

Universal Stress Proteins: From Gene to Function

National Key Laboratory of Green Pesticide, Key Laboratory of Green Pesticide and Agricultural Bioengineering, Ministry of Education, Guizhou University, Guiyang 550025, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(5), 4725; https://doi.org/10.3390/ijms24054725
Submission received: 30 January 2023
/
Revised: 23 February 2023
/
Accepted: 23 February 2023
/
Published: 1 March 2023
(This article belongs to the Special Issue Advances in Horticultural Research Based on Multi-Omics and Computational Biology)
Abstract
:Universal stress proteins (USPs) exist across a wide range of species and are vital for survival under stressful conditions. Due to the increasingly harsh global environmental conditions, it is increasingly important to study the role of USPs in achieving stress tolerance. This review discusses the role of USPs in organisms from three aspects: (1) organisms generally have multiple USP genes that play specific roles at different developmental periods of the organism, and, due to their ubiquity, USPs can be used as an important indicator to study species evolution; (2) a comparison of the structures of USPs reveals that they generally bind ATP or its analogs at similar sequence positions, which may underlie the regulatory role of USPs; and (3) the functions of USPs in species are diverse, and are generally directly related to the stress tolerance. In microorganisms, USPs are associated with cell membrane formation, whereas in plants they may act as protein chaperones or RNA chaperones to help plants withstand stress at the molecular level and may also interact with other proteins to regulate normal plant activities. This review will provide directions for future research, focusing on USPs to provide clues for the development of stress-tolerant crop varieties and for the generation of novel green pesticide formulations in agriculture, and to better understand the evolution of drug resistance in pathogenic microorganisms in medicine.
1. Introduction
In 1992, Nyström and Neidhardt [1] identified a protein in Escherichia coli that was overexpressed under stressful conditions and named it Universal Stress Protein (USP) [2,3]. The USP was shown to play an important role in various [4] aspects of phosphorylation [5] and coordination of glucose [6] and acetate metabolism in E. coli [7]. After the amino acid sequence of the first USP was determined in E. coli [8], USPs were identified in a range of organisms [9,10]. The E. coli USP gene family includes UspA, UspC (YecG), UspD (YiiT), UspE (YdaA), UspF (YnaF, UP03) and UspG (YbdQ), and they have been divided into three subfamilies (Figure 1a) [11]. Various E. coli USPs play different roles in this bacterium [12] and can be divided into two major classes (Figure 1b) and four minor subclasses (Figure 1c) [13].
Follow-up studies have found that USP genes are present not only in E. coli [14], but also in the genomes of other bacteria [15], archaea, fungi, protozoa [10], and plants [16]. It was found that the USP genes are involved in the formation of biofilms [17] that help bacteria survive in an anaerobic environment [18]. Oryza sativa was the first eukaryote in which a USP protein was reported, and OsUSP1 was shown to be particularly closely related to the bacterial MJ0577-type ATP-binding USP protein, possibly playing a role in ethylene-mediated anaerobic stress adaptation in rice [19]. Subsequent studies [20] have detected USPs in a number of plant [21] species with expression upregulated in response to stresses [22] such as drought [23,24] or cold [25], as well as with plant root nodule formation [26]. Studies at the molecular level [27] have shown that USPs are associated with a range of functions, such as protection of nucleic acids, cellular defense [28], stress tolerance [29], protein scaffolding, and cellular protein transport [30]. The study of USPs in plants is not as in-depth as in microorganisms, and most of the studies on plant USPs focus on plant resistance studies, with few studies on structure and other aspects. In this paper, we summarize the studies on USPs from three aspects: genetic, structural and functional, in order to provide a reference for the study of USPs in plants.
2. Diversity of USP Genes
The Acinetobacter baumannii universal stress protein A (USPA) protects the bacterium, a major global public health threat, from H2O2 stress [31]. Schistosoma mansoni is the causative agent of human schistosomiasis, and all S. mansoni USP genes are transcribed during at least one developmental stage of the helminth life cycle, with the expression of six of these genes being upregulated in the trichocysts, a free-swimming developmental stage of S. mansoni that is essential for transmission to the intermediate snail host. After transmission into the intermediate host, the S. mansoni USP transcripts may be induced to perform specific functions triggered by environmental stressors [32].
