
Transcriptomics Reveal Molecular Differences in Equine Oocytes Vitrified before and after In Vitro Maturation
 , , ,
, , ,  , , , , and
, , , , and {kind=link}
{kind=link}
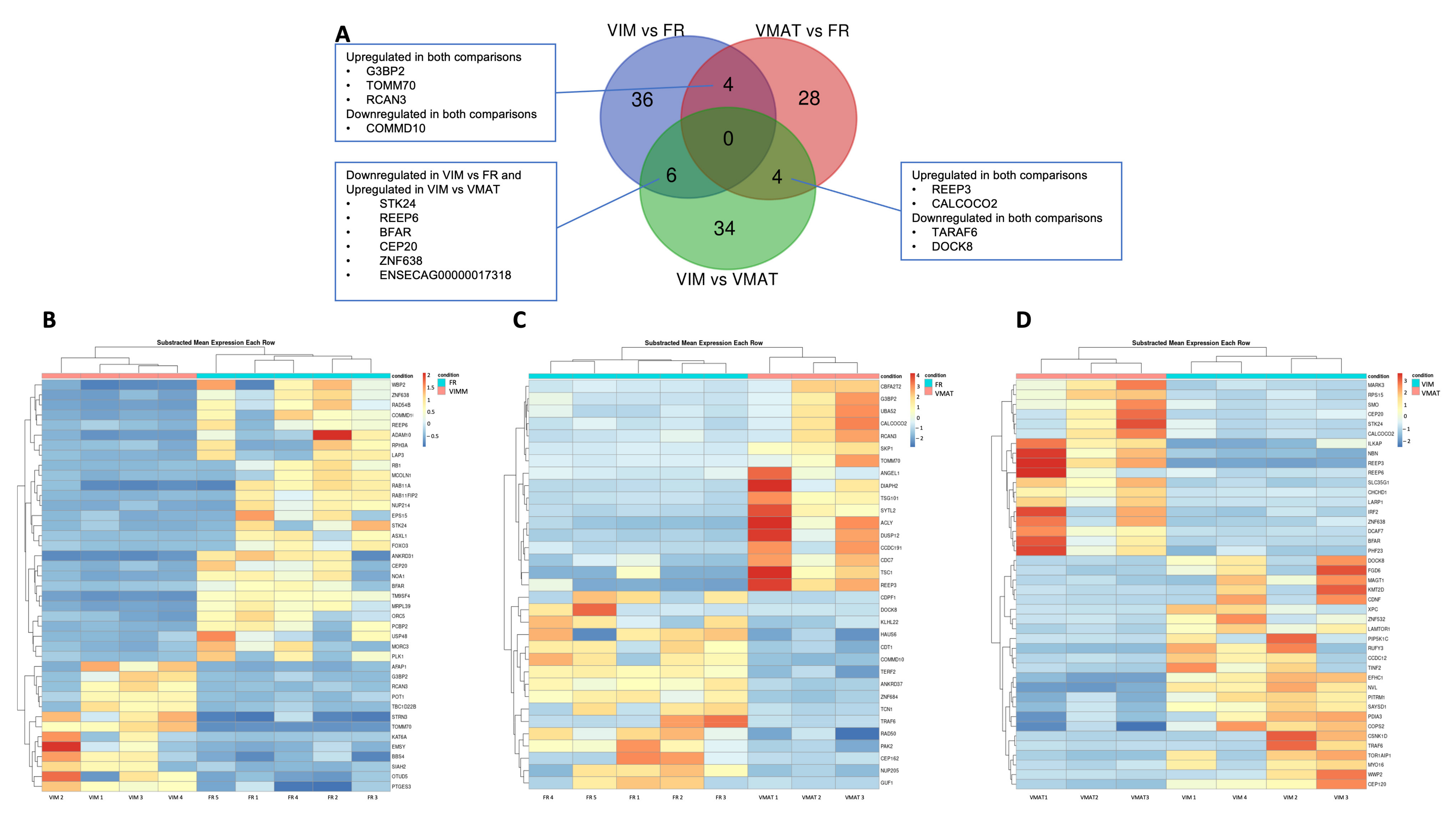{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Vitrification Impairs Oocyte Survival and Maturation
2.2. Vitrification of Immature and Mature Oocytes Affects the mRNA Profile
2.3. Vitrification Affects Pathways Involved in Oocyte Development
3. Discussion
4. Materials and Methods
4.1. Media and Reagents
4.2. Collection of Equine Immature Oocytes
4.3. Oocyte Vitrification and Warming
4.4. In Vitro Maturation
4.5. RNA Extraction and Sequencing
4.6. Differential Gene Expression Analysis
4.7. Statistical Analysis for Maturation and Survival Rates
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Coster, T.; Velez, D.A.; Van Soom, A.; Woelders, H.; Smits, K. Cryopreservation of equine oocytes: Looking into the crystal ball. Reprod. Fertil. Dev. 2020, 32, 453–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclellan, L.J.; Carnevale, E.M.; Silva, M.A.C.; Scoggin, C.F.; Bruemmer, J.E.; Squires, E.L. Pregnancies from Vitrifed Equine Oocytes Collected from Super-Stimulated and Non-Stimulated Mares. Theriogenology 2002, 58, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Escribano, N.; Bogado Pascottini, O.; Woelders, H.; Vandenberghe, L.; De Schauwer, C.; Govaere, J.; Van den Abbeel, E.; Vullers, T.; Ververs, C.; Roels, K.; et al. An Improved Vitrification Protocol for Equine Immature Oocytes, Resulting in a First Live Foal. Equine Vet. J. 2018, 50, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Tharasanit, T.; Colenbrander, B.; Stout, T. Effect of maturation stage at cryopreservation on post-thaw cytoskeleton quality and fertilizability of equine oocytes. Mol. Reprod. Dev. 2006, 73, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Canesin, H.S.; Brom-De-Luna, J.G.; Choi, Y.-H.; Pereira, A.M.; Macedo, G.G.; Hinrichs, K. Vitrification of germinal-vesicle stage equine oocytes: Effect of cryoprotectant exposure time on in-vitro embryo production. Cryobiology 2018, 81, 185–191. [Google Scholar] [CrossRef]
- Angel, D.; Canesin, H.S.; Brom-De-Luna, J.G.; Morado, S.; Dalvit, G.; Gomez, D.; Posada, N.; Pascottini, O.B.; Urrego, R.; Hinrichs, K.; et al. Embryo development after vitrification of immature and in vitro-matured equine oocytes. Cryobiology 2020, 92, 251–254. [Google Scholar] [CrossRef]
- Angel-Velez, D.; De Coster, T.; Azari-Dolatabad, N.; Fernandez-Montoro, A.; Benedetti, C.; Pascottini, O.B.; Woelders, H.; Van Soom, A.; Smits, K. New Alternative Mixtures of Cryoprotectants for Equine Immature Oocyte Vitrification. Animals 2021, 11, 3077. [Google Scholar] [CrossRef]
- Clérico, G.; Taminelli, G.; Veronesi, J.; Polola, J.; Pagura, N.; Pinto, C.; Sansinena, M. Mitochondrial function, blastocyst development and live foals born after ICSI of immature vitrified/warmed equine oocytes matured with or without melatonin. Theriogenology 2021, 160, 40–49. [Google Scholar] [CrossRef]
- Edgar, D.H.; Gook, D.A. A critical appraisal of cryopreservation (slow cooling versus vitrification) of human oocytes and embryos. Hum. Reprod. Update 2012, 18, 536–554. [Google Scholar] [CrossRef] [Green Version]
- Kuwayama, M.; Vajta, G.; Kato, O.; Leibo, S.P. Highly efficient vitrification method for cryopreservation of human oocytes. Reprod. Biomed. Online 2005, 11, 300–308. [Google Scholar] [CrossRef]
