
Genome-Wide CRISPR Screens Reveal ZATT as a Synthetic Lethal Target of TOP2-Poison Etoposide That Can Act in a TDP2-Independent Pathway
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
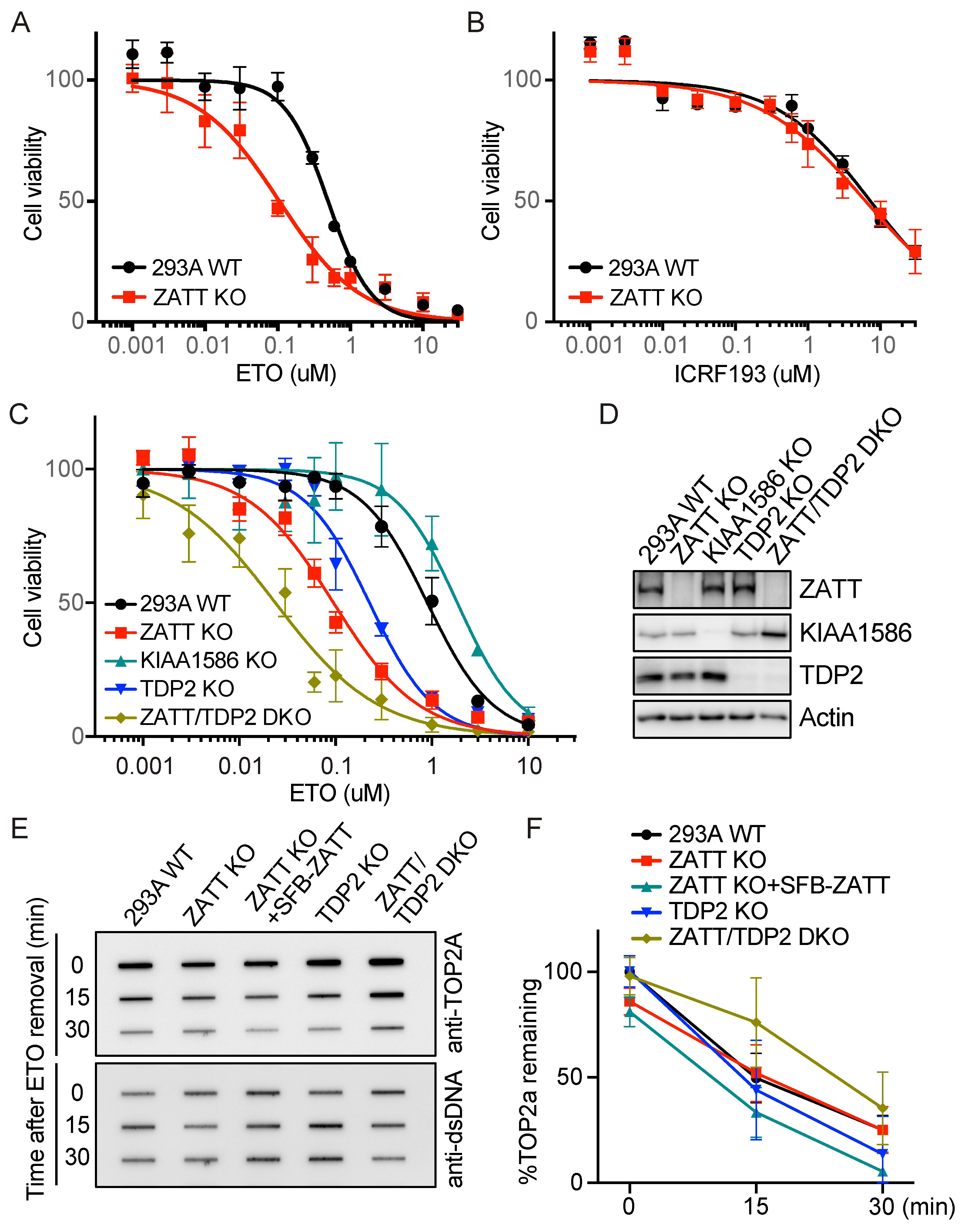
2.1. ZATT KO Cells Showed More Enhanced ETO Sensitivity Than TDP2 KO Cells
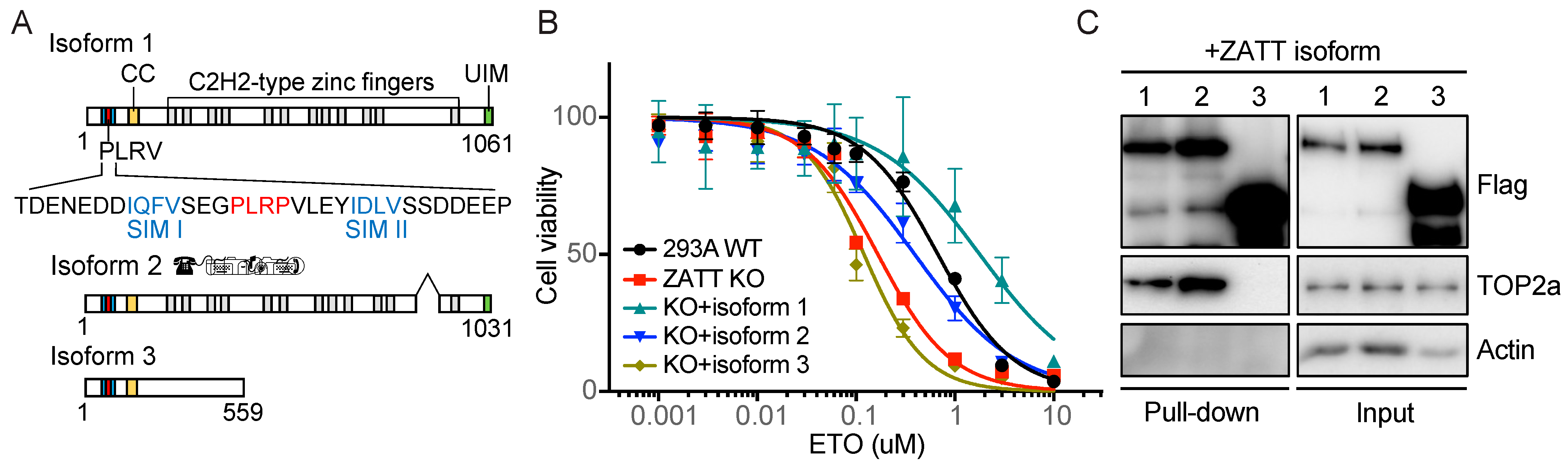
2.2. ZATT Isoform 3 Was Not Able to Rescue Etoposide Sensitivity in ZATT KO Cells
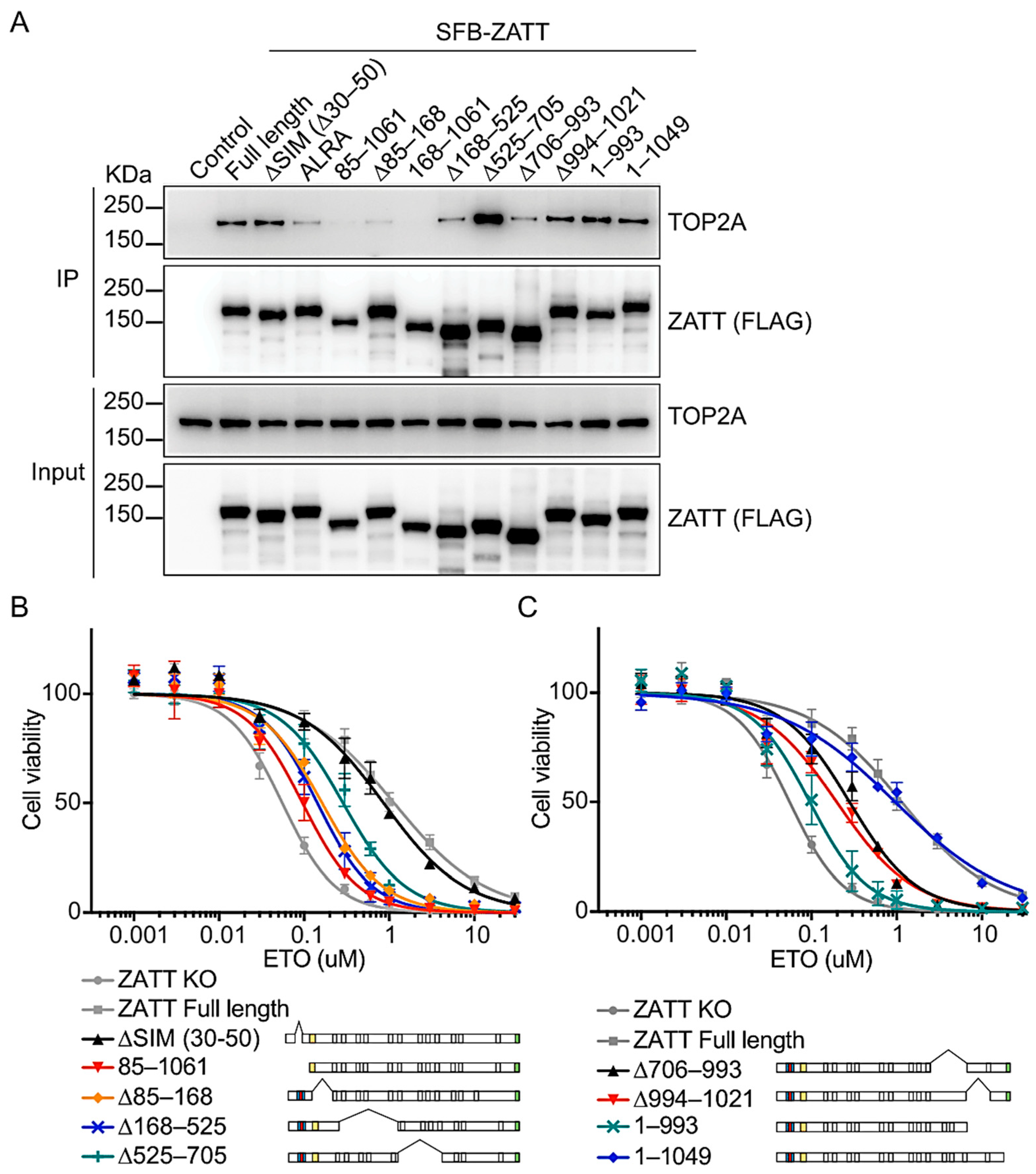
2.3. The N-Terminal 1–168 Residues of ZATT Is Essential for Its Interaction with TOP2
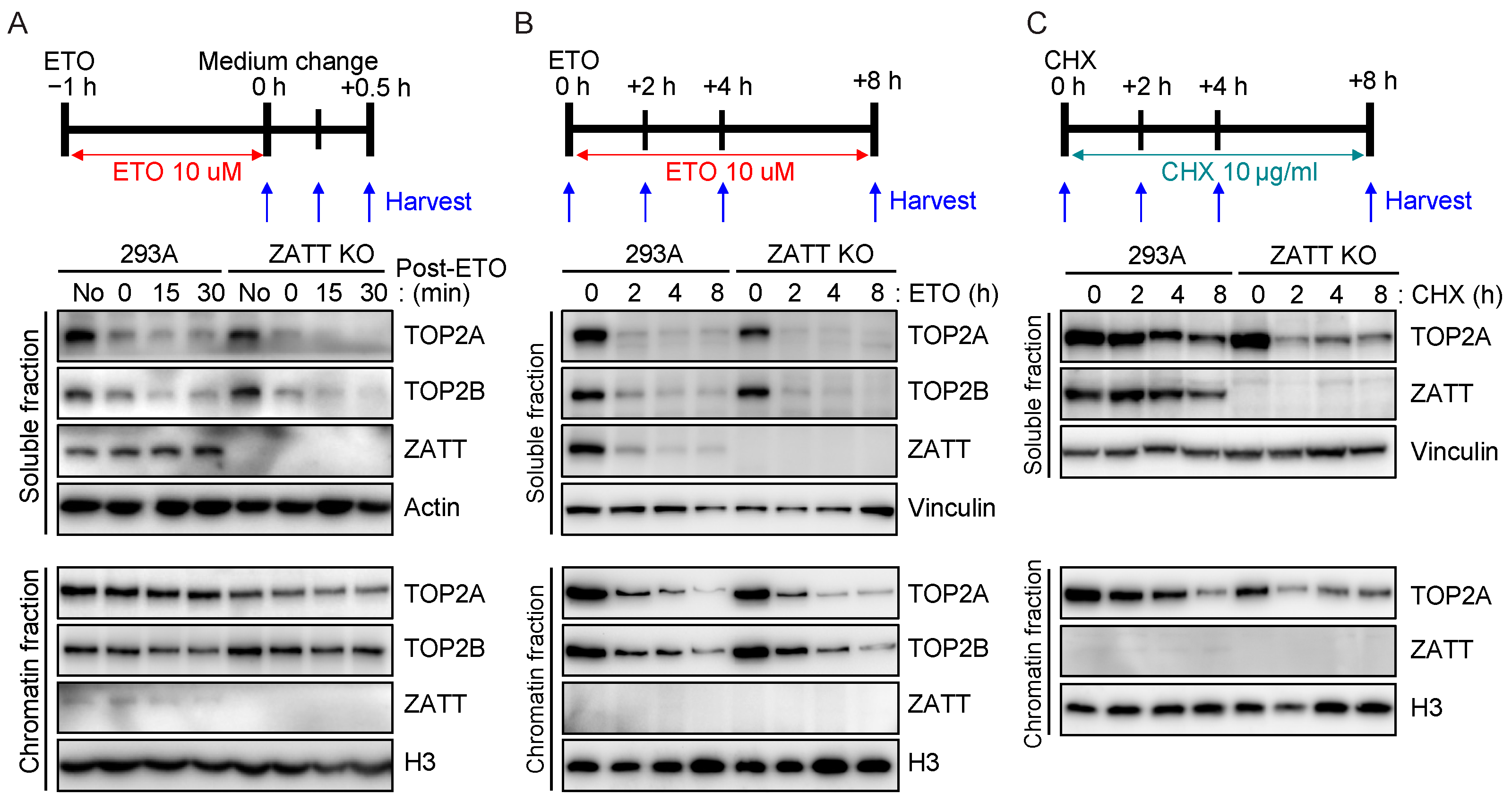
2.4. ZATT Depletion Leads to Destabilization of TOP2 Protein
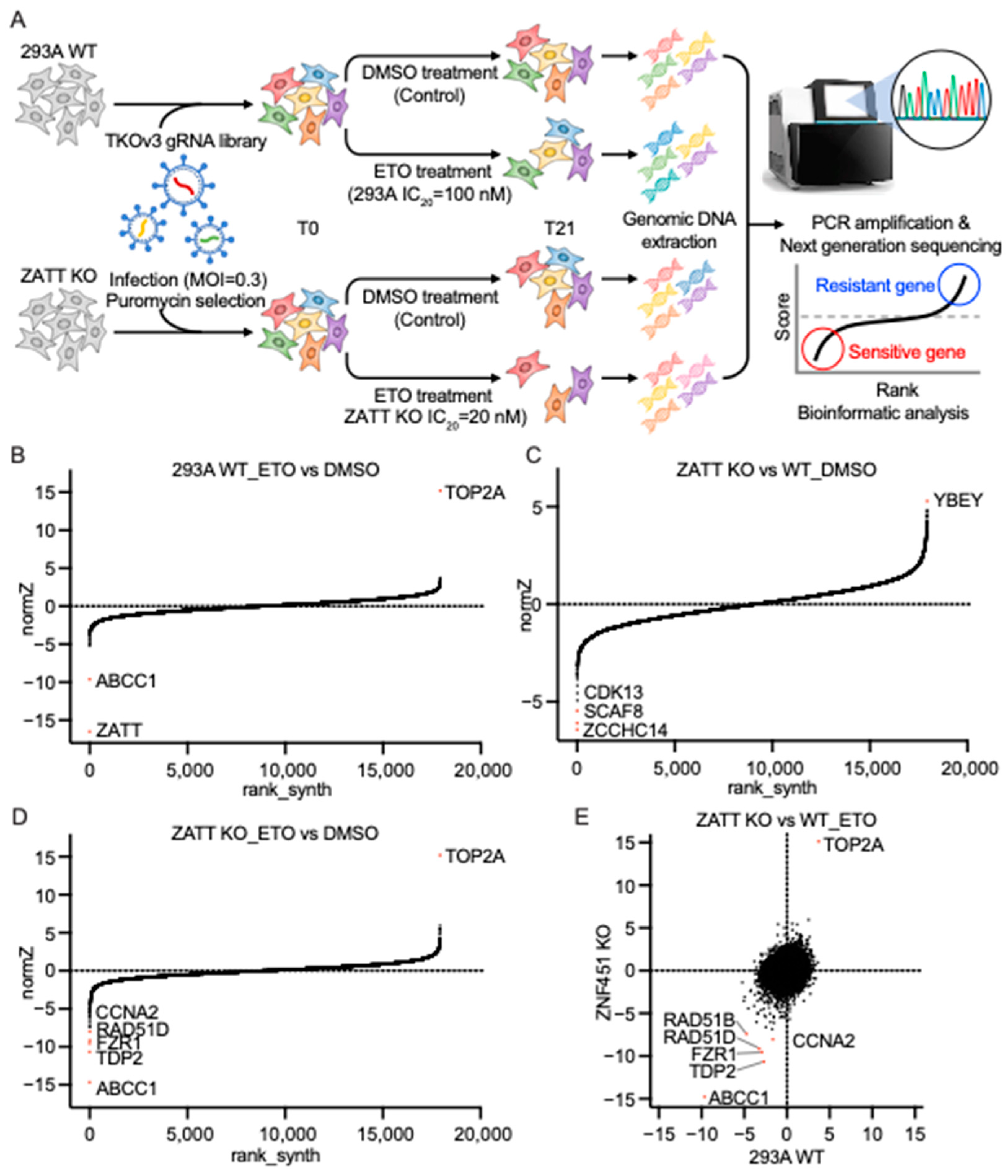
2.5. Genome-Wide CRISPR Screens Identified ZATT as a Critical Gene for ETO Sensitivity
3. Discussion
4. Materials and Methods
4.1. Cell Lines and CRISPR/Cas9-Mediated Gene Knockout
- ZATT-sgRNA #1: 5′ CACCGTGTTCTTGAATACATTGATC 3′
- KIAA1586 sgRNA #1: 5′ CACCCTGGAGATAAATCACTAGGG 3′
- TDP2-sgRNA #1: 5′ CACCGTCTCCCAGTCGTTCTCGGCC 3′
4.2. CellTiter-Glo Assay
4.3. Genome-Wide CRISPR/Cas9 Screens
4.4. Plasmids and Reconstitution Experiments
4.5. Western Blot Analysis
4.6. Rapid Approach to DNA Adduct Recovery Assay
4.7. Immunoprecipitation