USP genes can be an important marker for studying species evolution. Espinola et al. provided a broad framework for the evolution of the USP gene family to provide a basis for future studies on the emergence of USPs in other tax [33]. A recent study of the gene encoding the allosteric universal stress protein (USPA) in Halomonas spp. has demonstrated its role in microbial evolution, its presence suggesting that Halomonas spp. developed directly from primitive bacteria [34]. Therefore, the study of microbial USPs can help us understand the origins and mode of action of these proteins, which would be beneficial to the human exploitation of microbial resources.
The model plant Arabidopsis thaliana possesses as many as 41 USP genes (Table 1), which are extremely important in the developmental process of this plant [35] USP genes play specific roles at each developmental stage; different genes can also respond when the plants are subjected to different stresses [36]. For example, barley has HvUsp genes that are specifically expressed only in roots and leaves [37]. The AtUsp genes positively regulates genes encoding cell wall components, thus facilitating cell expansion during organ growth [38]. The AtUsp promoter can be highly induced by plant hormones and a number of abiotic stresses and can also be induced by other effectors to reduce the effect of stress on important traits in the plant [39]. The expression of At3g58450 is regulated by phytohormones; it is involved in A. thaliana seed germination and may also regulate the flowering process [36,40]. The expression of AtUsp17 can be induced by a number of stresses and this gene regulates salt tolerance in A. thaliana [41].
Studies targeting the 21 USPs in grape (Vitis vinifera) revealed that the VvUspA promoter contains potential hormone response and stress-related elements, suggesting that the VvUspA gene may be involved in various hormone and stress response pathways [42]. A full-length 678-bp cDNA fragment, containing a MsUsp 528-bp coding sequence, was identified within Medicago sativa (alfalfa) leaves, and this gene was found to be commonly expressed in leaves, stems, flowers, roots, rhizomes, and seeds, with the highest expression levels being in seeds and relatively low levels in flowers and roots [43]. The Solanum pennellii gene (SpUsp) is located on chromosome 1 with a length of 572 bp, including a 438-bp open reading frame (ORF), a 91-bp 5′-untranslated region, and a 43-bp 3′-untranslated region, and has been found to be a drought-responsive gene [44]. A total of 44 USP genes, ranging in length from 222 bp (OsUsp31) to 2817 bp (OsUsp20), have been identified in rice, located on 11 of the 12 chromosomes, with the exception of chromosome 4, and alternative splicing of the primary transcript has been recognized, enabling the synthesis of multiple proteins from a single gene [45]. Malus sieversii contains a MsUspA gene of length 222 bp, and its expression decreases reactive oxygen species (ROS) accumulation and enhances plant drought tolerance [46]. The analysis of 16 HvUsp genes in barley revealed that each gene contained two or three introns, which range in size from 75 to 941 bp, and expression is upregulated in response to salt stress [37]. There are 71 genes encoding USP-related structural domains in pigeonpea (Cajanus cajan) [47], 49 of which have been shown to be drought-responsive genes [48]. The GaUsp1 [49] gene in Gossypium arboreum is associated with drought response [50], whereas the GaUsp2 gene is associated with salt tolerance and osmotolerance [51]. The full-length sequence of SbUsp cDNA from the extreme halophyte Salicornia brachiata is 873 bp long, containing a 486-bp ORF, and the SbUsp gene promotes plant growth, reduces ROS accumulation, maintains ion homeostasis, and improves plant physiology under conditions of salt or osmotic stress [52]. This species was also found to contain two homologs of the intronless SbUsp gene, which encode salt- and osmosis-responsive USPs [53]. The medicinal plant Calotropis procera is a good model plant for the study of salt- and drought-tolerance genes, and the discovery of the USP gene of this plant is of great significance for the selection and breeding of anti-adverse varieties [54]. Salvia miltiorrhiza, a highly prized plant in traditional Chinese medicine, contains 32 cDNAs of USP family members, ranging in length from 279 bp (SmUSP11) to 2274 bp (SmUSP3) [55]. MaUsp genes identified from mulberry (Morus alba L.) are capable of conferring cellular-level tolerance in both prokaryotic and eukaryotic systems. This finding led to a new direction of thinking for subsequent USP studies, namely that maintaining cell-level tolerance is essential to maintaining growth under stress and that cell-level tolerance can be enhanced by overexpression of genes such as USPs [56].