- Papis, K.; Shimizu, M.; Saha, S.; Izaike, Y.; Modliński, J.A. Effects of Vitrification of Partially Denuded Bovine Immature Oocytes. Anim. Sci. Pap. Rep. 2013, 31, 14. [Google Scholar]
- Somfai, T.; Nakai, M.; Tanihara, F.; Noguchi, J.; Kaneko, H.; Kashiwazaki, N.; Egerszegi, I.; Nagai, T.; Kikuchi, K. Comparison of Ethylene Glycol and Propylene Glycol for the Vitrification of Immature Porcine Oocytes. J. Reprod. Dev. 2013, 59, 378–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Jia, B.; Quan, G.; Xiang, D.; Zhang, B.; Shao, Q.; Hong, Q. Vitrification of porcine immature oocytes: Association of equilibration manners with warming procedures, and permeating cryoprotectants effects under two temperatures. Cryobiology 2017, 75, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Van Blerkom, J.; Davis, P.W. Cytogenetic, cellular, and developmental consequences of cryopreservation of immature and mature mouse and human oocytes. Microsc. Res. Tech. 1994, 27, 165–193. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, B.; Attanasio, L.; De Rosa, A.; Monaco, E.; Di Palo, R.; Campanile, G. Cryopreservation of in vitro matured buffalo (Bubalus bubalis) oocytes by minimum volumes vitrification methods. Anim. Reprod. Sci. 2007, 98, 335–342. [Google Scholar] [CrossRef]
- Sripunya, N.; Somfai, T.; Inaba, Y.; Nagai, T.; Imai, K.; Parnpai, R. A Comparison of Cryotop and Solid Surface Vitrification Methods for the Cryopreservation of In Vitro Matured Bovine Oocytes. J. Reprod. Dev. 2010, 56, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Du, Y.; Lin, L.; Li, J.; Kragh, P.M.; Kuwayama, M.; Bolund, L.; Yang, H.; Vajta, G. Comparison of efficiency of open pulled straw (OPS) and Cryotop vitrification for cryopreservation of in vitro matured pig oocytes. Cryoletters 2008, 29, 315–320. [Google Scholar]
- Song, W.-Y.; Peng, Z.-F.; Chen, X.-M.; Jin, H.-X.; Yao, G.-D.; Shi, S.-L.; Yang, H.-Y.; Zhang, X.-Y.; Sun, Y.-P. Effects of Vitrification on Outcomes of In VivoMature, In Vitro-Mature and Immature Human Oocytes. Cell. Physiol. Biochem. 2016, 38, 2053–2062. [Google Scholar] [CrossRef]
- Sprícigo, J.; Morais, K.; Ferreira, A.; Machado, G.; Gomes, A.; Rumpf, R.; Franco, M.; Dode, M. Vitrification of bovine oocytes at different meiotic stages using the Cryotop method: Assessment of morphological, molecular and functional patterns. Cryobiology 2014, 69, 256–265. [Google Scholar] [CrossRef]
- Chaves, D.F.; Corbin, E.; Almiñana, C.; Locatelli, Y.; Souza-Fabjan, J.M.; Bhat, M.H.; Freitas, V.J.; Mermillod, P.; Sou-za-Fabjan, J.M.G.; Freitas, V.J.F. Vitrification of Immature and in Vitro Matured Bovine Cumulus-Oocyte Complexes: Effects on Oocyte Structure and Embryo Development. Livest. Sci. 2017, 199, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Mogas, T. Update on the vitrification of bovine oocytes and invitro-produced embryos. Reprod. Fertil. Dev. 2019, 31, 105–117. [Google Scholar] [CrossRef]
- Zhou, X.L.; al Naib, A.; Sun, D.W.; Lonergan, P. Bovine Oocyte Vitrification Using the Cryotop Method: Effect of Cumulus Cells and Vitrification Protocol on Survival and Subsequent Development. Cryobiology 2010, 61, 66–72. [Google Scholar] [CrossRef]
