
Biological Activities of Miracle Berry Supercritical Extracts as Metabolic Regulators in Chronic Diseases
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Extraction Yield
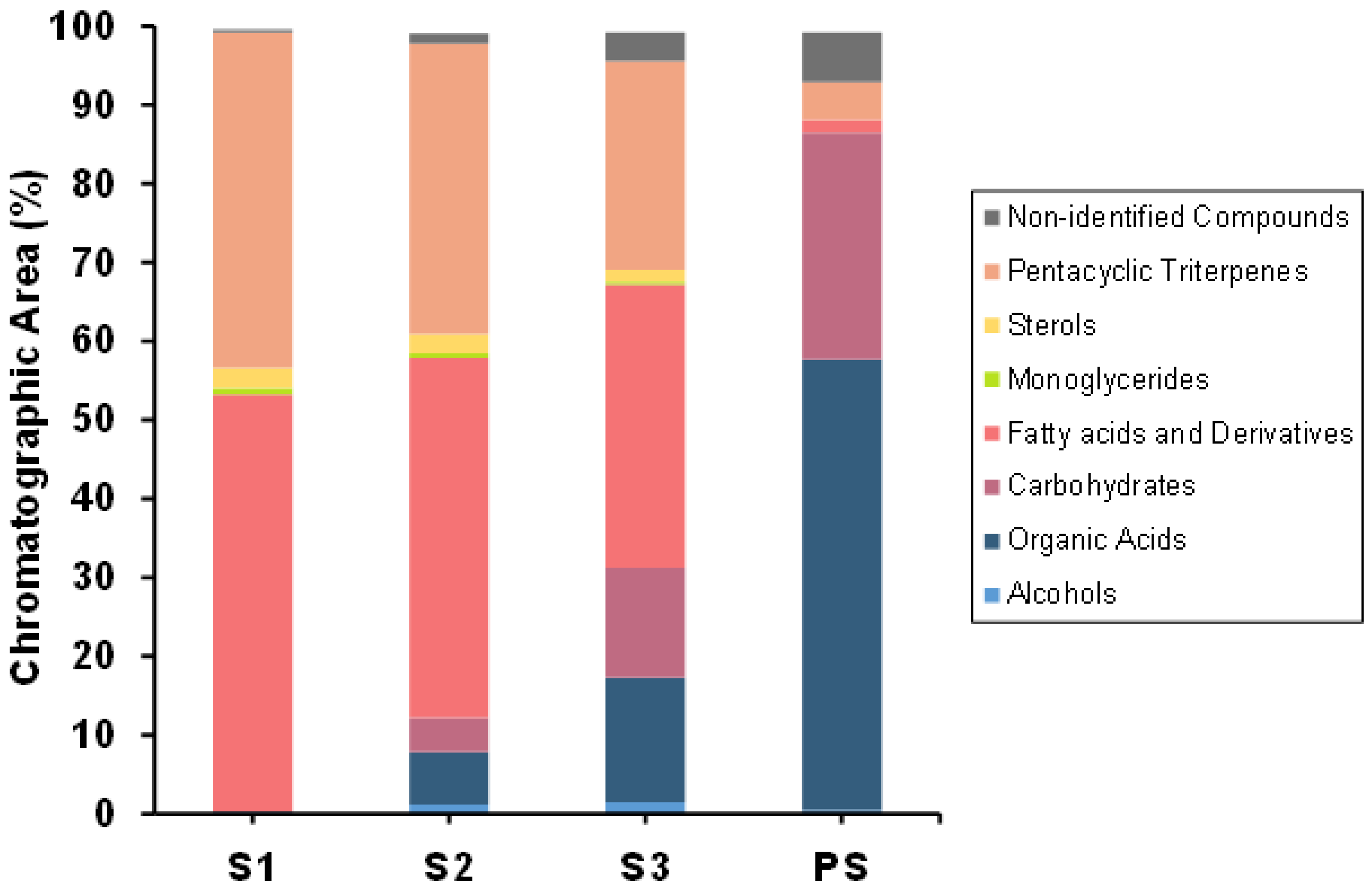
2.2. Chemical Composition of SFE Extracts
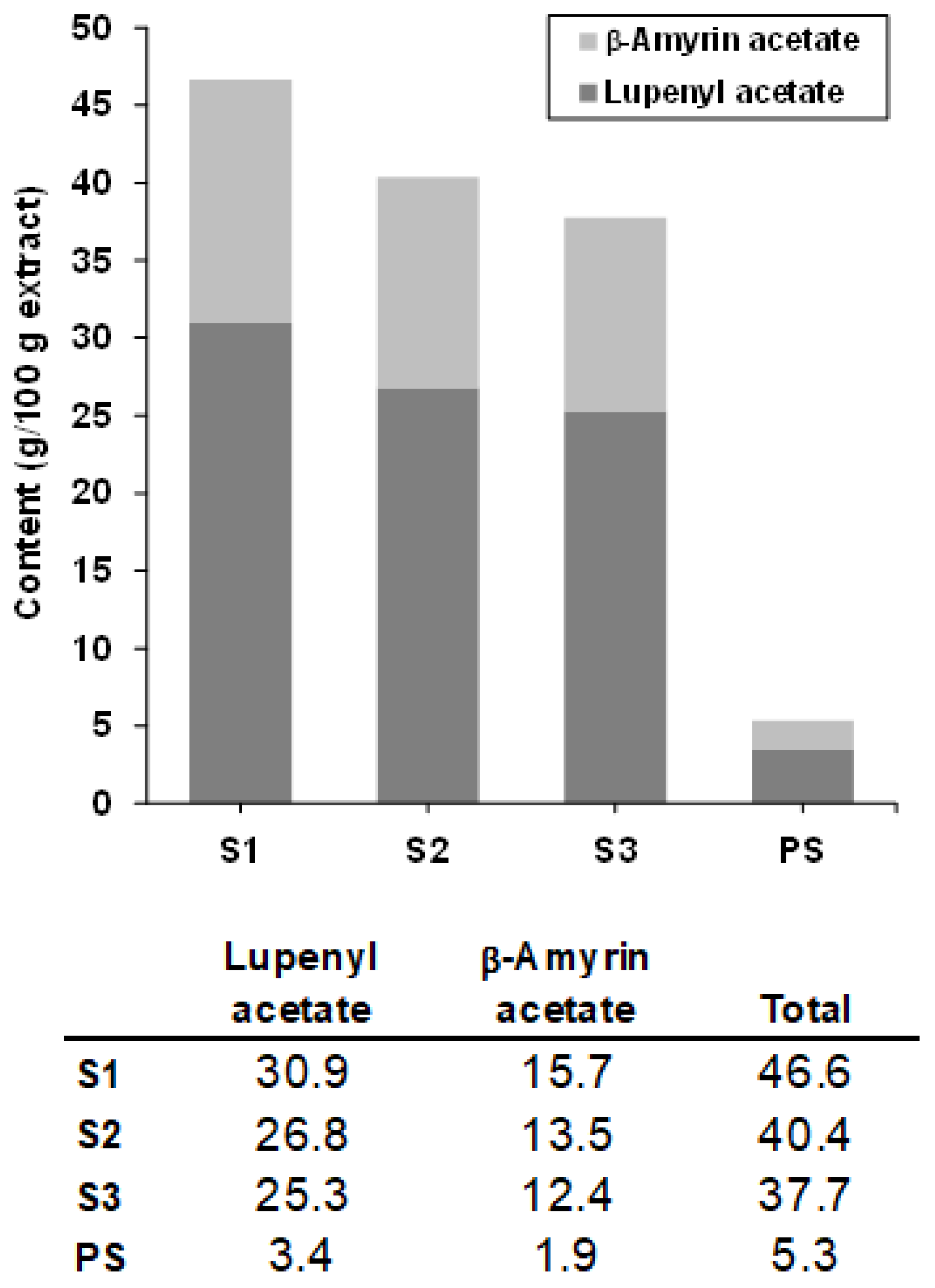
2.2.1. Chemical Characterization by GC-MS
2.2.2. Determination of the Total Phenolic Content (TPC) and Individual Phenolic Compounds by HPLC-PAD
2.3. Antioxidant Activity of SFE MB Extracts
2.3.1. In Vitro Antioxidant Activity
2.3.2. Cellular Antioxidant Activity
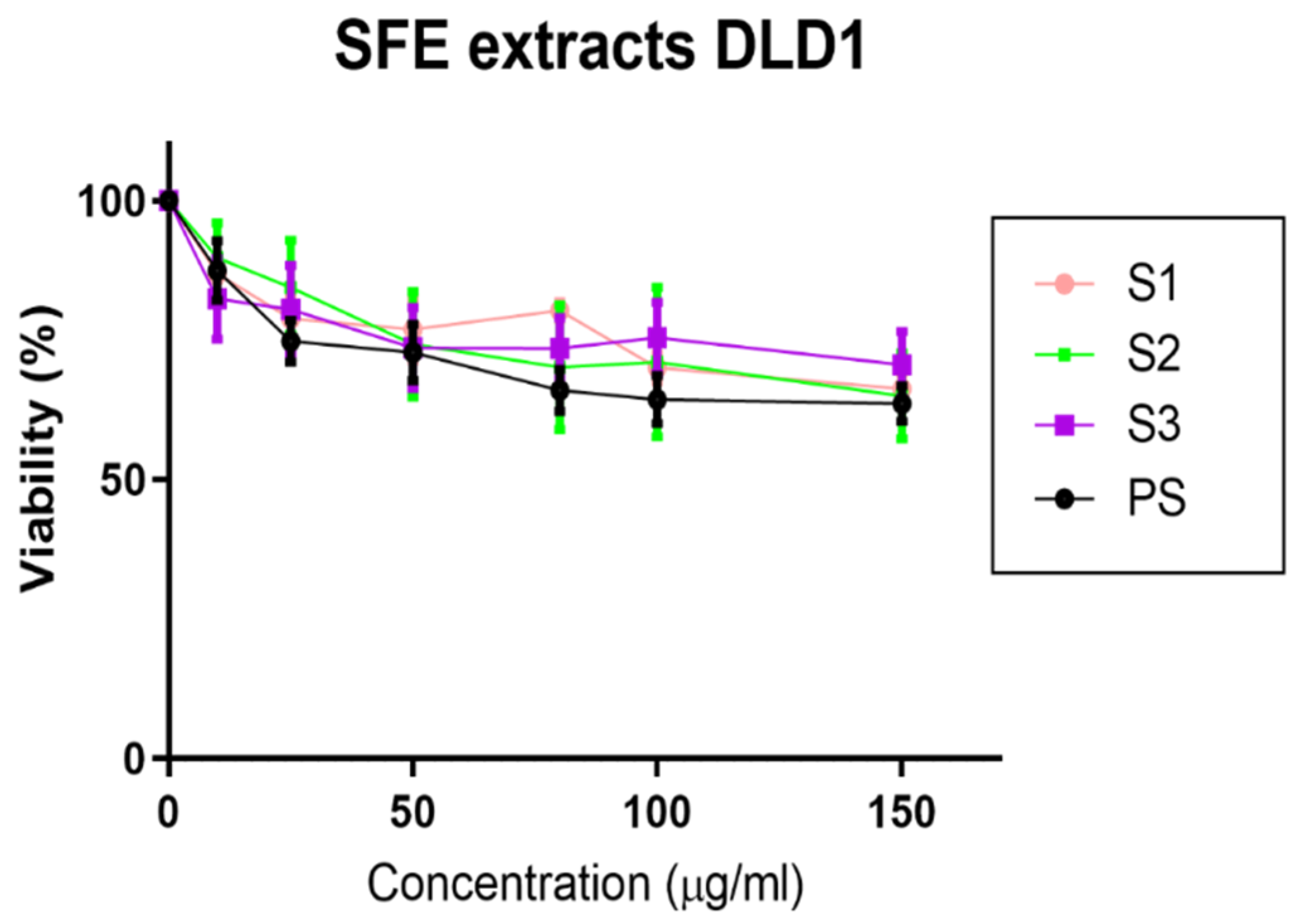
2.4. Effect of MB SFE Extracts on the Inhibition of Cell Viability of Colorectal Cancer Cells
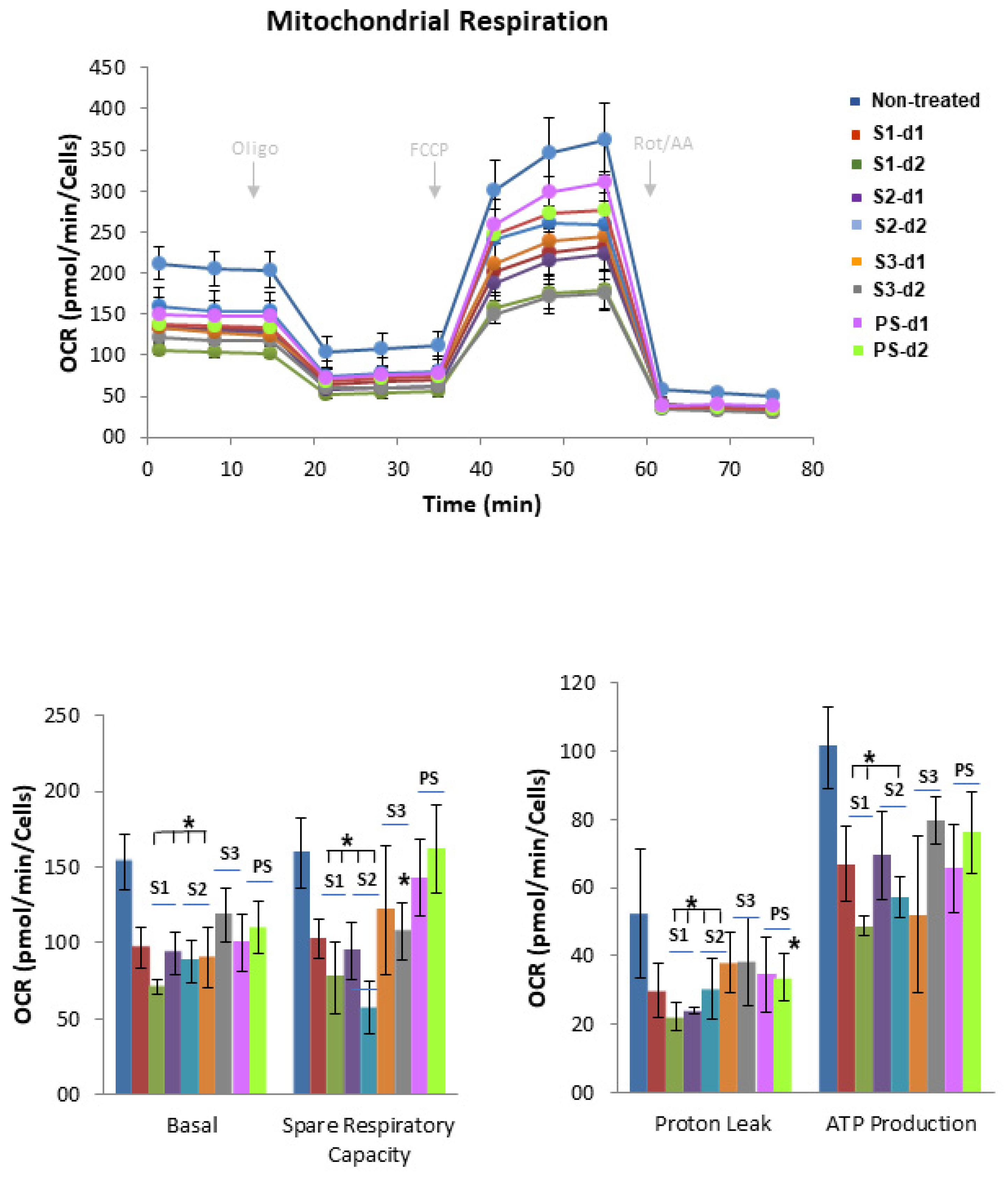
2.5. Effect of MB SFE Extracts on the Bioenergetic Profile of Colorectal Cancer Cells