3. USP Crystal Structure
Thomas et al. (1998) isolated and purified MJ0577 from the hyperthermophilic bacterium Methanococcus jannaschii, and resolved its protein crystal structure (1MJH, PDB DOI: 10.2210/pdb1MJH/pdb). The MJ0577 monomer consists of five-stranded twisted parallel β-sheet lamellae, with two on each side of the lamella helix, which can bind ATP, although the protein itself is not an ATPase, but exercises the function of an ATPase in physiological activities or as an ATP-mediated molecular switch in vivo [57]. Marcelo et al. (2001) resolved the crystal structure of the USPA of Haemophilus influenzae (1JMV, PDB DOI: 10.2210/pdb1JMV/pdb), which has a tertiary structure similar to that of the α/β-fold of MJ0577, but which does not bind ATP. The protein is present in solution as a dimer, with sulfate ions forming the dimer by chelating with the side chains through Arg28 and His29 [58]. Mycobacterium spp. stress protein MSMEG-3811 (5AHW, PDB DOI: 10.2210/pdb5AHW/pdb) binds cyclic adenosine monophosphate (cAMP), with the monomeric structure of this protein having a typical, open, twisted five-stranded parallel β-sheet layer with a topology of β3-β2-β1-β4-β5 sandwiched between six helices of different lengths, with a binding pocket that acts as a protein regulator of downstream effectors of cAMP-binding proteins, so that the abundance of this protein may also determine the amount of free cAMP in the cell [59]. In the same year, the crystal structures of YdaA (USPE, 4R2J, PDB DOI: 10.2210/pdb4R2J/pdb) and YnaF (USPF, 4R2L, 10.2210/pdb4R2L/pdb) of Salmonella typhimurium were resolved for the first time [60], and the monomer of YnaF was shown to consist of five chains (S1–S5) with the following chain order S3-S2-S1-S4-S5, the chain ends being connected by α-helices. The two YnaF plasmids form an asymmetric unit that is stabilized by interaction with chloride ions to form a tetramer with symmetry. YdaA is a larger protein than YnaF and the YdaA protomer includes an N-terminal structural domain (residues 1–149) and a C-terminal structural domain (residues 150–315) in addition to two tandem USP structural domains. The two protomers of YdaA combine to form a tetramer similar to the tetrameric structure of YnaF. To verify the functions of YdaA and YnaF, each of these two proteins was mutated and the crystal structures of the mutant proteins 4R2K (PDB DOI: 10.2210/pdb4R2K/pdb) and 4R2M (PDB DOI: 10.2210/pdb4R2M/pdb) were obtained [60].
NE1028 (3TNJ, PDB DOI: 10.2210/pdb3TNJ/pdb) from Nitrosomonas europaea has potential ATP-binding residues, and the structure of the complex formed by its binding to AMP was reported by Tkaczuk et al., who also reported that the mutant 2PFS (PDB DOI: 10.2210/pdb2PFS/pdb), namely the universal stress protein 3QTB complexed with dAMP from Archaeoglobus fulgidus(PDB DOI: 10.2210/pdb3QTB/pdb), and the universal stress protein 6HCD (PDB DOI: 10.2210/pdb6HCD/pdb) from archaea [30]. The histidine kinase KdpD in the KdpDE two-component system (TCS) contains a USP structural domain that binds to the second messenger cyclic diadenosine monophosphate (c-di-AMP) and is used to regulate the transcriptional output of the TCS in thick-walled bacterial taxa such as Staphylococcus aureus. Given this structure, Dutta et al., suggested that the USP structural domain in the KdpD histidine kinase may represent a new USP subfamily [61]. The crystal structure of E. coli USPE (5CB0, PDB DOI: 10.2210/pdb5CB0/pdb) was resolved [62], showing that USPE folds into a fan-shaped structure and has a hydrophobic cavity bound to its ligand [63]. The Arabidopsis protein At3g01520 (2GM3, PDB DOI: 10.2210/pdb2GM3/pdb) is the only eukaryotic universal stress protein crystal structure that has been resolved, and the structure shows that it is an aggregate and that each monomer is bound to an AMP molecule its [64].