- Diez, C.; Muñoz, M.; Caamaño, J.; Gomez, E. Cryopreservation of the Bovine Oocyte: Current Status and Perspectives. Reprod. Domest. Anim. 2012, 47, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Ishii, T.; Tomita, K.; Sakakibara, H.; Ohkura, S. Embryogenesis of vitrified mature bovine oocytes is improved in the presence of multi-layered cumulus cells. J. Reprod. Dev. 2018, 64, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Dujíčková, L.; Makarevich, A.V.; Olexiková, L.; Kubovičová, E.; Strejček, F. Methodological approaches for vitrification of bovine oocytes. Zygote 2020, 29, 1–11. [Google Scholar] [CrossRef]
- Rojas, C.; Palomo, M.J.; Albarracín, J.L.; Mogas, T. Vitrification of immature and in vitro matured pig oocytes: Study of distribution of chromosomes, microtubules, and actin microfilaments. Cryobiology 2004, 49, 211–220. [Google Scholar] [CrossRef]
- Purohit, G.N.; Meena, H.; Solanki, K. Effects of Vitrification on Immature and in vitro Matured, Denuded and Cumulus Compact Goat Oocytes and Their Subsequent Fertilization. J. Reprod. Infertil. 2012, 13, 53–59. [Google Scholar]
- Maclellan, L.J.; Stokes, J.E.; Preis, K.A.; Mccue, P.M.; Carnevale, E.M. Vitrification, Warming, ICSI and Transfer of Equine Oocytes Matured in Vivo. Anim. Reprod. Sci. 2010, 121, 260–261. [Google Scholar] [CrossRef]
- Roser, J.F.; Meyers-Brown, G. Superovulation in the Mare: A Work in Progress. J. Equine Veter Sci. 2012, 32, 376–386. [Google Scholar] [CrossRef]
- Tharasanit, T.; Colleoni, S.; Galli, C.; Colenbrander, B.; Stout, T.A.E. Protective effects of the cumulus-corona radiata complex during vitrification of horse oocytes. Reproduction 2009, 137, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Agnieszka, N.; Joanna, K.; Wojciech, W.; Adam, O. In vitro maturation of equine oocytes followed by two vitrification protocols and subjected to either intracytoplasmic sperm injection (ICSI) or parthenogenic activation. Theriogenology 2021, 162, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Ducheyne, K.; Rizzo, M.; Beitsma, M.; Deelen, C.; Daels, P.; Stout, T.; De Ruijter-Villani, M. Vitrifying Equine Oocytes at the Germinal Vesicle Stage Disturbs Spindle Morphology and Chromosome Alignment. J. Equine Veter Sci. 2018, 66, 178. [Google Scholar] [CrossRef]
- Dalcin, L.; Silva, R.C.; Paulini, F.; Silva, B.D.M.; Neves, J.P.; Lucci, C.M. Cytoskeleton structure, pattern of mitochondrial activity and ultrastructure of frozen or vitrified sheep embryos. Cryobiology 2013, 67, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.-W.; Shi, W.-Q.; Zhang, Q.-J.; Zhao, X.-M.; Yan, C.L.; Hou, Y.-P.; Zhou, G.-B.; Fan, Z.-Q.; Suo, L.; Wusiman, A.; et al. Positive effects of Taxol pretreatment on morphology, distribution and ultrastructure of mitochondria and lipid droplets in vitrification of in vitro matured porcine oocytes. Anim. Reprod. Sci. 2009, 115, 158–168. [Google Scholar] [CrossRef]
- Gutnisky, C.; Morado, S.; Gadze, T.; Donato, A.; Alvarez, G.; Dalvit, G.; Cetica, P. Morphological, biochemical and functional studies to evaluate bovine oocyte vitrification. Theriogenology 2020, 143, 18–26. [Google Scholar] [CrossRef]
- Berthelot-Ricou, A.; Perrin, J.; di Giorgio, C.; de Meo, M.; Botta, A.; Courbiere, B. Assessment of 1,2-propanediol (PrOH) genotoxicity on mouse oocytes by comet assay. Fertil. Steril. 2011, 96, 1002–1007. [Google Scholar] [CrossRef]
- Ishida, G.M.; Saito, H.; Ohta, N.; Takahashi, T.; Ito, M.M.; Saito, T.; Nakahara, K.; Hiroi, M. The optimal equilibration time for mouse embryos frozen by vitrification with trehalose. Hum. Reprod. 1997, 12, 1259–1262. [Google Scholar] [CrossRef] [Green Version]