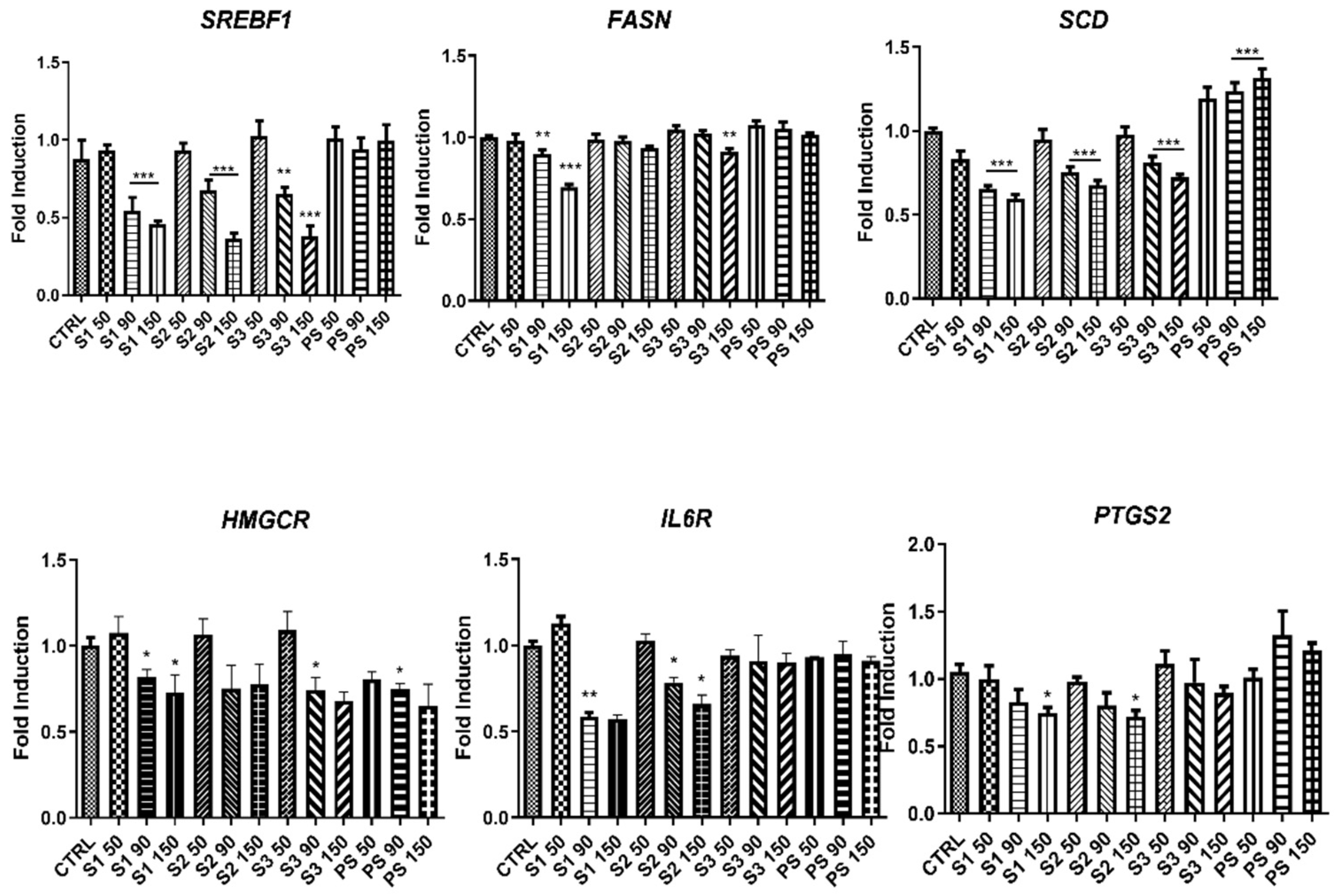
2.6. Influence of MB SFE Extracts in the Expression of Lipid Metabolism Genes
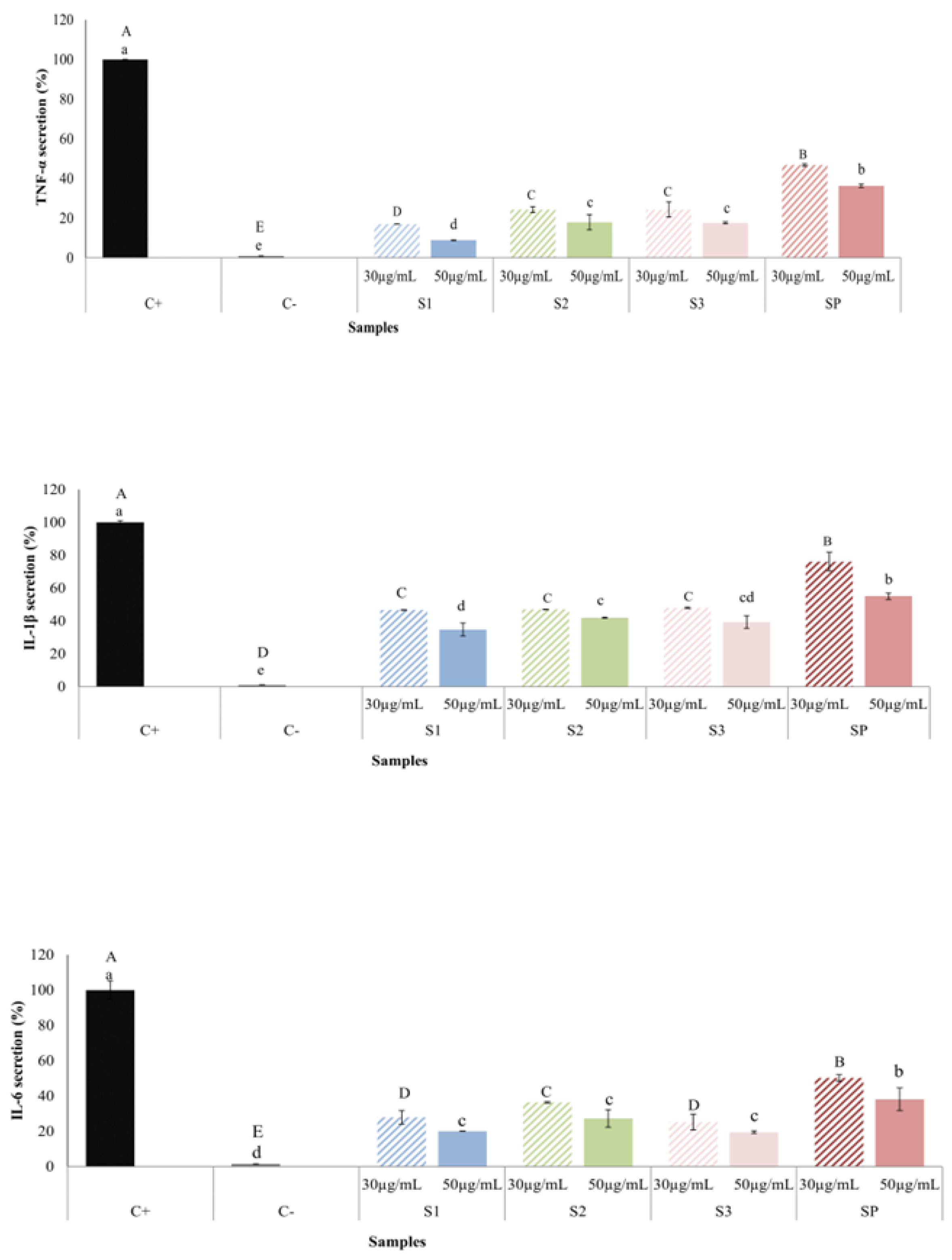
2.7. Anti-Inflammatory Effect of MB SFE Extracts
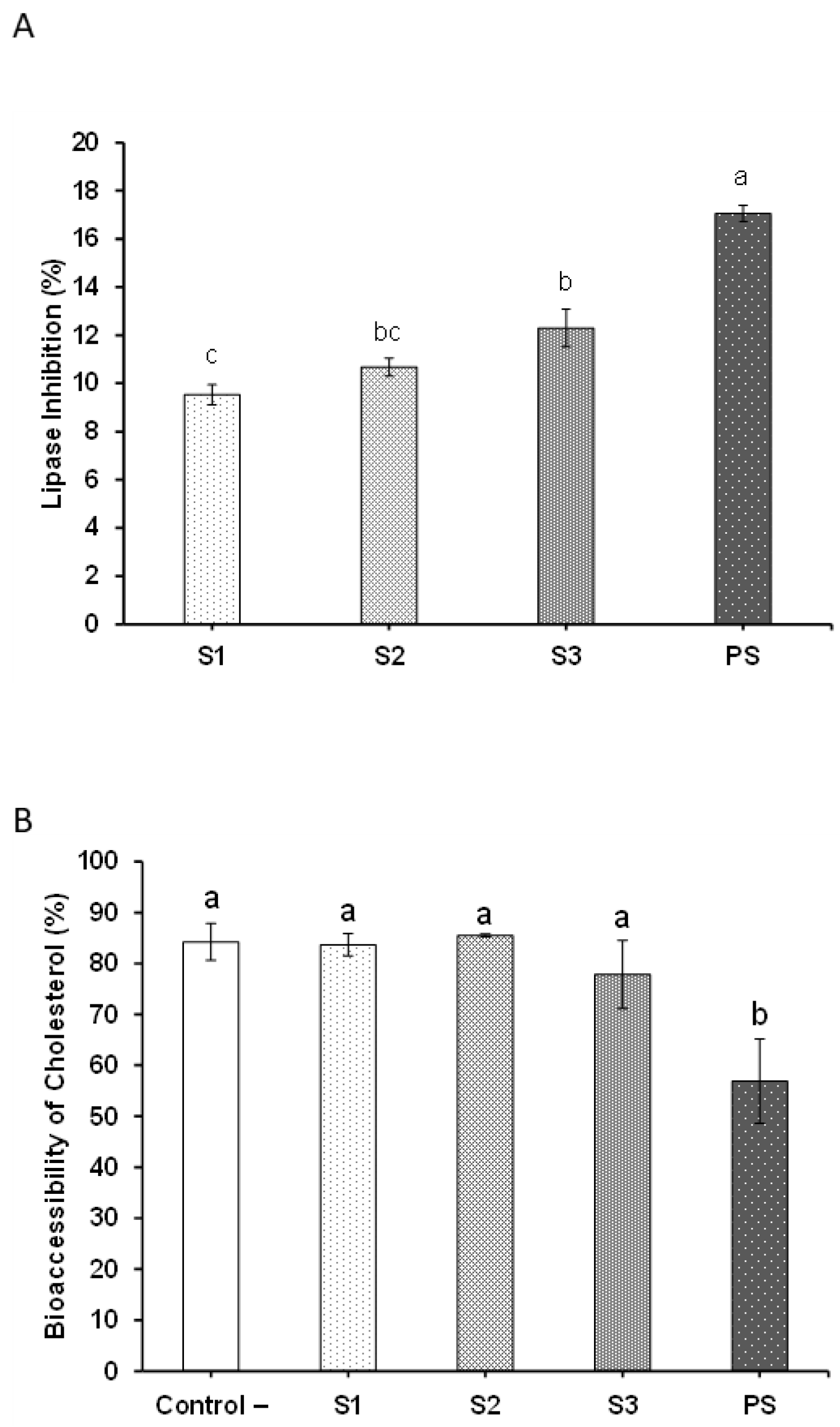
2.8. Potential Hypolipidemic Effects of MB SFE Extracts
3. Discussion
4. Materials and Methods
4.1. Reagents and Cell Culture
4.2. Supercritical Fluid Extraction of MB Extracts
4.3. Determination of Phenolic Compounds in the Supercritical Fluid Extracts from MB
4.4. Characterization of the Supercritical Fluid Extracts by Gas Chromatography–Mass Spectrometry (GC-MS)
4.5. Measurement of the Antioxidant Activity of Supercritical Fluid Extracts from MB
4.6. Analysis of the Immunomodulatory Activity of Supercritical Fluid Extracts from MB