The crystalline small molecules containing ATP or ATP analogues (AMP, ANP, etc.) were selected for sequence comparison and structure superposition, and the results are shown in Figure 2.
It is well known that structure determines function, and by combing through them we found that most USPs structures rarely exist as single chains when they are resolved, (Table 2) they are often aggregated in even chains, while ATP is involved in protein crystallization, which may be an important reason for its function in plants.
4. Functional Diversity of USPs
4.1. Functional Diversity of Prokaryotic USPs
USPs are important regulatory stress proteins which have been reported from a wide range of bacterial species [65], which help bacteria survive under conditions of stress. USPs have various functions in fine bacteria, such as ATP hydrolysis [66], modification of membrane properties [67] and chlorine sensing [68]. Mycobacterium tuberculosis expresses USPs to survive hypoxia and carbon monoxide stress, and it was found that this bacterium expresses ten USPs, which were divided into five classes (Figure 3a) [69]. Universal stress proteins from M. tuberculosis, such as Rv2623 and Rv2624c, bind ATP [27], whereas Rv1636 binds more to cAMP than to ATP [70]. Rv2623 regulates mycobacterial growth both in vivo and in vitro (interacting with Rv1747; Figure 3b), suggesting that it is an essential protein for M. tuberculosis during the chronic phase of host infection. It was found that the ATP-binding activity of Rv2623 determines the growth-regulating properties of the USP and that Rv2623 may act as an ATP-dependent signaling intermediate during persistent infection of the host [71]. Rv2624c alters the abundance of arginine in vivo by binding to ATP, to make M. tuberculosis more likely to survive in the host [72]. At the same time, overexpression of the universal stress protein BCG-2013 in M. tuberculosis, which is associated with latency, increases the bifunctional catalase-peroxidase KatG level, which makes the overexpressed strain more sensitive to isoniazid (INH) [73]. Species in the bacterial family Desulfovibrionaceae have 651 sequences associated with the expression of USPs, which can be divided into four classes based on the number of structural domains and ATP-binding motifs (Figure 3c) The USPs are associated with the survival of members of the Desulfovibrionaceae in anaerobic aquatic environments and also with cellular uptake of inorganic mercury and the production of methylmercury [74].
USPs may be a major regulator of bacterial survival, and it has been suggested that the USP of Micrococcus luteus may be a switch for metabolic control in this bacterium, with related studies suggesting that this USPA616 regulates glyoxylate shunting to make it more likely for the bacterium to survive under stressful conditions [65]. USP4207 from Mycobacterium smegmatis is closely involved in biofilm formation, with strains lacking USP4207 exhibiting reduced biofilm formation in vivo, resulting in coarser colony morphology [43]. The presence of USP76 in lung cells is a hallmark of Burkholderia infection [75]. Listeria innocua ATCC 33090 contains a novel ATP-binding USP that is upregulated not only during the stable phase but also during the exponential growth period, and it has been found to be involved in the in vivo bacterial response system when acid stress is encountered during the exponential period [76]. USPF is involved in the tolerance process of atypical enteropathogenic E. coli (aEPEC) and other Enterobacteriaceae [77].
USPs are associated not only with bacterial stress tolerance but also with the pathogenicity of pathogenic bacteria, and are important factors for the survival or persistence of various pathogens [78,79]. A. baumannii USPA plays an important role in the pathogenicity of the two most lethal, infectious diseases caused by this bacterial human pathogen, pneumonia and sepsis, and it is an essential component of the virulence mechanism of A. baumannii [31]. Edwardsiella piscicida USP13 is essential for pathogenicity and can help block the host immune response to pathogen infection [80]. USPC was found to function as a scaffold for signaling in the KdpD/KdpE-P/DNA complex [81] and to regulate the expression of the ion pump/channel complex kdpFABC [82], the N-terminal domain of YdaA binds zinc and might play a role in lipid metabolism [60].