- Sterzik, K.; Rosenbusch, B.; Grab, D.; Wahl, A.; Beier, H.M.; Lauritzen, C. Numerical chromosome anomalies after fertilization of freeze-thawed mouse oocytes. Arch. Gynecol. Obstet. 1992, 251, 133–138. [Google Scholar] [CrossRef]
- Tamura, A.N.; Huang, T.T.; Marikawa, Y. Impact of Vitrification on the Meiotic Spindle and Components of the Microtubule-Organizing Center in Mouse Mature Oocytes1. Biol. Reprod. 2013, 89, 112. [Google Scholar] [CrossRef]
- Hinrichs, K.; Love, L.; Varner, D.; Walckenaer, B.; Choi, Y. Chromatin Configuration Within the Germinal Vesicle of Horse Oocytes: Changes Post Mortem and Relationship to Meiotic and Developmental Competence1. Biol. Reprod. 2005, 72, 1142–1150. [Google Scholar] [CrossRef]
- De Leon, P.; Campos, V.; Corcini, C.; Santos, E.; Rambo, G.; Lucia, T.; Deschamps, J.; Collares, T. Cryopreservation of immature equine oocytes, comparing a solid surface vitrification process with open pulled straws and the use of a synthetic ice blocker. Theriogenology 2012, 77, 21–27. [Google Scholar] [CrossRef]
- Canesin, H.S.; Brom-De-Luna, J.G.; Choi, Y.-H.; Ortiz, I.; Diaw, M.; Hinrichs, K. Blastocyst development after intracytoplasmic sperm injection of equine oocytes vitrified at the germinal-vesicle stage. Cryobiology 2017, 75, 52–59. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Li, X.-X.; Xu, Y.-K.; Wu, H.; Zheng, J.-J.; Yu, X.-L. Developmental competence and gene expression of immature oocytes following liquid helium vitrification in bovine. Cryobiology 2014, 69, 428–433. [Google Scholar] [CrossRef]
- Ebrahimi, B.; Valojerdi, M.R.; Eftekhari-Yazdi, P.; Baharvand, H.; Farrokhi, A. IVM and gene expression of sheep cumulus–oocyte complexes following different methods of vitrification. Reprod. Biomed. Online 2010, 20, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Azari, M.; Kafi, M.; Ebrahimi, B.; Fatehi, R.; Jamalzadeh, M. Oocyte maturation, embryo development and gene expression following two different methods of bovine cumulus-oocyte complexes vitrification. Veter Res. Commun. 2017, 41, 49–56. [Google Scholar] [CrossRef]
- Turathum, B.; Saikhun, K.; Sangsuwan, P.; Kitiyanant, Y. Effects of Vitrification on Nuclear Maturation, Ultrastructural Changes and Gene Expression of Canine Oocytes. Reprod. Biol. Endocrinol. 2010, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Stigliani, S.; Moretti, S.; Anserini, P.; Casciano, I.; Venturini, P.L.; Scaruffi, P. Storage time does not modify the gene expression profile of cryopreserved human metaphase II oocytes. Hum. Reprod. 2015, 30, 2519–2526. [Google Scholar] [CrossRef] [Green Version]
- Monzo, C.; Haouzi, D.; Roman, K.; Assou, S.; Dechaud, H.; Hamamah, S. Slow freezing and vitrification differentially modify the gene expression profile of human metaphase II oocytes. Hum. Reprod. 2012, 27, 2160–2168. [Google Scholar] [CrossRef]
- Barberet, J.; Ducreux, B.; Bruno, C.; Guilleman, M.; Simonot, R.; Lieury, N.; Guilloteau, A.; Bourc’His, D.; Fauque, P. Comparison of oocyte vitrification using a semi-automated or a manual closed system in human siblings: Survival and transcriptomic analyses. J. Ovarian Res. 2022, 15, 1–14. [Google Scholar] [CrossRef]
