
Unravelling the Effects of Syndecan-4 Knockdown on Skeletal Muscle Functions
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. In Vivo Experiments
Decreased Grip Force and Voluntary Running in SDC4 Mice
2.2. In Vitro Experiments
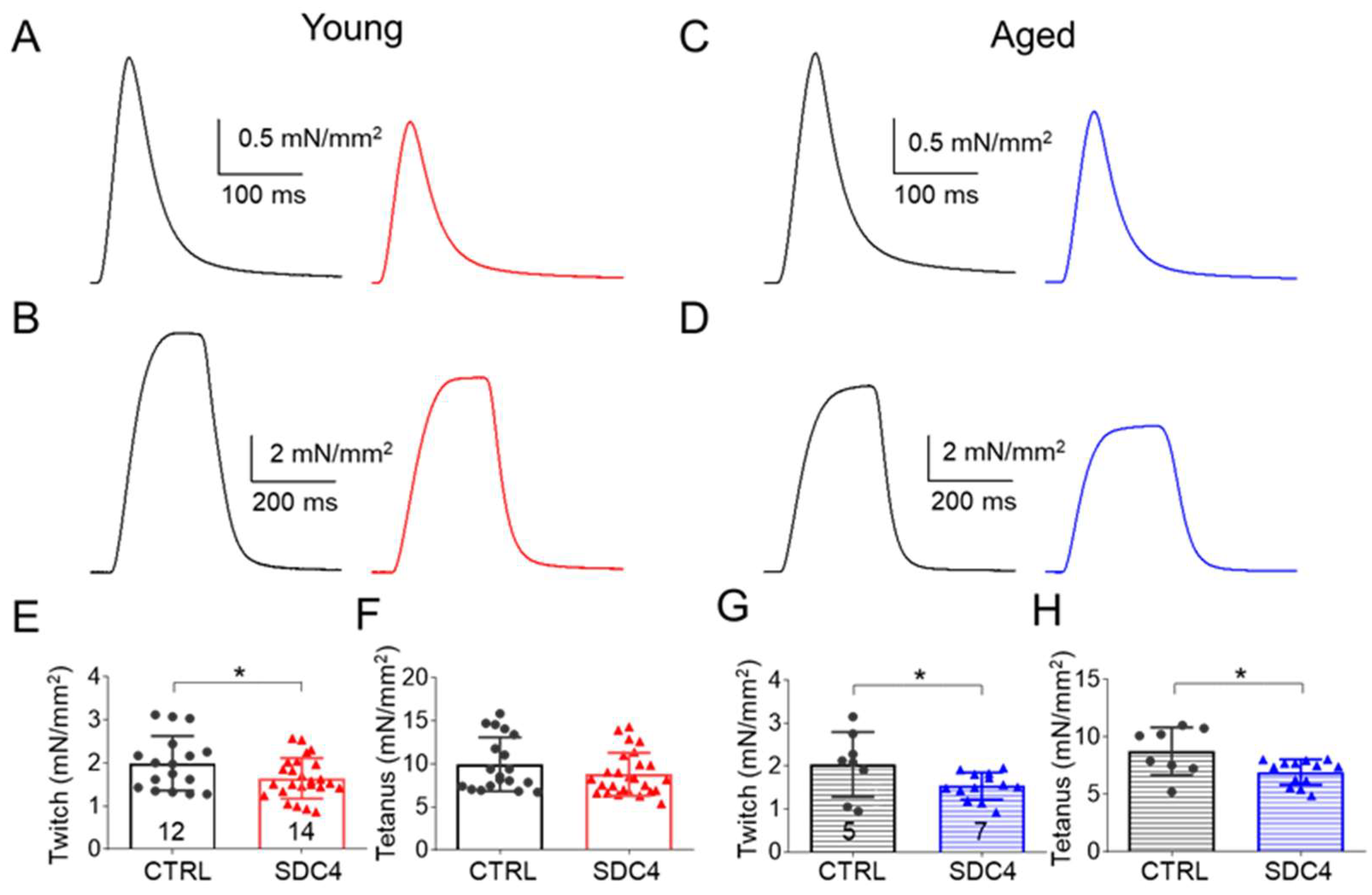
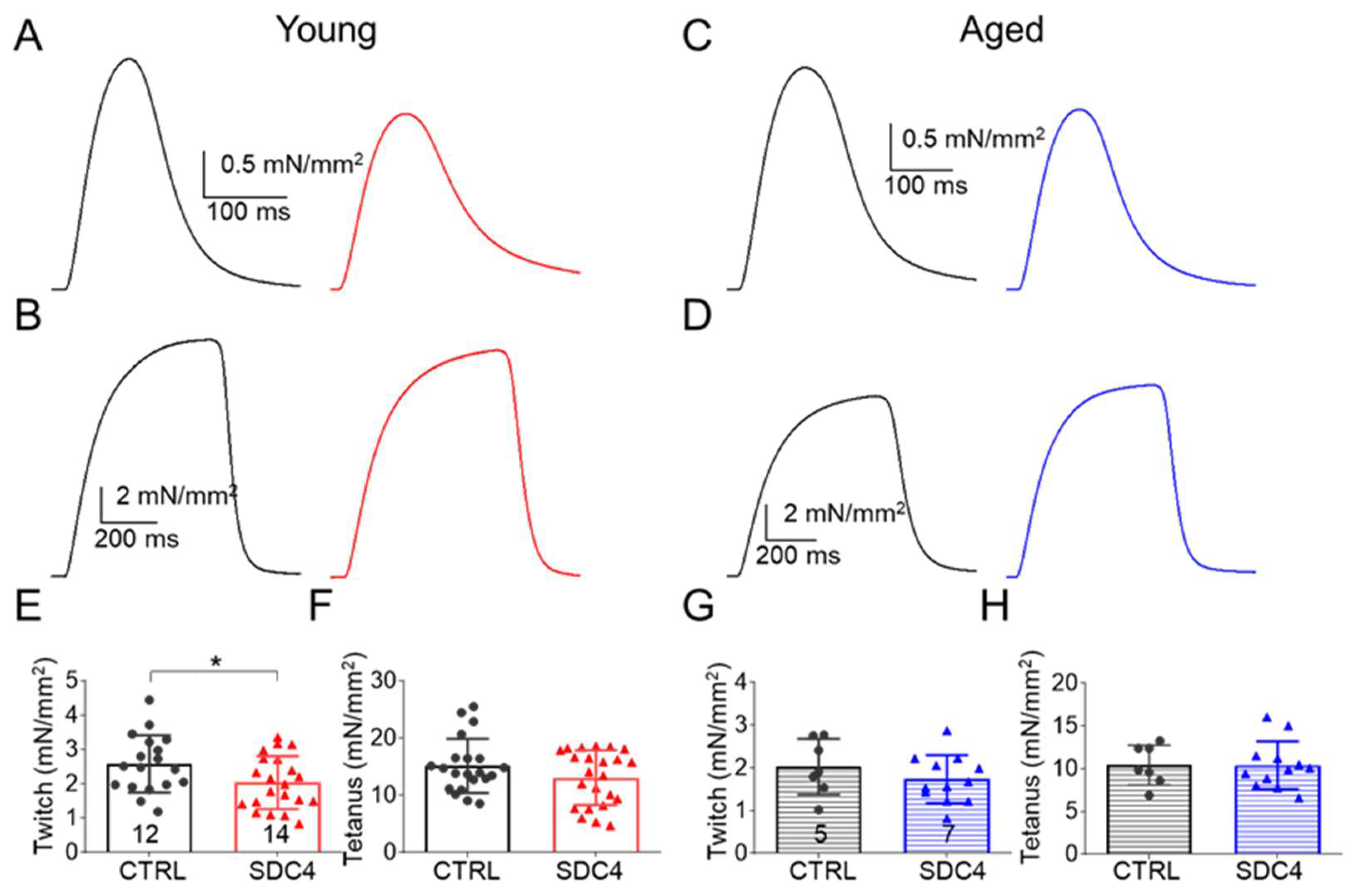
2.2.1. Decreased Twitch Force in SDC4 Mice
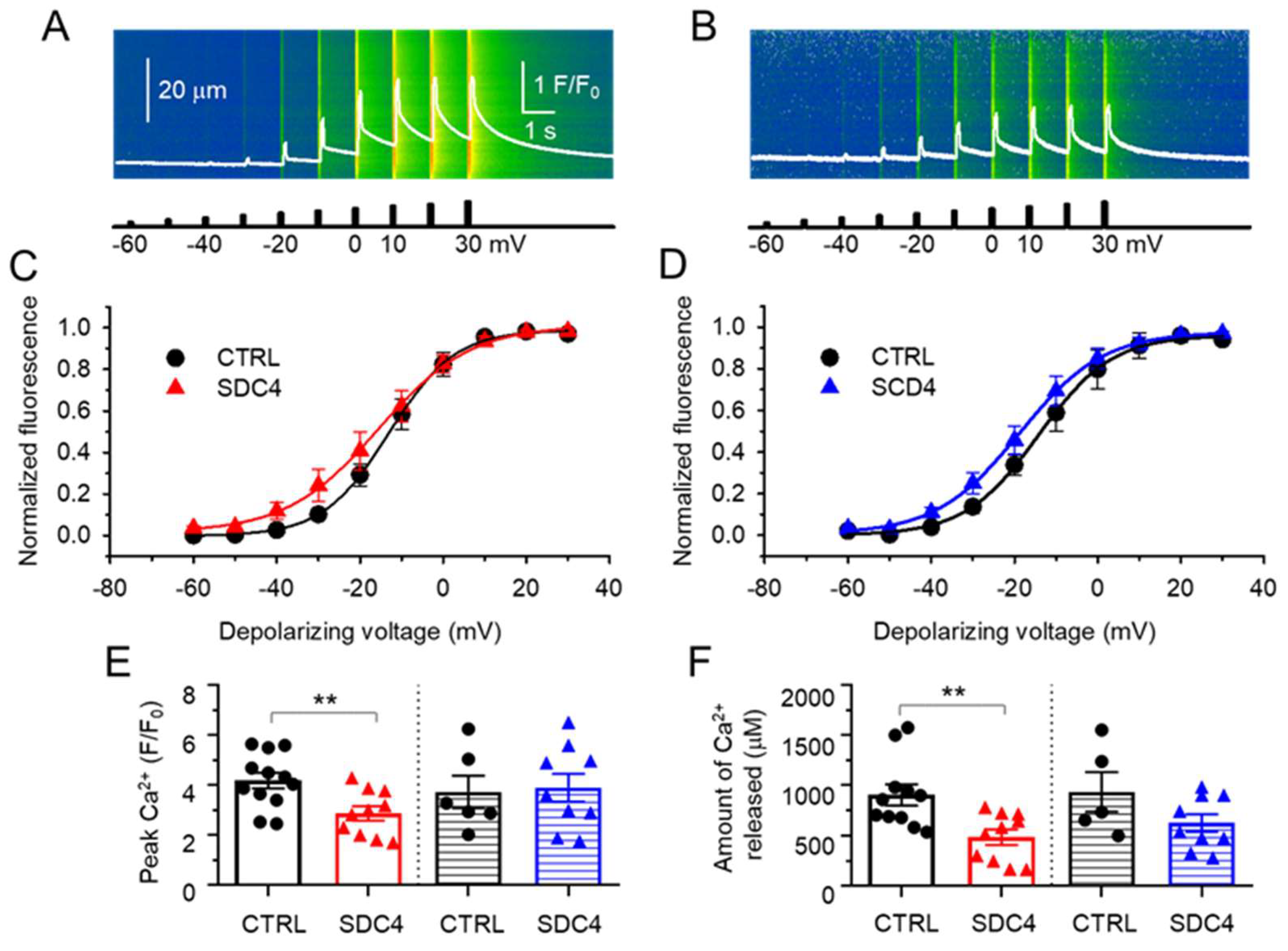
2.2.2. Suppressed SR Ca2+ Release with Unaltered Voltage Dependence in SDC4 Mice
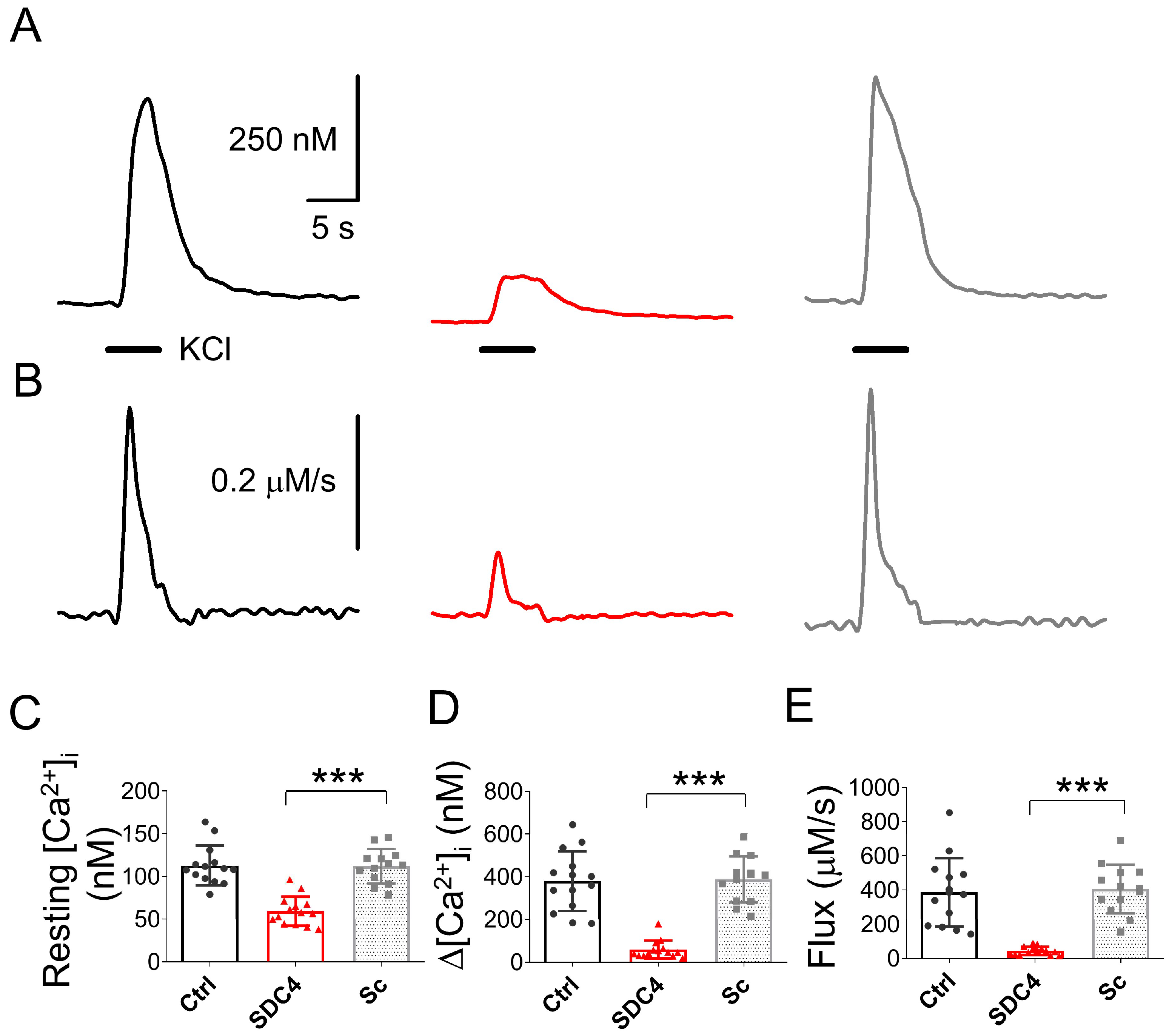
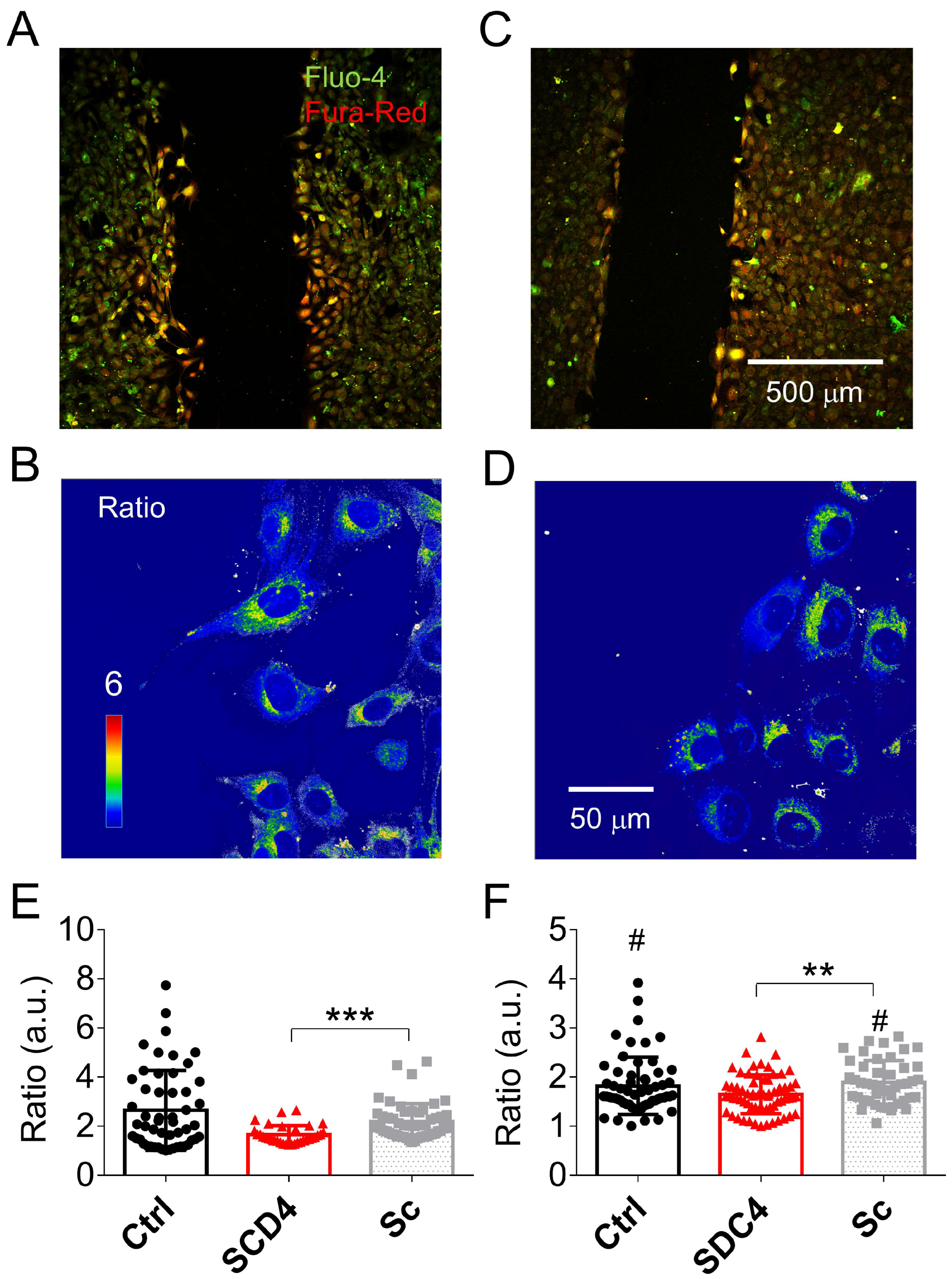
2.2.3. Altered Calcium Homeostasis in Syndecan-4-Silenced C2C12 Cells
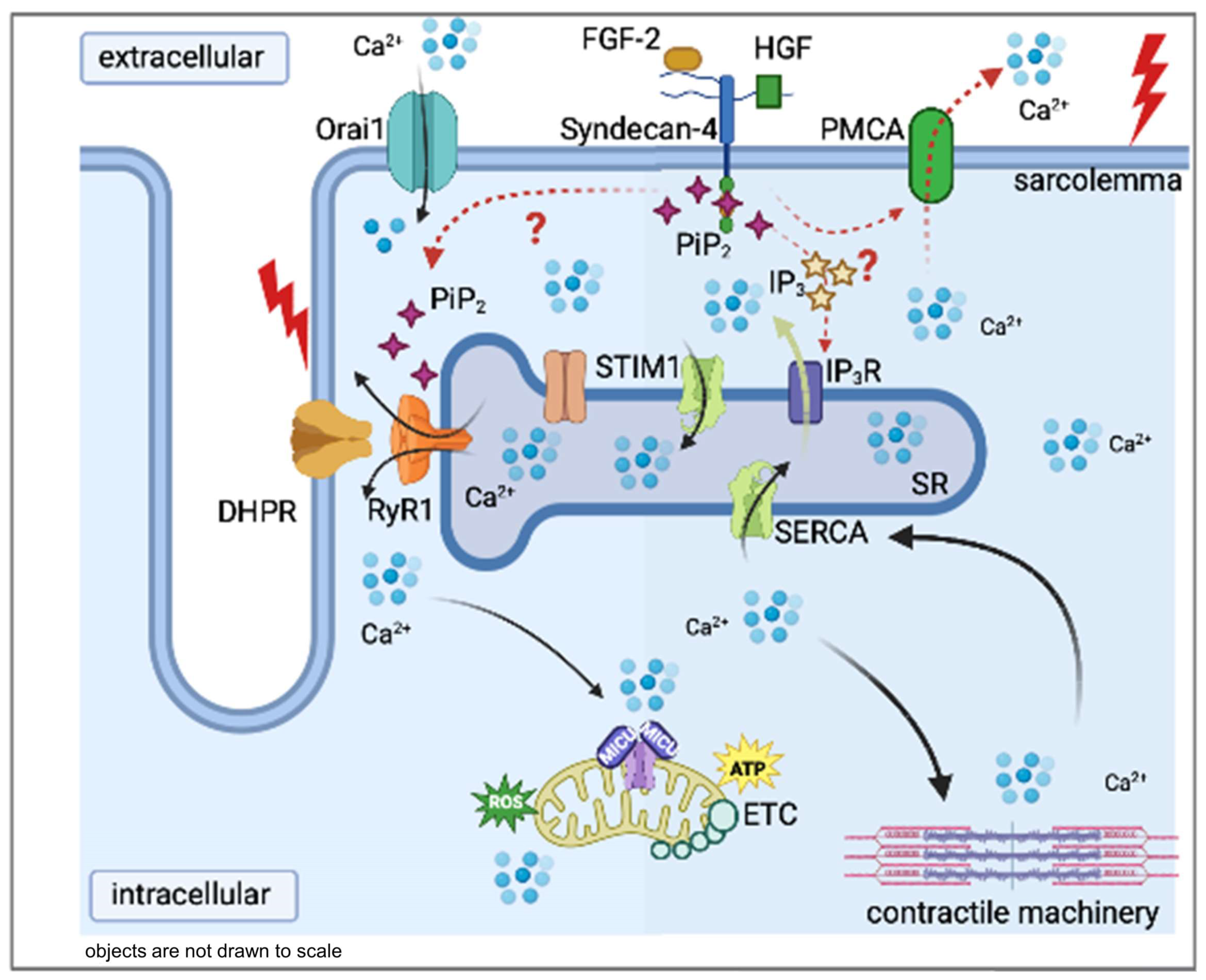
3. Discussion
4. Materials and Methods
4.1. Animal Care and Genotyping
4.2. In Vivo Experiments
4.2.1. Voluntary Activity Wheel Measurement
4.2.2. Forepaw Grip Test
4.3. In Vitro Experiments
4.3.1. Measurement of Muscle Force
4.3.2. Isolation of Single Skeletal Muscle Fibres
4.3.3. Voltage Clamp and Calculation of Intracellular Ca2+ Concentration
4.3.4. Confocal Microscopy and Image Analysis
4.3.5. Cell Culture and Plasmids
4.3.6. Whole-Cell Calcium Measurements in Cell Culture
4.3.7. Calcium Concentration Measurements during Migration
4.3.8. Chemicals and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flück, M.; Hoppeler, H. Molecular basis of skeletal muscle plasticity—From gene to form and function. Rev. Physiol. Biochem. Pharmacol. 2003, 146, 159–216. [Google Scholar] [CrossRef]
- Thomas, K.; Engler, A.J.; Meyer, G.A. Extracellular matrix regulation in the muscle satellite cell niche. Connect. Tissue Res. 2015, 56, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Xian, X.; Gopal, S.; Couchman, J.R. Syndecans as receptors and organizers of the extracellular matrix. Cell Tissue Res. 2010, 339, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Cornelison, D.D.W.; Filla, M.S.; Stanley, H.M.; Rapraeger, A.C.; Olwin, B.B. Syndecan-3 and syndecan-4 specifically mark skeletal muscle satellite cells and are implicated in satellite cell maintenance and muscle regeneration. Dev. Biol. 2001, 239, 79–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; McFarland, D.C.; Nestor, K.E.; Velleman, S.G. Differential expression of membrane-associated heparan sulfate proteoglycans in the skeletal muscle of turkeys with different growth rates. Poult. Sci. 2006, 85, 422–428. [Google Scholar] [CrossRef]