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.C. Cellular roles of DNA topoisomerases: A molecular perspective. Nat. Rev. Mol. Cell Biol. 2002, 3, 430–440. [Google Scholar] [CrossRef]
- Pommier, Y.; Nussenzweig, A.; Takeda, S.; Austin, C. Human topoisomerases and their roles in genome stability and organization. Nat. Rev. Mol. Cell Biol. 2022, 23, 407–427. [Google Scholar] [CrossRef] [PubMed]
- Nitiss, J.L. DNA topoisomerase II and its growing repertoire of biological functions. Nat. Rev. Cancer 2009, 9, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.F.; Rowe, T.C.; Yang, L.; Tewey, K.M.; Chen, G.L. Cleavage of DNA by mammalian DNA topoisomerase II. J. Biol. Chem. 1983, 258, 15365–15370. [Google Scholar] [CrossRef] [PubMed]
- Canela, A.; Maman, Y.; Huang, S.N.; Wutz, G.; Tang, W.; Zagnoli-Vieira, G.; Callen, E.; Wong, N.; Day, A.; Peters, J.M.; et al. Topoisomerase II-Induced Chromosome Breakage and Translocation Is Determined by Chromosome Architecture and Transcriptional Activity. Mol. Cell 2019, 75, 252–266.e8. [Google Scholar] [CrossRef]
- Sciascia, N.; Wu, W.; Zong, D.; Sun, Y.; Wong, N.; John, S.; Wangsa, D.; Ried, T.; Bunting, S.F.; Pommier, Y.; et al. Suppressing proteasome mediated processing of topoisomerase II DNA-protein complexes preserves genome integrity. Elife 2020, 9, e53447. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Cortés-Ledesma, F.; El Khamisy, S.F.; Caldecott, K.W. TDP2/TTRAP is the major 5’-tyrosyl DNA phosphodiesterase activity in vertebrate cells and is critical for cellular resistance to topoisomerase II-induced DNA damage. J. Biol. Chem. 2011, 286, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Cortes Ledesma, F.; El Khamisy, S.F.; Zuma, M.C.; Osborn, K.; Caldecott, K.W. A human 5’-tyrosyl DNA phosphodiesterase that repairs topoisomerase-mediated DNA damage. Nature 2009, 461, 674–678. [Google Scholar] [CrossRef]
- Gao, R.; Schellenberg, M.J.; Huang, S.Y.; Abdelmalak, M.; Marchand, C.; Nitiss, K.C.; Nitiss, J.L.; Williams, R.S.; Pommier, Y. Proteolytic degradation of topoisomerase II (Top2) enables the processing of Top2·DNA and Top2·RNA covalent complexes by tyrosyl-DNA-phosphodiesterase 2 (TDP2). J. Biol. Chem. 2014, 289, 17960–17969. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.C.; Swan, R.L.; Sondka, Z.; Padget, K.; Cowell, I.G.; Austin, C.A. Effect of TDP2 on the Level of TOP2-DNA Complexes and SUMOylated TOP2-DNA Complexes. Int. J. Mol. Sci. 2018, 19, 2056. [Google Scholar] [CrossRef] [Green Version]
- Koidl, S.; Eisenhardt, N.; Fatouros, C.; Droescher, M.; Chaugule, V.K.; Pichler, A. The SUMO2/3 specific E3 ligase ZNF451-1 regulates PML stability. Int. J. Biochem. Cell Biol. 2016, 79, 478–487. [Google Scholar] [CrossRef]