- Ma, Y.; Long, C.; Liu, G.; Bai, H.; Ma, L.; Bai, T.; Zuo, Y.; Li, S. WGBS combined with RNA-seq analysis revealed that Dnmt1 affects the methylation modification and gene expression changes during mouse oocyte vitrification. Theriogenology 2021, 177, 11–21. [Google Scholar] [CrossRef]
- Gao, L.; Jia, G.; Li, A.; Ma, H.; Huang, Z.; Zhu, S.; Hou, Y.; Fu, X. RNA-Seq transcriptome profiling of mouse oocytes after in vitro maturation and/or vitrification. Sci. Rep. 2017, 7, 13245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Ma, Y.; Wei, S.; Pan, B.; Qi, Y.; Hou, Y.; Meng, Q.; Zhou, G.; Han, H. Dynamic Changes in the Global Transcriptome of Bovine Germinal Vesicle Oocytes after Vitrification Followed by in Vitro Maturation. Reprod. Fertil. Dev. 2018, 30, 1298–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Li, C.-Y.; Zhu, H.-B.; Hao, H.-S.; Wang, H.-Y.; Yan, C.-L.; Zhao, S.-J.; Du, W.-H.; Wang, D.; Liu, Y.; et al. Effect of vitrification on the mRNA transcriptome of bovine oocytes. Reprod. Domest. Anim. 2017, 52, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, Z.-Y.; Cai, M.-D.; Li, X.-X.; Li, Y.-H.; Lei, Y.; Yu, X.-L. Effect of vitrification temperature and cryoprotectant concentrations on the mRNA transcriptome of bovine mature oocytes after vitrifying at immature stage. Theriogenology 2020, 148, 225–235. [Google Scholar] [CrossRef]
- Jia, B.-Y.; Xiang, D.-C.; Quan, G.-B.; Zhang, B.; Shao, Q.-Y.; Hong, Q.-H.; Wu, G.-Q. Transcriptome analysis of porcine immature oocytes and surrounding cumulus cells after vitrification and in vitro maturation. Theriogenology 2019, 134, 90–97. [Google Scholar] [CrossRef]
- Matzuk, M.M.; Burns, K.H.; Viveiros, M.M.; Eppig, J.J. Intercellular Communication in the Mammalian Ovary: Oocytes Carry the Conversation. Science 2002, 296, 2178–2180. [Google Scholar] [CrossRef]
- Labrecque, R.; Sirard, M.-A. The study of mammalian oocyte competence by transcriptome analysis: Progress and challenges. Mol. Hum. Reprod. 2014, 20, 103–116. [Google Scholar] [CrossRef]
- Telford, N.A.; Watson, A.J.; Schultz, G.A. Transition from maternal to embryonic control in early mammalian development: A comparison of several species. Mol. Reprod. Dev. 1990, 26, 90–100. [Google Scholar] [CrossRef]
- Eppig, J.J. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod. Fertil. Dev. 1996, 8, 485–489. [Google Scholar] [CrossRef]
- Sirard, M. Resumption of meiosis: Mechanism involved in meiotic progression and its relation with developmental competence. Theriogenology 2001, 55, 1241–1254. [Google Scholar] [CrossRef]
- Bouniol-Baly, C.; Hamraoui, L.; Guibert, J.; Beaujean, N.; Szöllösi, M.S.; Debey, P. Differential Transcriptional Activity Associated with Chromatin Configuration in Fully Grown Mouse Germinal Vesicle Oocytes. Biol. Reprod. 1999, 60, 580–587. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, R.; Viveiros, M.M.; Burns, K.H.; Adashi, E.Y.; Matzuk, M.M.; Eppig, J.J. Major chromatin remodeling in the germinal vesicle (GV) of mammalian oocytes is dispensable for global transcriptional silencing but required for centromeric heterochromatin function. Dev. Biol. 2004, 275, 447–458. [Google Scholar] [CrossRef]
- Su, Y.-Q.; Sugiura, K.; Woo, Y.; Wigglesworth, K.; Kamdar, S.; Affourtit, J.; Eppig, J.J. Selective degradation of transcripts during meiotic maturation of mouse oocytes. Dev. Biol. 2007, 302, 104–117. [Google Scholar] [CrossRef] [Green Version]