- Velleman, S.G.; Song, Y. Development and growth of the avian pectoralis major (Breast) muscle: Function of syndecan-4 and glypican-1 in adult myoblast proliferation and differentiation. Front. Physiol. 2017, 8, 577. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Echtermeyer, F.; Thilo, F.; Theilmeier, G.; Schmidt, A.; Schülein, R.; Jensen, B.L.; Loddenkemper, C.; Jankowski, V.; Marcussen, N.; et al. The proteoglycan syndecan 4 regulates transient receptor potential canonical 6 channels via RhoA/rho-associated protein kinase signaling. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopal, S.; Søgaard, P.; Multhaupt, H.A.B.; Pataki, C.; Okina, E.; Xian, X.; Pedersen, M.E.; Stevens, T.; Griesbeck, O.; Park, P.W.; et al. Transmembrane proteoglycans control stretch-activated channels to set cytosolic calcium levels. J. Cell Biol. 2015, 210, 1199–1211. [Google Scholar] [CrossRef] [Green Version]
- Becsky, D.; Szabo, K.; Gyulai-Nagy, S.; Gajdos, T.; Bartos, Z.; Balind, A.; Dux, L.; Horvath, P.; Erdelyi, M.; Homolya, L.; et al. Syndecan-4 Modulates Cell Polarity and Migration by Influencing Centrosome Positioning and Intracellular Calcium Distribution. Front. Cell Dev. Biol. 2020, 8, 575227. [Google Scholar] [CrossRef]
- Ugarte, G.; Santander, C.; Brandan, E. Syndecan-4 and β1 integrin are regulated by electrical activity in skeletal muscle: Implications for cell adhesion. Matrix Biol. 2010, 29, 383–392. [Google Scholar] [CrossRef]
- Olguin, H.; Brandan, E. Expression and localization of proteoglycans during limb myogenic activation. Dev. Dyn. 2001, 221, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Do, M.K.Q.; Shimizu, N.; Suzuki, T.; Ohtsubo, H.; Mizunoya, W.; Nakamura, M.; Sawano, S.; Furuse, M.; Ikeuchi, Y.; Anderson, J.E.; et al. Transmembrane proteoglycans syndecan-2, 4, receptor candidates for the impact of HGF and FGF2 on semaphorin 3A expression in early-differentiated myoblasts. Physiol. Rep. 2015, 3, e12553. [Google Scholar] [CrossRef] [PubMed]