- Schellenberg, M.J.; Lieberman, J.A.; Herrero-Ruiz, A.; Butler, L.R.; Williams, J.G.; Munoz-Cabello, A.M.; Mueller, G.A.; London, R.E.; Cortes-Ledesma, F.; Williams, R.S. ZATT (ZNF451)-mediated resolution of topoisomerase 2 DNA-protein cross-links. Science 2017, 357, 1412–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagnoli-Vieira, G.; Caldecott, K.W. TDP2, TOP2, and SUMO: What is ZATT about? Cell Res. 2017, 27, 1405–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Bu, M.; Chen, X.; Ding, L.; Yang, Y.; Han, J.; Feng, X.H.; Xu, P.; Liu, T.; Ying, S.; et al. The ZATT-TOP2A-PICH Axis Drives Extensive Replication Fork Reversal to Promote Genome Stability. Mol. Cell 2021, 81, 198–211.E6. [Google Scholar] [CrossRef] [PubMed]
- Riccio, A.A.; Schellenberg, M.J.; Williams, R.S. Molecular mechanisms of topoisomerase 2 DNA-protein crosslink resolution. Cell Mol. Life Sci. 2020, 77, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Eisenhardt, N.; Chaugule, V.K.; Koidl, S.; Droescher, M.; Dogan, E.; Rettich, J.; Sutinen, P.; Imanishi, S.Y.; Hofmann, K.; Palvimo, J.J.; et al. A new vertebrate SUMO enzyme family reveals insights into SUMO-chain assembly. Nat. Struct. Mol. Biol. 2015, 22, 959–967. [Google Scholar] [CrossRef]