- Sha, Q.; Yu, J.; Guo, J.; Dai, X.; Jiang, J.; Zhang, Y.; Yu, C.; Ji, S.; Jiang, Y.; Zhang, S.; et al. CNOT 6L couples the selective degradation of maternal transcripts to meiotic cell cycle progression in mouse oocyte. EMBO J. 2018, 37, e99333. [Google Scholar] [CrossRef]
- Takeuchi, H.; Yamamoto, M.; Fukui, M.; Inoue, A.; Maezawa, T.; Nishioka, M.; Kondo, E.; Ikeda, T.; Matsumoto, K.; Miyamoto, K. Single-Cell Profiling of Transcriptomic Changes during in Vitro Maturation of Human Oocytes. Reprod. Med. Biol. 2022, 21, e12464. [Google Scholar] [CrossRef]
- Brevini, T.; Lonergan, P.; Cillo, F.; Francisci, C.; Favetta, L.; Fair, T.; Gandolfi, F. Evolution of mRNA polyadenylation between oocyte maturation and first embryonic cleavage in cattle and its relation with developmental competence. Mol. Reprod. Dev. 2002, 63, 510–517. [Google Scholar] [CrossRef]
- Reyes, J.M.; Chitwood, J.L.; Ross, P.J. RNA-Seq profiling of single bovine oocyte transcript abundance and its modulation by cytoplasmic polyadenylation. Mol. Reprod. Dev. 2015, 82, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Fuqua, B.K.; Hasin, Y.; Yukhtman, C.; Vulpe, C.D.; Lusis, A.J.; Pellegrini, M. A Comparison between Whole Transcript and 3’ RNA Sequencing Methods Using Kapa and Lexogen Library Preparation Methods 06 Biological Sciences 0604 Genetics. BMC Genom. 2019, 20, 9. [Google Scholar] [CrossRef]
- Somfai, T.; Men, N.T.; Noguchi, J.; Kaneko, H.; Kashiwazaki, N.; Kikuchi, K. Optimization of cryoprotectant treatment for the vitrification of immature cumulus-enclosed porcine oocytes: Comparison of sugars, combinations of permeating cryoprotectants and equilibration regimens. J. Reprod. Dev. 2015, 61, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Escribano, N.; Smits, K.; Piepers, S.; Abbeel, E.V.D.; Woelders, H.; Van Soom, A. Role of cumulus cells during vitrification and fertilization of mature bovine oocytes: Effects on survival, fertilization, and blastocyst development. Theriogenology 2016, 86, 635–641. [Google Scholar] [CrossRef]
- Shirazi, A.; Naderi, M.M.; Hassanpour, H.; Heidari, M.; Borjian, S.; Sarvari, A.; Akhondi, M.M. The effect of ovine oocyte vitrification on expression of subset of genes involved in epigenetic modifications during oocyte maturation and early embryo development. Theriogenology 2016, 86, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, A.; Scoggin, C.; Bradecamp, E.; Ali, H.; Troedsson, M.; Meyers, S.; Dini, P.; de la Fuente, A.; Scoggin, C.; Bradecamp, E.; et al. 146 Transcriptome Characterisation of Equine Oocyte Maturation. Reprod. Fertil. Dev. 2021, 34, 311. [Google Scholar] [CrossRef] [PubMed]
- Bachvarova, R.; De Leon, V.; Johnson, A.; Kaplan, G.; Paynton, B.V. Changes in total RNA, polyadenylated RNA, and actin mRNA during meiotic maturation of mouse oocytes. Dev. Biol. 1985, 108, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Chamayou, S.; Bonaventura, G.; Alecci, C.; Tibullo, D.; Di Raimondo, F.; Guglielmino, A.; Barcellona, M. Consequences of metaphase II oocyte cryopreservation on mRNA content. Cryobiology 2011, 62, 130–134. [Google Scholar] [CrossRef]
- Huo, Y.; Yuan, P.; Qin, Q.; Yan, Z.; Yan, L.; Liu, P.; Li, R.; Yan, J.; Qiao, J. Effects of vitrification and cryostorage duration on single-cell RNA-Seq profiling of vitrified-thawed human metaphase II oocytes. Front. Med. 2021, 15, 144–154. [Google Scholar] [CrossRef]