4.7. Analysis of the Inhibition of Cell Viability of Supercritical Fluid Extracts from MB
4.8. Gene Expression Analysis
4.9. Analysis of Mitochondrial Respiration by Extracellular Flux Analysis of the Oxygen Consumption Rate (OCR)
4.10. Hypolipidemic Activity
4.10.1. Pancreatic Lipase Inhibition Assay
4.10.2. Effect on the Intestinal Bioaccessibility of Cholesterol
4.11. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buckmire, R.E.; Francis, F.J. Anthocyanins and Flavonols of Miracle Fruit, Synsepalum dulcificum, Schum. J. Food Sci. 1976, 41, 1363–1365. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Dried Fruits of Synsepalum dulcificum as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFS2 2021, 19, e06600. [Google Scholar] [CrossRef]
- Belloir, C.; Neiers, F.; Briand, L. Sweeteners and Sweetness Enhancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Inglett, G.E.; Chen, D. Contents of Phenolics and Flavonoids and Antioxidant Activities in Skin, Pulp, and Seeds of Miracle Fruit. J. Food Sci. 2011, 76, C479–C482. [Google Scholar] [CrossRef]
- Wilken, M.K.; Satiroff, B.A. Pilot Study of “Miracle Fruit” to Improve Food Palatability for Patients Receiving Chemotherapy. Clin. J. Oncol. Nurs. 2012, 16, E173–E177. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Shen, Y.; Zhang, X.; Prinyawiwatkul, W.; Xu, Z. Antioxidant-Rich Phytochemicals in Miracle Berry (Synsepalum dulcificum) and Antioxidant Activity of Its Extracts. Food Chem. 2014, 153, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-M.; Chou, Y.-T.; Hong, Z.-L.; Chen, H.-A.; Chang, Y.-C.; Yang, W.-L.; Chang, H.-C.; Mai, C.-T.; Chen, C.-Y. Bioconstituents from Stems of Synsepalum dulcificum Daniell (Sapotaceae) Inhibit Human Melanoma Proliferation, Reduce Mushroom Tyrosinase Activity and Have Antioxidant Properties. J. Taiwan Inst. Chem. Eng. 2011, 42, 204–211. [Google Scholar] [CrossRef]
- Seong, J.; Oyong, G.; Cabrera, E. Synsepalum dulcificum Extracts Exhibit Cytotoxic Activity on Human Colorectal Cancer Cells and Upregulate C-Fos and c-Jun Early Apoptotic Gene Expression. Asian Pac. J. Trop. Biomed. 2018, 8, 173. [Google Scholar] [CrossRef]
- World Cancer Research Fund International. Colorectal Cancer Statistics. Available online: https://www.wcrf.org/cancer-trends/colorectal-cancer-statistics/ (accessed on 23 February 2022).