- Lee, K.C.; Bramley, R.L.; Cowell, I.G.; Jackson, G.H.; Austin, C.A. Proteasomal inhibition potentiates drugs targeting DNA topoisomerase II. Biochem. Pharmacol. 2016, 103, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Cappadocia, L.; Pichler, A.; Lima, C.D. Structural basis for catalytic activation by the human ZNF451 SUMO E3 ligase. Nat. Struct. Mol. Biol. 2015, 22, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.L.; Malotky, E.; O’Bryan, J.P. Analysis of the role of ubiquitin-interacting motifs in ubiquitin binding and ubiquitylation. J. Biol. Chem. 2004, 279, 33528–33537. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; North, B.J.; Shu, J. Regulation of topoisomerase II stability and activity by ubiquitination and SUMOylation: Clinical implications for cancer chemotherapy. Mol. Biol. Rep. 2021, 48, 6589–6601. [Google Scholar] [CrossRef]
- Cole, S.P. Multidrug resistance protein 1 (MRP1, ABCC1), a “multitasking” ATP-binding cassette (ABC) transporter. J. Biol. Chem. 2014, 289, 30880–30888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhang, P. The function of APC/CCdh1 in cell cycle and beyond. Cell Div. 2009, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagano, M.; Pepperkok, R.; Verde, F.; Ansorge, W.; Draetta, G. Cyclin A is required at two points in the human cell cycle. EMBO J. 1992, 11, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.W.; Mok, J.W.; Telerman, S.B.; Amson, R.; Telerman, A.; Choi, K.W. Topoisomerase II is regulated by translationally controlled tumor protein for cell survival during organ growth in Drosophila. Cell Death Dis. 2021, 12, 811. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Song, C.; Du, X.; Zhang, C.; Liu, Y.; Liang, L.; He, J.; Lamb, K.; Shen, W.H.; Yin, Y. PTEN stabilizes TOP2A and regulates the DNA decatenation. Sci. Rep. 2015, 5, 17873. [Google Scholar] [CrossRef] [Green Version]
- Nabet, B.; Ferguson, F.M.; Seong, B.K.A.; Kuljanin, M.; Leggett, A.L.; Mohardt, M.L.; Robichaud, A.; Conway, A.S.; Buckley, D.L.; Mancias, J.D.; et al. Rapid and direct control of target protein levels with VHL-recruiting dTAG molecules. Nat. Commun. 2020, 11, 4687. [Google Scholar] [CrossRef]
- Eguren, M.; Alvarez-Fernandez, M.; Garcia, F.; Lopez-Contreras, A.J.; Fujimitsu, K.; Yaguchi, H.; Luque-Garcia, J.L.; Fernandez-Capetillo, O.; Munoz, J.; Yamano, H.; et al. A Synthetic Lethal Interaction between APC/C and Topoisomerase Poisons Uncovered by Proteomic Screens. Cell Rep. 2014, 6, 670–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.M.; Li, J.Y.; Hu, R.K.; Xu, B.J.; Huang, G.L.; Huang, W.L.; Chen, B.; He, J.; Cao, Y. Cyclin A2/cyclin-dependent kinase 1-dependent phosphorylation of Top2a is required for S phase entry during retinal development in zebrafish. J. Genet. Genom. 2021, 48, 63–74. [Google Scholar] [CrossRef]
