons in Retinal Synapses and Disrupts Neurotransmitter Release. EMBO J. 2016, 35, 1098–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonntag, S.; Dedek, K.; Dorgau, B.; Schultz, K.; Schmidt, K.F.; Cimiotti, K.; Weiler, R.; Löwel, S.; Willecke, K.; Janssen-Bienhold, U. Ablation of Retinal Horizontal Cells from Adult Mice Leads to Rod Degeneration and Remodeling in the Outer Retina. J. Neurosci. 2012, 32, 10713–10724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Sarria, I.; Fehlhaber, K.E.; Kamasawa, N.; Orlandi, C.; James, K.N.; Hazen, J.L.; Gardner, M.R.; Farzan, M.; Lee, A.; et al. Mechanism for Selective Synaptic Wiring of Rod Photoreceptors into the Retinal Circuitry and Its Role in Vision. Neuron 2015, 87, 1248–1260. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fajardo, D.; Guerrero-Given, D.; Samuel, M.A.; Ohtsuka, T.; Boye, S.E.; Kamasawa, N.; Martemyanov, K.A. Post-Developmental Plasticity of the Primary Rod Pathway Allows Restoration of Visually Guided Behaviors. Curr. Biol. 2022, 32, 4783–4796.e3. [Google Scholar] [CrossRef]
- Furukawa, T.; Morrow, E.M.; Cepko, C.L. Crx, a Novel Otx-like Homeobox Gene, Shows Photoreceptor-Specific Expression and Regulates Photoreceptor Differentiation. Cell 1997, 91, 531–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, A.; Furukawa, A.; Koike, C.; Tano, Y.; Aizawa, S.; Matsuo, I.; Furukawa, T. Otx2 Homeobox Gene Controls Retinal Photoreceptor Cell Fate and Pineal Gland Development. Nat. Neurosci. 2003, 6, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Deguchi-Tawarada, M.; Inoue, E.; Takao-Kikitsu, E.; Inoue, M.; Kitajima, I.; Ohtsuka, T.; Takai, Y. Active Zone Protein CAST Is a Component of Conventional and Ribbon Synapses in Mouse Retina. J. Comp. Neurol. 2006, 495, 480–496. [Google Scholar] [CrossRef]
- Kobayashi, S.; Hida, Y.; Ishizaki, H.; Inoue, E.; Tanaka-Okamoto, M.; Yamasaki, M.; Miyazaki, T.; Fukaya, M.; Kitajima, I.; Takai, Y.; et al. The Active Zone Protein CAST Regulates Synaptic Vesicle Recycling and Quantal Size in the Mouse Hippocampus. Eur. J. Neurosci. 2016, 44, 2272–2284. [Google Scholar] [CrossRef]
- Ströh, S.; Sonntag, S.; Janssen-Bienhold, U.; Schultz, K.; Cimiotti, K.; Weiler, R.; Willecke, K.; Dedek, K. Cell-Specific Cre Recombinase Expression Allows Selective Ablation of Glutamate Receptors from Mouse Horizontal Cells. PLoS ONE 2013, 8, e83076. [Google Scholar] [CrossRef] [Green Version]
- Sudhof, T.C. The Presynaptic Active Zone. Neuron 2012, 75, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acuna, C.; Liu, X.; Südhof, T.C. How to Make an Active Zone: Unexpected Universal Functional Redundancy between RIMs and RIM-BPs. Neuron 2016, 91, 792–807. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.S.H.; Held, R.G.; Wong, M.Y.; Liu, C.; Karakhanyan, A.; Kaeser, P.S. Fusion Competent Synaptic Vesicles Persist upon Active Zone Disruption and Loss of Vesicle Docking. Neuron 2016, 91, 777–791. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.; Li, S. Liver Kinase B1 Functions as a Regulator for Neural Development and a Therapeutic Target for Neural Repair. Cells 2022, 11, 2861. [Google Scholar] [CrossRef]
- Barnes, A.P.; Lilley, B.N.; Pan, Y.A.; Plummer, L.J.; Powell, A.W.; Raines, A.N.; Sanes, J.R.; Polleux, F. LKB1 and SAD Kinases Define a Pathway Required for the Polarization of Cortical Neurons. Cell 2007, 129, 549–563. [Google Scholar] [CrossRef] [Green Version]
- Shelly, M.; Poo, M.M. Role of LKB1-SAD/MARK Pathway in Neuronal Polarization. Dev. Neurobiol. 2011, 71, 508–527. [Google Scholar] [CrossRef]