- Johnson, C.M.; Wei, C.; Ensor, J.E.; Smolenski, D.J.; Amos, C.I.; Levin, B.; Berry, D.A. Meta-Analyses of Colorectal Cancer Risk Factors. Cancer Causes Control. 2013, 24, 1207–1222. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-C.; Liu, I.-M.; Cheng, J.-T. Improvement of Insulin Resistance by Miracle Fruit (Synsepalum dulcificum) in Fructose-Rich Chow-Fed Rats. Phytother. Res. 2006, 20, 987–992. [Google Scholar] [CrossRef]
- Han, Y.; Wu, J.; Wang, C. Modulatory Effects of Miracle Fruit Ethanolic Extracts on Glucose Uptake through the Insulin Signaling Pathway in C2C12 Mouse Myotubes Cells. Food Sci. Nutr. 2019, 7, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez de Cedrón, M.; Wagner, S.; Reguero, M.; Menéndez-Rey, A.; Ramírez de Molina, A. Miracle Berry as a Potential Supplement in the Control of Metabolic Risk Factors in Cancer. Antioxidants 2020, 9, 1282. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Kumar, P.; Deshmukh, R.R.; Bishayee, A.; Kumar, S. Pentacyclic Triterpenes: New Tools to Fight Metabolic Syndrome. Phytomedicine 2018, 50, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.-Y.; Huang, S.-T.; Lin, M.-L.; Lai, J.-T. Polyphenol Measurement and Antioxidant Activity of Miracle Fruit. Int. J. Chem. Eng. Appl. 2015, 6, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-G.; Li, B.; Fu, Q.; Zhang, X.-M.; Ma, F.-Y.; Hu, Y. Miracle Fruit Leaf Extract: Antioxidant Activity Evaluation, Constituent Identification, and Medical Applications. Anal. Lett. 2021, 54, 2211–2226. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, F.; Zhang, L.; Wu, Y.; Hu, B.; Zhang, Y.; Li, Y.; Liu, H. Growth Inhibition and Apoptosis Induced by Lupeol, a Dietary Triterpene, in Human Hepatocellular Carcinoma Cells. Biol. Pharm. Bull. 2011, 34, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; He, C.; Xu, F.; Xu, X.; Liu, L.; Xu, M.; Guo, Z.; Wang, Y.; Liao, J.; Li, Y. Lupeol Inhibits Osteosarcoma Progression by Up-Regulation of HMGA2 via Regulating MiR-212-3p. J. Orthop. Surg. Res. 2020, 15, 374. [Google Scholar] [CrossRef]
- Tarapore, R.S.; Siddiqui, I.A.; Adhami, V.M.; Spiegelman, V.S.; Mukhtar, H. The Dietary Terpene Lupeol Targets Colorectal Cancer Cells with Constitutively Active Wnt/β-Catenin Signaling. Mol. Nutr. Food Res. 2013, 57, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- DeBerardinis, R.J.; Lum, J.J.; Hatzivassiliou, G.; Thompson, C.B. The Biology of Cancer: Metabolic Reprogramming Fuels Cell Growth and Proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Vargas, T.; Moreno-Rubio, J.; Herranz, J.; Cejas, P.; Molina, S.; González-Vallinas, M.; Mendiola, M.; Burgos, E.; Aguayo, C.; Custodio, A.B.; et al. ColoLipidGene: Signature of Lipid Metabolism-Related Genes to Predict Prognosis in Stage-II Colon Cancer Patients. Oncotarget 2015, 6, 7348–7363. [Google Scholar] [CrossRef] [Green Version]
- Menendez, J.A.; Lupu, R. Fatty Acid Synthase and the Lipogenic Phenotype in Cancer Pathogenesis. Nat. Rev. Cancer 2007, 7, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Glynne-Jones, R.; Wyrwicz, L.; Tiret, E.; Brown, G.; Rödel, C.; Cervantes, A.; Arnold, D. Rectal Cancer: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2017, 28, iv22–iv40. [Google Scholar] [CrossRef] [PubMed]
- Carethers, J.M. Review: Systemic Treatment of Advanced Colorectal Cancer: Tailoring Therapy to the Tumor. Therap. Adv. Gastroenterol. 2008, 1, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Melino, G.; Amelio, I.; Jiang, J.; Wang, Y.; Shi, Y. Recent Advances in Cancer Immunotherapy. Discov. Onc. 2021, 12, 27. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.P.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer Prevention and Therapy through the Modulation of Transcription Factors by Bioactive Natural Compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef]
- Gómez de Cedrón, M.; Vargas, T.; Madrona, A.; Jiménez, A.; Pérez-Pérez, M.-J.; Quintela, J.-C.; Reglero, G.; San-Félix, A.; Ramírez de Molina, A. Novel Polyphenols That Inhibit Colon Cancer Cell Growth Affecting Cancer Cell Metabolism. J. Pharmacol. Exp. Ther. 2018, 366, 377–389. [Google Scholar] [CrossRef]
- Jimenez-Garcia, S.N.; Guevara-Gonzalez, R.G.; Miranda-Lopez, R.; Feregrino-Perez, A.A.; Torres-Pacheco, I.; Vazquez-Cruz, M.A. Functional Properties and Quality Characteristics of Bioactive Compounds in Berries: Biochemistry, Biotechnology, and Genomics. Food Res. Int. 2013, 54, 1195–1207. [Google Scholar] [CrossRef]