- Brandan, E.; Gutierrez, J. Role of skeletal muscle proteoglycans during myogenesis. Matrix Biol. 2013, 32, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Varga, D.; Vegh, A.G.; Liu, N.; Xiao, X.; Xu, L.; Dux, L.; Erdelyi, M.; Rovo, L.; Keller-Pinter, A. Syndecan-4 affects myogenesis via Rac1-mediated actin remodeling and exhibits copy-number amplification and increased expression in human rhabdomyosarcoma tumors. Cell. Mol. Life Sci. 2022, 79, 122. [Google Scholar] [CrossRef] [PubMed]
- Cornelison, D.D.W.; Wilcox-Adelman, S.A.; Goetinck, P.F.; Rauvala, H.; Rapraeger, A.C.; Olwin, B.B. Essential and separable roles for Syndecan-3 and Syndecan-4 in skeletal muscle development and regeneration. Genes Dev. 2004, 18, 2231–2236. [Google Scholar] [CrossRef] [Green Version]
- Rønning, S.B.; Carlson, C.R.; Aronsen, J.M.; Pisconti, A.; Høst, V.; Lunde, M.; Liland, K.H.; Sjaastad, I.; Kolset, S.O.; Christensen, G.; et al. Syndecan-4–/– Mice Have Smaller Muscle Fibers, Increased Akt/mTOR/S6K1 and Notch/HES-1 Pathways, and Alterations in Extracellular Matrix Components. Front. Cell Dev. Biol. 2020, 8, 730. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C.; Jorgensen, A.O. Structure and development of E-C coupling units in skeletal muscle. Annu. Rev. Physiol. 1994, 56, 509–534. [Google Scholar] [CrossRef]
- Meissner, G.; Lu, X. Dihydropyridine receptor-ryanodine receptor interactions in skeletal muscle excitation-contraction coupling. Biosci. Rep. 1995, 15, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Osses, N.; Brandan, E. Changes in secreted and cell associated proteoglycan synthesis during conversion of myoblasts to osteoblasts in response to bone morphogenetic protein-2: Role of decorin in cell response to BMP-2. J. Cell. Physiol. 2006, 206, 58–67. [Google Scholar] [CrossRef]