- Chen, T.; Sun, Y.; Ji, P.; Kopetz, S.; Zhang, W. Topoisomerase IIalpha in chromosome instability and personalized cancer therapy. Oncogene 2015, 34, 4019–4031. [Google Scholar] [CrossRef] [Green Version]
- Antoniou-Kourounioti, M.; Mimmack, M.L.; Porter, A.C.G.; Farr, C.J. The Impact of the C-Terminal Region on the Interaction of Topoisomerase II Alpha with Mitotic Chromatin. Int. J. Mol. Sci. 2019, 20, 1238. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Feng, X.; Su, D.; Wang, G.; Wang, C.; Tang, M.; Paulucci-Holthauzen, A.; Hart, T.; Chen, J. Genome-wide CRISPR screen uncovers a synergistic effect of combining Haspin and Aurora kinase B inhibition. Oncogene 2020, 39, 4312–4322. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Tang, M.; Dede, M.; Su, D.; Pei, G.; Jiang, D.; Wang, C.; Chen, Z.; Li, M.; Nie, L.; et al. Genome-wide CRISPR screens using isogenic cells reveal vulnerabilities conferred by loss of tumor suppressors. Sci. Adv. 2022, 8, eabm6638. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, G.; Feng, X.; Shepherd, P.; Zhang, J.; Tang, M.; Chen, Z.; Srivastava, M.; McLaughlin, M.E.; Navone, N.M.; et al. Genome-wide CRISPR screens reveal synthetic lethality of RNASEH2 deficiency and ATR inhibition. Oncogene 2019, 38, 2451–2463. [Google Scholar] [CrossRef]