- Obafemi, T.O.; Akinmoladun, A.C.; Olaleye, M.T.; Agboade, S.O.; Onasanya, A.A. Antidiabetic Potential of Methanolic and Flavonoid-Rich Leaf Extracts of Synsepalum dulcificum in Type 2 Diabetic Rats. J. Ayurveda Integr. Med. 2017, 8, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Chung, H.Y.; Xuan, W.; Wang, G.; Li, Y. The Cholesterol-lowering Activity of Miracle Fruit (Synsepalum dulcificum). J. Food Biochem. 2020, 44, e13185. [Google Scholar] [CrossRef]
- Shi, Y.-C.; Lin, K.-S.; Jhai, Y.-F.; Lee, B.-H.; Han, Y.; Cui, Z.; Hsu, W.-H.; Wu, S.-C. Miracle Fruit (Synsepalum dulcificum) Exhibits as a Novel Anti-Hyperuricaemia Agent. Molecules 2016, 21, 140. [Google Scholar] [CrossRef] [Green Version]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid Metabolic Reprogramming in Cancer Cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef] [Green Version]
- Peck, B.; Schulze, A. Lipid Desaturation—The next Step in Targeting Lipogenesis in Cancer? FEBS J. 2016, 283, 2767–2778. [Google Scholar] [CrossRef] [PubMed]
- Al Jitan, S.; Alkhoori, S.A.; Yousef, L.F. Phenolic Acids from Plants: Extraction and Application to Human Health. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 58, pp. 389–417. ISBN 978-0-444-64056-7. [Google Scholar]
- Dreher, D.; Junod, A.F. Role of Oxygen Free Radicals in Cancer Development. Eur. J. Cancer 1996, 32, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Ziech, D.; Franco, R.; Georgakilas, A.G.; Georgakila, S.; Malamou-Mitsi, V.; Schoneveld, O.; Pappa, A.; Panayiotidis, M.I. The Role of Reactive Oxygen Species and Oxidative Stress in Environmental Carcinogenesis and Biomarker Development. Chem.-Biol. Interact. 2010, 188, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Lawenda, B.D.; Kelly, K.M.; Ladas, E.J.; Sagar, S.M.; Vickers, A.; Blumberg, J.B. Should Supplemental Antioxidant Administration Be Avoided During Chemotherapy and Radiation Therapy? J. Natl. Cancer Inst. 2008, 100, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Sayin, V.I.; Ibrahim, M.X.; Larsson, E.; Nilsson, J.A.; Lindahl, P.; Bergo, M.O. Antioxidants Accelerate Lung Cancer Progression in Mice. Sci. Transl. Med. 2014, 6, 221ra15. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hata, K.; Ishikawa, K.; Hori, K.; Konishi, T. Differentiation-Inducing Activity of Lupeol, a Lupane-Type Triterpene from Chinese Dandelion Root (Hokouei-Kon), on a Mouse Melanoma Cell Line. Biol. Pharm. Bull. 2000, 23, 962–967. [Google Scholar] [CrossRef] [Green Version]
- Hata, K. Role of P38 MAPK in Lupeol-Induced B16 2F2 Mouse Melanoma Cell Differentiation. J. Biochem. 2003, 134, 441–445. [Google Scholar] [CrossRef]
- Carvalho, K.; de Melo, T.; de Melo, K.; Quinderé, A.; de Oliveira, F.; Viana, A.; Nunes, P.; Quetz, J.; Viana, D.; da Silva, A.; et al. Amyrins from Protium Heptaphyllum Reduce High-Fat Diet-Induced Obesity in Mice via Modulation of Enzymatic, Hormonal And Inflammatory Responses. Planta Med. 2016, 83, 285–291. [Google Scholar] [CrossRef]
- Singh, A.B.; Yadav, D.K.; Maurya, R.; Srivastava, A.K. Antihyperglycaemic Activity of α-Amyrin Acetate in Rats and Db/Db Mice. Nat. Prod. Res. 2009, 23, 876–882. [Google Scholar] [CrossRef]
- Liu, X.; Su, K.; Sun, X.; Jiang, Y.; Wang, L.; Hu, C.; Zhang, C.; Lu, M.; Du, X.; Xing, B. Sec62 Promotes Stemness and Chemoresistance of Human Colorectal Cancer through Activating Wnt/β-Catenin Pathway. J. Exp. Clin. Cancer Res. 2021, 40, 132. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Li, Y.; Ren, P.; Zhang, C.; Wang, L.; Du, X.; Xing, B. MiR-6716-5p Promotes Metastasis of Colorectal Cancer through Downregulating NAT10 Expression. Cancer Manag. Res. 2019, 11, 5317–5332. [Google Scholar] [CrossRef] [Green Version]
- Radišauskas, R.; Kuzmickienė, I.; Milinavičienė, E.; Everatt, R. Hypertension, Serum Lipids and Cancer Risk: A Review of Epidemiological Evidence. Medicina 2016, 52, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Murai, T. Cholesterol Lowering: Role in Cancer Prevention and Treatment. Biol. Chem. 2015, 396, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouzas, A.; Gómez de Cedrón, M.; Colmenarejo, G.; Laparra-Llopis, J.M.; Moreno-Rubio, J.; Montoya, J.J.; Reglero, G.; Casado, E.; Tabares, B.; Sereno, M.; et al. Phenolic Diterpenes from Rosemary Supercritical Extract Inhibit Non-Small Cell Lung Cancer Lipid Metabolism and Synergise with Therapeutic Drugs in the Clinic. Front. Oncol. 2022, 12, 1046369. [Google Scholar] [CrossRef] [PubMed]