- Melzer, W.; Rios, E.; Schneider, M.F. Time course of calcium release and removal in skeletal muscle fibers. Biophys. J. 1984, 45, 637–641. [Google Scholar] [CrossRef] [Green Version]
- Schuhmeier, R.P.; Melzer, W. Voltage-dependent Ca2+ Fluxes in Skeletal Myotubes Determined Using a Removal Model Analysis. J. Gen. Physiol. 2004, 123, 33–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller-Pinter, A.; Szabo, K.; Kocsis, T.; Deak, F.; Ocsovszki, I.; Zvara, A.; Puskas, L.; Szilak, L.; Dux, L. Syndecan-4 influences mammalian myoblast proliferation by modulating myostatin signalling and G1/S transition. FEBS Lett. 2018, 592, 3139–3151. [Google Scholar] [CrossRef]
- Greene, D.K.; Tumova, S.; Couchman, J.R.; Woods, A. Syndecan-4 associates with α-actinin. J. Biol. Chem. 2003, 278, 7617–7623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, R.; Moreno, M.; Oliva, C.; Orbenes, C.; Larraín, J. Syndecan-4 regulates non-canonical wnt signalling and is essential for convergent and extension movements in xenopus embryos. Nat. Cell Biol. 2006, 8, 492–500. [Google Scholar] [CrossRef]
- Bertrand, J.; Stange, R.; Hidding, H.; Echtermeyer, F.; Nalesso, G.; Godmann, L.; Timmen, M.; Bruckner, P.; Dell’Accio, F.; Raschke, M.J.; et al. Syndecan 4 supports bone fracture repair, but not fetal skeletal development, in mice. Arthritis Rheum. 2013, 65, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.E.; Herum, K.M.; Rana, Z.A.; Skrbic, B.; Askevold, E.T.; Dahl, C.P.; Vistnes, M.; Hasic, A.; Kvaløy, H.; Sjaastad, I.; et al. Innate immune signaling induces expression and shedding of the heparan sulfate proteoglycan syndecan-4 in cardiac fibroblasts and myocytes, affecting inflammation in the pressure-overloaded heart. FEBS J. 2013, 280, 2228–2247. [Google Scholar] [CrossRef] [Green Version]
- Gondelaud, F.; Ricard-Blum, S. Structures and interactions of syndecans. FEBS J. 2019, 286, 2994–3007. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Oh, E.S.; Woods, A.; Couchman, J.R.; Lee, W. Solution structure of a syndecan-4 cytoplasmic domain and its interaction with phosphatidylinositol 4,5-bisphosphate. J. Biol. Chem. 1998, 273, 13022–13029. [Google Scholar] [CrossRef] [Green Version]