- Chang, C.-C.; Shapiro, D.B.; Bernal, D.P.; Wright, G.; I Kort, H.; Nagy, Z.P. Human oocyte vitrification: In-vivo and in-vitro maturation outcomes. Reprod. Biomed. Online 2008, 17, 684–688. [Google Scholar] [CrossRef]
- Inoue, J.; Gohda, J.; Akiyama, T. Characteristics and Biological Functions of TRAF6. In TNF Receptor Associated Factors (TRAFs); Springer: Berlin/Heidelberg, Germany, 2007; pp. 72–79. [Google Scholar]
- He, L.; Wu, X.; Siegel, R.; Lipsky, P.E. TRAF6 Regulates Cell Fate Decisions by Inducing Caspase 8-dependent Apoptosis and the Activation of NF-κB. J. Biol. Chem. 2006, 281, 11235–11249. [Google Scholar] [CrossRef] [Green Version]
- Perkins, N.D. Integrating cell-signalling pathways with NF-κB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Krishnaswamy, J.K.; Singh, A.; Gowthaman, U.; Wu, R.; Gorrepati, P.; Nascimento, M.S.; Gallman, A.; Liu, D.; Rhebergen, A.M.; Calabro, S.; et al. Coincidental loss of DOCK8 function in NLRP10-deficient and C3H/HeJ mice results in defective dendritic cell migration. Proc. Natl. Acad. Sci. USA 2015, 112, 3056–3061. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Torii, T.; Kawahara, K.; Tanoue, A.; Yamauchi, J. Dock8 interacts with Nck1 in mediating Schwann cell precursor migration. Biochem. Biophys. Rep. 2016, 6, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Soma-Nagae, T.; Nada, S.; Kitagawa, M.; Takahashi, Y.; Mori, S.; Oneyama, C.; Okada, M. The Lysosomal Signaling Anchor P18/LAMTOR1 Controls Epidermal Development by Regulating Lysosome-Mediated Catabolic Processes. J. Cell. Sci. 2013, 126, 3575–3584. [Google Scholar] [CrossRef] [Green Version]
- Gumus, E.; Sari, I.; Yilmaz, M.; Cetin, A. Investigation of LAMTOR1 gene and protein expressions in germinal vesicle and metaphase II oocytes and embryos from 1-cell to blastocyst stage in a mouse model. Gene Expr. Patterns 2018, 28, 72–76. [Google Scholar] [CrossRef]
- Nakaseko, Y.; Yanagida, M. Cytoskeleton in the cell cycle. Nature 2001, 412, 291–292. [Google Scholar] [CrossRef]
- Humeau, J.; Bravo-San Pedro, J.M.; Vitale, I.; Nuñez, L.; Villalobos, C.; Kroemer, G.; Senovilla, L. Calcium signaling and cell cycle: Progression or death. Cell Calcium 2018, 70, 3–15. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Girka, E.; Gatenby, L.; Gutierrez, E.J.; Bondioli, K.R. The effects of microtubule stabilizing and recovery agents on vitrified bovine oocytes. Theriogenology 2022, 182, 9–16. [Google Scholar] [CrossRef]
- Pitchayapipatkul, J.; Somfai, T.; Matoba, S.; Parnpai, R.; Nagai, T.; Geshi, M.; Vongpralub, T. Microtubule stabilisers docetaxel and paclitaxel reduce spindle damage and maintain the developmental competence of in vitro-mature bovine oocytes during vitrification. Reprod. Fertil. Dev. 2017, 29, 2028–2039. [Google Scholar] [CrossRef]
- E Park, S.; Chung, H.M.; Cha, K.Y.; Hwang, W.S.; Lee, E.S.; Lim, J.M. Cryopreservation of ICR mouse oocytes: Improved post-thawed preimplantation development after vitrification using Taxol™, a cytoskeleton stabilizer. Fertil. Steril. 2001, 75, 1177–1184. [Google Scholar] [CrossRef]
- Succu, S.; Berlinguer, F.; Leoni, G.G.; Bebbere, D.; Satta, V.; Marco-Jiménez, F.; Pasciu, V.; Naitana, S. Calcium concentration in vitrification medium affects the developmental competence of in vitro matured ovine oocytes. Theriogenology 2011, 75, 715–721. [Google Scholar] [CrossRef]