- Gómez de Cedrón, M.; Laparra, J.M.; Loria-Kohen, V.; Molina, S.; Moreno-Rubio, J.; Montoya, J.J.; Torres, C.; Casado, E.; Reglero, G.; Ramírez de Molina, A. Tolerability and Safety of a Nutritional Supplement with Potential as Adjuvant in Colorectal Cancer Therapy: A Randomized Trial in Healthy Volunteers. Nutrients 2019, 11, 2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalva, M.; Jaime, L.; Aguado, E.; Nieto, J.A.; Reglero, G.; Santoyo, S. Anti-Inflammatory and Antioxidant Activities from the Basolateral Fraction of Caco-2 Cells Exposed to a Rosmarinic Acid Enriched Extract. J. Agric. Food Chem. 2018, 66, 1167–1174. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. ISBN 978-0-12-182200-2. [Google Scholar]
- Navarro del Hierro, J.; Reglero, G.; Martin, D. Chemical Characterization and Bioaccessibility of Bioactive Compounds from Saponin-Rich Extracts and Their Acid-Hydrolysates Obtained from Fenugreek and Quinoa. Foods 2020, 9, 1159. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Herrera, T.; Navarro del Hierro, J.; Fornari, T.; Reglero, G.; Martin, D. Inhibitory Effect of Quinoa and Fenugreek Extracts on Pancreatic Lipase and α-Amylase under in Vitro Traditional Conditions or Intestinal Simulated Conditions. Food Chem. 2019, 270, 509–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro Del Hierro, J.; Casado-Hidalgo, G.; Reglero, G.; Martin, D. The Hydrolysis of Saponin-Rich Extracts from Fenugreek and Quinoa Improves Their Pancreatic Lipase Inhibitory Activity and Hypocholesterolemic Effect. Food Chem. 2021, 338, 128113. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Source | Pressure (Bar) | T (°C) | CO2 (g/min) | EtOH (%) | Yield (%) |
|---|---|---|---|---|---|---|
| S1 | Seed | 200 | 40 | 70 | 0 | 3.48 |
| S2 | Seed | 200 | 40 | 70 | 7 | 4.20 |
| S3 | Seed | 200 | 40 | 70 | 14 | 4.80 |
| PS | Pulp + skin | 200 | 40 | 70 | 7 | 4.35 |
| Extract | TPC(mg GAE/g Extract) | ABTS (mmol Trolox/g Extract) | DPPH (mmol Trolox/g Extract) |
|---|---|---|---|
| S1 | 13.70 ± 0.75 | 0.006 ± 0.000 | 0.000 ± 0.000 |
| S2 | 14.33 ± 0.57 | 0.029 ± 0.032 | 0.005 ± 0.000 |
| S3 | 16.43 ± 0.10 | 0.033 ± 0.001 | 0.009 ± 0.000 |
| PS | 11.83 ± 0.28 | 0.046 ± 0.002 | 0.021 ± 0.001 |
| Extract | Rt (min) | Compound | mg Compound/g Extract |
|---|---|---|---|
| PS | 10.019 | Gallic acid | 0.05 ± 0.04 |
| 10.739 | Gallic acid isomer | 0.14 ± 0.00 | |
| 11.995 | Benzoic acid n.i. | 0.02 ± 0.01 | |
| 12.123 | Benzoic acid n.i. | 0.04 ± 0.00 | |
| 15.556 | Flavonol n.i. | 1.52 ± 1.21 | |
| 17.476 | Flavonol n.i. | 1.46 ± 1.15 | |
| 17.675 | Anthocyanin n.i. | 0.17 ± 0.11 | |
| 18.226 | Flavonol n.i. | 2.06 ± 1.19 | |
| 18.521 | Flavonol n.i. | 1.55 ± 1.20 | |
| 22.372 | Quercetin derivative. | 0.02 ± 0.02 | |
| 22.771 | Hyperoxide | 0.02 ± 0.05 | |
| 28.645 | Caffeic acid derivative | 0.20 ± 0.10 | |
| 33.656 | Flavonol n.i. | 0.05 ± 0.01 |
| Extract | EC50 (μg/mL) |
|---|---|
| S1 | 244.69 ± 5.70 |
| S2 | >2000 |
| S3 | 1092.33 ± 5.31 |
| PS | 303.53 ± 3.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, S.; Gómez de Cedrón, M.; Navarro del Hierro, J.; Martín-Hernández, D.; Siles, M.d.l.N.; Santoyo, S.; Jaime, L.; Martín, D.; Fornari, T.; Ramírez de Molina, A. Biological Activities of Miracle Berry Supercritical Extracts as Metabolic Regulators in Chronic Diseases. Int. J. Mol. Sci. 2023, 24, 6957. https://doi.org/10.3390/ijms24086957
Wagner S, Gómez de Cedrón M, Navarro del Hierro J, Martín-Hernández D, Siles MdlN, Santoyo S, Jaime L, Martín D, Fornari T, Ramírez de Molina A. Biological Activities of Miracle Berry Supercritical Extracts as Metabolic Regulators in Chronic Diseases. International Journal of Molecular Sciences. 2023; 24(8):6957. https://doi.org/10.3390/ijms24086957
Chicago/Turabian StyleWagner, Sonia, Marta Gómez de Cedrón, Joaquín Navarro del Hierro, Diego Martín-Hernández, María de las Nieves Siles, Susana Santoyo, Laura Jaime, Diana Martín, Tiziana Fornari, and Ana Ramírez de Molina. 2023. "Biological Activities of Miracle Berry Supercritical Extracts as Metabolic Regulators in Chronic Diseases" International Journal of Molecular Sciences 24, no. 8: 6957. https://doi.org/10.3390/ijms24086957