- González Rodríguez, E.; Lefebvre, R.; Bodnár, D.; Legrand, C.; Szentesi, P.; Vincze, J.; Poulard, K.; Bertrand-Michel, J.; Csernoch, L.; Buj-Bello, A.; et al. Phosphoinositide substrates of myotubularin affect voltage-activated Ca2+ release in skeletal muscle. Pflugers Arch. Eur. J. Physiol. 2014, 466, 973–985. [Google Scholar] [CrossRef]
- Araya, R.; Liberona, J.L.; Cárdenas, J.C.; Riveros, N.; Estrada, M.; Powell, J.A.; Carrasco, M.A.; Jaimovich, E. Dihydropyridine receptors as voltage sensors for a depolarization-evoked, IP3R-mediated, slow calcium signal in skeletal muscle cells. J. Gen. Physiol. 2003, 121, 3–16. [Google Scholar] [CrossRef] [Green Version]
- de Luca, M.; Klimentidis, Y.C.; Casazza, K.; Chambers, M.M.; Cho, R.; Harbison, S.T.; Jumbo-Lucioni, P.; Zhang, S.; Leips, J.; Fernandez, J.R. A conserved role for syndecan family members in the regulation of whole-body energy metabolism. PLoS ONE 2010, 5, e11286. [Google Scholar] [CrossRef]
- Eveland, M.; Brokamp, G.A.; Lue, C.H.; Harbison, S.T.; Leips, J.; De Luca, M. Knockdown expression of Syndecan in the fat body impacts nutrient metabolism and the organismal response to environmental stresses in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2016, 477, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Ishiguro, K.; Kojima, T.; Saito, H.; Muramatsu, T.; Kadomatsu, K.; Taguchi, O. Syndecan-4 expression is associated with follicular atresia in mouse ovary. Histochem. Cell Biol. 1999, 112, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Singlár, Z.; Ganbat, N.; Szentesi, P.; Osgonsandag, N.; Szabó, L.; Telek, A.; Fodor, J.; Dienes, B.; Gönczi, M.; Csernoch, L.; et al. Genetic Manipulation of CB1 Cannabinoid Receptors Reveals a Role in Maintaining Proper Skeletal Muscle Morphology and Function in Mice. Int. J. Mol. Sci. 2022, 23, 15653. [Google Scholar] [CrossRef] [PubMed]