- Larman, M.G.; Sheehan, C.B.; Gardner, D.K. Calcium-free vitrification reduces cryoprotectant-induced zona pellucida hardening and increases fertilization rates in mouse oocytes. Reproduction 2006, 131, 53–61. [Google Scholar] [CrossRef]
- Paynton, B.V.; Rempel, R.; Bachvarova, R. Changes in state of adenylation and time course of degradation of maternal mRNAs during oocyte maturation and early embryonic development in the mouse. Dev. Biol. 1988, 129, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.; Hinrichs, K.; Schnauffer, K.; Morganti, M.; Argo, C.M. Effect of Oocyte Source and Transport Time on Rates of Equine Oocyte Maturation and Cleavage after Fertilization by ICSI, with a Note on the Validation of Equine Embryo Morphological Classification. Clin. Theriogenol. 2016, 8, 25–39. [Google Scholar]
- Krueger, F.T.G. A Wrapper Tool around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files. 2015. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 13 February 2023).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Heger, A.; Sudbery, I. UMI-tools: Modeling sequencing errors in Unique Molecular Identifiers to improve quantification accuracy. Genome Res. 2017, 27, 491–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Geistlinger, L.; Csaba, G.; Santarelli, M.; Ramos, M.; Schiffer, L.; Turaga, N.; Law, C.; Davis, S.; Carey, V.; Morgan, M.; et al. Toward a gold standard for benchmarking gene set enrichment analysis. Briefings Bioinform. 2021, 22, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Tarca, A.L.; Draghici, S.; Bhatti, G.; Romero, R. Down-weighting overlapping genes improves gene set analysis. BMC Bioinform. 2012, 13, 136. [Google Scholar] [CrossRef] [Green Version]
- Geistlinger, L.; Csaba, G.; Zimmer, R. Bioconductor’s EnrichmentBrowser: Seamless Navigation through Combined Results of Set- & Network-Based Enrichment Analysis. BMC Bioinform. 2016, 17, 45. [Google Scholar] [CrossRef]
- Gu, Z.; Hübschmann, D. Simplify enrichment: A bioconductor package for clustering and visualizing functional enrichment results. Genom. Proteom. Bioinform. 2022. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- CRAN—Package MultcompView. Available online: https://cran.r-project.org/web/packages/multcompView/index.html (accessed on 8 December 2022).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angel-Velez, D.; Meese, T.; Hedia, M.; Fernandez-Montoro, A.; De Coster, T.; Pascottini, O.B.; Van Nieuwerburgh, F.; Govaere, J.; Van Soom, A.; Pavani, K.; et al. Transcriptomics Reveal Molecular Differences in Equine Oocytes Vitrified before and after In Vitro Maturation. Int. J. Mol. Sci. 2023, 24, 6915. https://doi.org/10.3390/ijms24086915
Angel-Velez D, Meese T, Hedia M, Fernandez-Montoro A, De Coster T, Pascottini OB, Van Nieuwerburgh F, Govaere J, Van Soom A, Pavani K, et al. Transcriptomics Reveal Molecular Differences in Equine Oocytes Vitrified before and after In Vitro Maturation. International Journal of Molecular Sciences. 2023; 24(8):6915. https://doi.org/10.3390/ijms24086915
Chicago/Turabian StyleAngel-Velez, Daniel, Tim Meese, Mohamed Hedia, Andrea Fernandez-Montoro, Tine De Coster, Osvaldo Bogado Pascottini, Filip Van Nieuwerburgh, Jan Govaere, Ann Van Soom, Krishna Pavani, and et al. 2023. "Transcriptomics Reveal Molecular Differences in Equine Oocytes Vitrified before and after In Vitro Maturation" International Journal of Molecular Sciences 24, no. 8: 6915. https://doi.org/10.3390/ijms24086915