- Kiianitsa, K.; Maizels, N. A rapid and sensitive assay for DNA-protein covalent complexes in living cells. Nucleic. Acids. Res. 2013, 41, e104. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-M.; Zhang, H.; Nie, L.; Wang, C.; Huang, M.; Feng, X.; Tang, M.; Chen, Z.; Xiong, Y.; Lee, N.; et al. Genome-Wide CRISPR Screens Reveal ZATT as a Synthetic Lethal Target of TOP2-Poison Etoposide That Can Act in a TDP2-Independent Pathway. Int. J. Mol. Sci. 2023, 24, 6545. https://doi.org/10.3390/ijms24076545
Park J-M, Zhang H, Nie L, Wang C, Huang M, Feng X, Tang M, Chen Z, Xiong Y, Lee N, et al. Genome-Wide CRISPR Screens Reveal ZATT as a Synthetic Lethal Target of TOP2-Poison Etoposide That Can Act in a TDP2-Independent Pathway. International Journal of Molecular Sciences. 2023; 24(7):6545. https://doi.org/10.3390/ijms24076545
Chicago/Turabian StylePark, Jeong-Min, Huimin Zhang, Litong Nie, Chao Wang, Min Huang, Xu Feng, Mengfan Tang, Zhen Chen, Yun Xiong, Namsoo Lee, and et al. 2023. "Genome-Wide CRISPR Screens Reveal ZATT as a Synthetic Lethal Target of TOP2-Poison Etoposide That Can Act in a TDP2-Independent Pathway" International Journal of Molecular Sciences 24, no. 7: 6545. https://doi.org/10.3390/ijms24076545