- Szentesi, P.; Jacquemond, V.; Kovács, L.; Csernoch, L. Intramembrane charge movement and sarcoplasmic calcium release in enzymatically isolated mammalian skeletal muscle fibres. J. Physiol. 1997, 505, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.; Gönczi, M.; Sztretye, M.; Dienes, B.; Oláh, T.; Szabó, L.; Csoma, E.; Szentesi, P.; Szigeti, G.P.; Marty, I.; et al. Altered expression of triadin 95 causes parallel changes in localized Ca2+ release events and global Ca2+ signals in skeletal muscle cells in culture. J. Physiol. 2008, 586, 5803–5818. [Google Scholar] [CrossRef] [PubMed]
- Sztretye, M.; Geyer, N.; Vincze, J.; Al-Gaadi, D.; Oláh, T.; Szentesi, P.; Kis, G.; Antal, M.; Balatoni, I.; Csernoch, L.; et al. SOCE Is Important for Maintaining Sarcoplasmic Calcium Content and Release in Skeletal Muscle Fibers. Biophys. J. 2017, 113, 2496–2507. [Google Scholar] [CrossRef] [Green Version]
- Royer, L.; Pouvreau, S.; Ríos, E. Evolution and modulation of intracellular calcium release during long-lasting, depleting depolarization in mouse muscle. J. Physiol. 2008, 586, 4609–4629. [Google Scholar] [CrossRef]
- Szappanos, H.; Cseri, J.; Deli, T.; Kovács, L.; Csernoch, L. Determination of depolarisation- and agonist-evoked calcium fluxes on skeletal muscle cells in primary culture. J. Biochem. Biophys. Methods 2004, 59, 89–101. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young | Aged | |||
|---|---|---|---|---|
| CTRL | SDC4 | CTRL | SDC4 | |
| Body weight (g) | 26.7 ± 1.1 | 24.9 ± 0.9 | 32.8 ± 0.8 | 30.1 ± 0.5 ** |
| Maximal force (mN) | 148.8 ± 7.2 | 126.1 ± 3.8 ** | 154.3 ± 7.3 | 134.1 ± 4.5 * |
| Force normalized to body weight (mN/g) | 55.8 ± 1.5 | 51.1 ± 1.2 * | 50.9 ± 2.3 | 43.8 ± 2.0 * |
| Number of mice | 9 | 12 | 11 | 18 |
| Gender distribution | 4 male, 5 female | 8 male, 4 female | 11 male | 18 male |
| Young | Aged | |||
|---|---|---|---|---|
| CTRL | SDC4 | CTRL | SDC4 | |
| Distance (m/day) | 7335.7 ± 155.6 | 7571.1 ± 279.6 | 3203.7 ± 206.8 | 2231.7 ± 177.0 ** |
| Average speed (m/min) | 14.3 ± 0.2 | 11.5 ± 0.5 *** | 5.2 ± 0.3 | 4.2 ± 0.2 * |
| Max speed (m/min) | 27.4 ± 0.2 | 23.2 ± 1.1 ** | 12.3 ± 0.4 | 9.7 ± 0.5 ** |
| Time (min/day) | 516 ± 6 | 631 ± 7 *** | 326 ± 10 | 270 ± 14 ** |
| Number of mice | 4 | 5 | 4 | 4 |