- Mochida, S.; Hida, Y.; Tanifuji, S.; Hagiwara, A.; Hamada, S.; Abe, M.; Ma, H.; Yasumura, M.; Kitajima, I.; Sakimura, K.; et al. SAD-B Phosphorylation of CAST Controls Active Zone Vesicle Recycling for Synaptic Depression. Cell Rep. 2016, 16, 2901–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Sugita, S.; Kurimoto, Y.; Takahashi, M. Trends of Stem Cell Therapies in Age-Related Macular Degeneration. J. Clin. Med. 2021, 10, 1785. [Google Scholar] [CrossRef] [PubMed]
- Nakada, D.; Saunders, T.L.; Morrison, S.J. Lkb1 Regulates Cell Cycle and Energy Metabolism in Haematopoietic Stem Cells. Nature 2010, 468, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, M.; Yuasa, S.; Fujisawa, H.; Naruse, I.; Saga, S.; Mishina, M.; Yagi, T. Disruption of Semaphorin III/D Gene Causes Severe Abnormality in Peripheral Nerve Projection. Neuron 1997, 19, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Mishina, M.; Sakimura, K. Conditional Gene Targeting on the Pure C57BL/6 Genetic Background. Neurosci. Res. 2007, 58, 105–112. [Google Scholar] [CrossRef]
- Takeuchi, T.; Miyazaki, T.; Watanabe, M.; Mori, H.; Sakimura, K.; Mishina, M. Control of Synaptic Connection by Glutamate Receptor Δ2 in the Adult Cerebellum. J. Neurosci. 2005, 25, 2146–2156. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, T.; Nomura, T.; Tsujita, M.; Suzuki, M.; Fuse, T.; Mori, H.; Mishina, M. Flp Recombinase Transgenic Mice of C57BL/6 Strain for Conditional Gene Targeting. Biochem. Biophys. Res. Commun. 2002, 293, 953–957. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Radulovic, T.; Goral, R.O.; Thomas, C.; Suarez Montesinos, M.; Guerrero-Given, D.; Hagiwara, A.; Putzke, T.; Hida, Y.; Abe, M.; et al. CAST/ELKS Proteins Control Voltage-Gated Ca2+ Channel Density and Synaptic Release Probability at a Mammalian Central Synapse. Cell Rep. 2018, 24, 284–293.e6. [Google Scholar] [CrossRef] [Green Version]
- Kono, J.; Konno, K.; Talukder, A.H.; Fuse, T.; Abe, M.; Uchida, K.; Horio, S.; Sakimura, K.; Watanabe, M.; Itoi, K. Distribution of Corticotropin-Releasing Factor Neurons in the Mouse Brain: A Study Using Corticotropin-Releasing Factor-Modified Yellow Fluorescent Protein Knock-in Mouse. Brain Struct. Funct. 2017, 222, 1705–1732. [Google Scholar] [CrossRef]
- Cao, Y.; Posokhova, E.; Martemyanov, K.A. TRPM1 Forms Complexes with Nyctalopin in Vivo and Accumulates in Postsynaptic Compartment of ON-Bipolar Neurons in MGLuR6-Dependent Manner. J. Neurosci. 2011, 31, 11521–11526. [Google Scholar] [CrossRef] [Green Version]
- Deguchi-Tawarada, M.; Inoue, E.; Takao-Rikitsu, E.; Inoue, M.; Ohtsuka, T.; Takai, Y. CAST2: Identification and Characterization of a Protein Structurally Related to the Presynaptic Cytomatrix Protein CAST. Genes Cells 2004, 9, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, T.; Takao-Rikitsu, E.; Inoue, E.; Inoue, M.; Takeuchi, M.; Matsubara, K.; Deguchi-Tawarada, M.; Satoh, K.; Morimoto, K.; Nakanishi, H.; et al. CAST: A Novel Protein of the Cytomatrix at the Active Zone of Synapses That Forms a Ternary Complex with RIM1 and Munc13-1. J. Cell Biol. 2002, 158, 577–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hagiwara, A.; Mizutani, A.; Kawamura, S.; Abe, M.; Hida, Y.; Sakimura, K.; Ohtsuka, T. Critical Role of the Presynaptic Protein CAST in Maintaining the Photoreceptor Ribbon Synapse Triad. Int. J. Mol. Sci. 2023, 24, 7251. https://doi.org/10.3390/ijms24087251
Hagiwara A, Mizutani A, Kawamura S, Abe M, Hida Y, Sakimura K, Ohtsuka T. Critical Role of the Presynaptic Protein CAST in Maintaining the Photoreceptor Ribbon Synapse Triad. International Journal of Molecular Sciences. 2023; 24(8):7251. https://doi.org/10.3390/ijms24087251
Chicago/Turabian StyleHagiwara, Akari, Ayako Mizutani, Saki Kawamura, Manabu Abe, Yamato Hida, Kenji Sakimura, and Toshihisa Ohtsuka. 2023. "Critical Role of the Presynaptic Protein CAST in Maintaining the Photoreceptor Ribbon Synapse Triad" International Journal of Molecular Sciences 24, no. 8: 7251. https://doi.org/10.3390/ijms24087251