| Young | Aged | |||||||
|---|---|---|---|---|---|---|---|---|
| Twitch | Tetanus | Twitch | Tetanus | |||||
| CTRL | SDC4 | CTRL | SDC4 | CTRL | SDC4 | CTRL | SDC4 | |
| Number of muscles | 17 | 18 | 17 | 18 | 7 | 12 | 7 | 12 |
| Muscle weight (mg) | 15.5 ± 0.5 | 14.4 ± 0.3 * | 15.5 ± 0.5 | 14.4 ± 0.3 * | 13.9 ± 0.4 | 17.1 ± 0.3 *** | 13.9 ± 0.4 | 17.1 ± 0.3 *** |
| Peak force (mN) | 2.04 ± 0.10 | 1.77 ± 0.08 * | 10.16 ± 0.35 | 9.34 ± 0.31 | 1.74 ± 0.26 | 1.81 ± 0.13 | 7.31 ± 0.55 | 8.05 ± 0.31 |
| Force (mN/mm2) | 1.99 ± 0.15 | 1.65 ± 0.09 * | 9.96 ± 0.72 | 8.79 ± 0.49 | 2.04 ± 0.27 | 1.54 ± 0.09 * | 8.75 ± 0.73 | 6.93 ± 0.31 * |
| TTP (ms) | 32.9 ± 1.4 | 29.9 ± 0.2 | 172.2 ± 4.0 | 157.7 ± 6.7 | 38.1 ± 3.1 | 30.9 ± 1.2 * | 193.5 ± 4.6 | 192.2 ± 3.1 |
| HRT (ms) | 30.1 ± 0.8 | 26.6 ± 0.9 | 73.4 ± 4.1 | 99.5 ± 7.1 ** | 27.6 ± 1.5 | 22.9 ± 0.6 ** | 81.9 ± 7.2 | 79.3 ± 4.3 |
| Duration (ms) | 210.2 ± 29.1 | 164.4 ± 13.1 | 336.4 ± 5.4 | 338.5 ± 4.2 | 207.7 ± 25.2 | 132.0 ± 5.6 ** | 374.4 ± 12.9 | 356.5 ± 6.6 |
| CSA (mm2) | 1.07 ± 0.06 | 1.11 ± 0.05 | 1.07 ± 0.06 | 1.11 ± 0.05 | 0.84 ± 0.06 | 1.18 ± 0.07 ** | 0.84 ± 0.06 | 1.18 ± 0.07 ** |
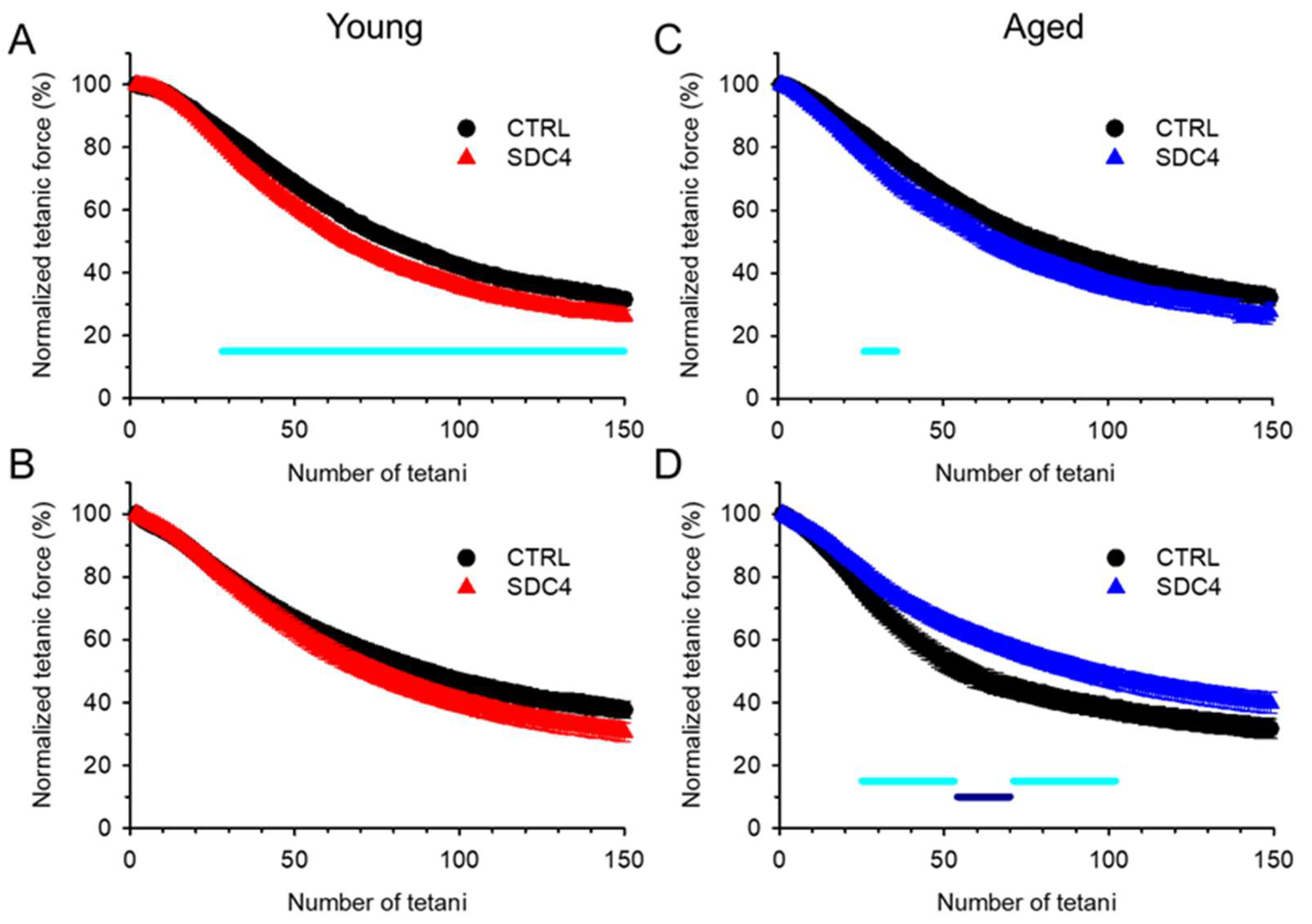
| Fatigue at 50 (%) | 31.6 ± 1.7 | 38.7 ± 2.2 * | 33.3 ± 1.6 | 40.5 ± 3.1 | ||||
| Fatigue at 100 (%) | 57.7 ± 1.8 | 64.8 ± 2.0 * | 57.0 ± 2.7 | 63.6 ± 2.9 | ||||
| Fatigue at 150 (%) | 68.4 ± 1.9 | 74.1 ± 1.8 * | 67.9 ± 2.6 | 72.2 ± 2.7 | ||||
| Young | Aged | |||||||
|---|---|---|---|---|---|---|---|---|
| Twitch | Tetanus | Twitch | Tetanus | |||||
| CTRL | SDC4 | CTRL | SDC4 | CTRL | SDC4 | CTRL | SDC4 | |
| Number of muscles | 18 | 18 | 18 | 18 | 7 | 12 | 7 | 12 |
| Muscle weight (mg) | 16.3 ± 0.7 | 14.5 ± 0.2 ** | 16.3 ± 0.7 | 14.5 ± 0.2 ** | 14.0 ± 0.5 | 16.8 ± 0.6 ** | 14.0 ± 0.5 | 16.8 ± 0.6 ** |
| Peak force (mN) | 2.17 ± 0.12 | 1.76 ± 0.11 * | 12.93 ± 0.30 | 11.33 ± 0.58 * | 1.70 ± 0.15 | 1.77 ± 0.13 | 8.85 ± 0.56 | 10.17 ± 0.48 |
| Force (mN/mm2) | 2.58 ± 0.20 | 2.03 ± 0.17 * | 15.12 ± 1.03 | 13.05 ± 1.00 | 2.03 ± 0.25 | 1.73 ± 0.16 | 10.42 ± 0.88 | 10.41 ± 0.81 |
| TTP (ms) | 74.1 ± 3.5 | 70.3 ± 2.4 | 507.5 ± 1.9 | 520.8 ± 1.9 *** | 102.9 ± 13.3 | 70.4 ± 2.7 ** | 535.7 ± 5.8 | 521.0 ± 1.5 ** |
| HRT (ms) | 72.1 ± 5.1 | 61.7 ± 3.1 * | 97.6 ± 3.9 | 95.7 ± 1.7 | 90.4 ± 12.8 | 54.6 ± 2.4 ** | 138.5 ± 7.4 | 123.6 ± 2.6 * |
| Duration (ms) | 324.3 ± 18.9 | 299.2 ± 12.0 | 720.0 ± 11.2 | 712.0 ± 5.1 | 389.2 ± 59.6 | 256.3 ± 11.9 * | 853.2 ± 26.5 | 791.7 ± 9.1 * |
| CSA (mm2) | 0.89 ± 0.06 | 0.90 ± 0.04 | 0.89 ± 0.06 | 0.90 ± 0.04 | 0.99 ± 0.09 | 1.03 ± 0.06 | 0.99 ± 0.09 | 1.03 ± 0.06 |
| Fatigue at 50 (%) | 33.6 ± 0.5 | 37.0 ± 3.0 | 46.1 ± 3.5 | 34.1 ± 2.6 * | ||||
| Fatigue at 100 (%) | 53.6 ± 0.6 | 59.8 ± 3.0 | 62.0 ± 2.9 | 51.7 ± 3.1 * | ||||
| Fatigue at 150 (%) | 62.3 ± 0.6 | 69.4 ± 2.9 | 14.0 ± 0.5 | 16.8 ± 0.6 ** | 68.3 ± 3.1 | 60.1 ± 3.3 | ||
| Young | Aged | |||
|---|---|---|---|---|
| CTRL | SDC4 | CTRL | SDC4 | |
| V50 (mV) | −12.4 ± 2.3 | −15.6 ± 3.2 | −12.3 ± 3.3 | −17.8 ± 2.5 |
| k (mV) | 6.7 ± 0.5 | 7.4 ± 1.2 | 9.2 ± 1.4 | 9.8 ± 1.0 |
| Fitted [Ca2+]i,max (F/F0) | 1.99 ± 0.22 | 1.12 ± 0.16 ** | 2.04 ± 0.62 | 2.02 ± 0.36 |
| Number of fibres | 12 | 14 | 5 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sztretye, M.; Singlár, Z.; Ganbat, N.; Al-Gaadi, D.; Szabó, K.; Köhler, Z.M.; Dux, L.; Keller-Pintér, A.; Csernoch, L.; Szentesi, P. Unravelling the Effects of Syndecan-4 Knockdown on Skeletal Muscle Functions. Int. J. Mol. Sci. 2023, 24, 6933. https://doi.org/10.3390/ijms24086933
Sztretye M, Singlár Z, Ganbat N, Al-Gaadi D, Szabó K, Köhler ZM, Dux L, Keller-Pintér A, Csernoch L, Szentesi P. Unravelling the Effects of Syndecan-4 Knockdown on Skeletal Muscle Functions. International Journal of Molecular Sciences. 2023; 24(8):6933. https://doi.org/10.3390/ijms24086933
Chicago/Turabian StyleSztretye, Mónika, Zoltán Singlár, Nyamkhuu Ganbat, Dána Al-Gaadi, Kitti Szabó, Zoltán Márton Köhler, László Dux, Anikó Keller-Pintér, László Csernoch, and Péter Szentesi. 2023. "Unravelling the Effects of Syndecan-4 Knockdown on Skeletal Muscle Functions" International Journal of Molecular Sciences 24, no. 8: 6933. https://doi.org/10.3390/ijms24086933