4.2. Functional Diversity of Eukaryotic USPs
USPs also carry out multiple functions in eukaryotic organisms, and Schistosoma mansoni USPs may play a role [83] in defense against hydrogen peroxide-induced oxidative stress [32], so that USPs are now novel targets for human schistosomiasis intervention and treatment [84]. USPs, as important stress-regulated proteins in plants, are involved in a variety of physiological activities.
4.2.1. Versatility of A. thaliana USPs
There are 53 USPs in Arabidopsis, and these USPs are divided into four groups (Table 1). They are expressed in almost all parts of the plant and show tissue specificity and different functions at various developmental stages [35]. AT3G53990 has several possible functions in Arabidopsis. AT3G53990 transforms itself from a low molecular weight (LMW) complex to a high molecular weight (HMW) complex in response to high temperature scorching stress, and such a structural change could help plants protect key intracellular proteins in high temperature environments, and it would participate as a protein chaperone to play a stabilizing role as a protein [85] (Figure 4a). AT5G35380 is an effector of proline accumulation at low water potential [86]. The GRUSP protein, encoded by the AT3G58450 gene, is a novel regulatory component of the flowering signal transduction pathway in Arabidopsis, and overexpression of this protein interferes with the flowering signal, so that flowering is delayed, and a decrease in the concentration of the endogenous, bioactive gibberellins GA1 and GA3 is detected [40]. GRUSP is also associated with Arabidopsis seed germination and has a similar regulatory pattern to the hormone, but unlike abscisic acid, it promotes seed germination [36]. HRU1 (Hypoxia-responsive universal stress protein) coordinates oxygen sensing and ROS signaling under hypoxic conditions [87]. AtUSP17 negatively mediates salt tolerance in Arabidopsis by regulating ethylene, ABA, ROS, and G-protein signaling and responses [41].
As important stress-regulated proteins, USPs are involved in a range of physiological activities in plants. It has been shown that cold-shock tolerance is usually achieved by RNA chaperones [88], and that, in Arabidopsis, AtUSP becomes involved in plant physiological activities as an RNA chaperone, which helps RNA-bound proteins to exercise their functions correctly by preventing RNA misfolding or by resolving misfolded RNAs [89] (Figure 4b). Meanwhile, when subjected to environmental stress, the expression of USPs causes upregulation of the expression of secondary metabolites in plants, resulting in an increase in intracytoplasmic solutes and achieving a greater tolerance to stress [46]. More importantly, USPs can also reduce ROS production in plants [90], maintain ROS homeostasis, alleviate oxidative damage caused by ROS, and improve tolerance to oxidative stress [91]. Interestingly, Arabidopsis AtUSP (At3g53990) exhibits anti-fungal activity by generating ROS and causing mitochondrial damage in the pathogenic fungi [92]. This suggests that the study of plant USPs could be beneficial for the selection of disease-resistant varieties and the creation of novel pesticides.
Plants have evolved complex redox signaling [91] regulatory systems [93] and can tolerate stress by activating specific intracellular redox-mediated defense signaling pathways [94,95]. The plant redox system was found to regulate ROS concentration in plants in relation to cell metabolism [96,97], apoptosis [98,99], and carbon metabolism and photosynthesis-related processes [100,101]. It has been shown that three Arabidopsis USP proteins, HRU1, AtUSP, and At3g17020, interact with their redox chaperone thioredoxin-h1, and that the structural transition of AtUSPs is induced by external redox changes, accompanied by changes in their function.
4.2.2. Functional Diversity of USPs in Other Plants
The tomato USPA (SlRd2) is an ATP-binding protein that forms homodimers in plants. SlRd2 is a novel interactor and phosphorylation target of SlCipk6, a member of the CIPK (CBL-interacting protein kinase) family, and functionally regulates SlCipk6-mediated ROS generation [102]. Under drought conditions, overexpression of the S. pennellii gene (SpUsp) causes upregulation of a large number of chlorophyll a- and b-binding proteins and photosystem proteins in plants, increasing the ABA concentration, closing stomata, alleviating oxidative damage caused by ROS, and improving tolerance to oxidative stress [44].
The expression and regulation of Gossypium arboretum GUSP1 under drought stress results in drought tolerance in this plant [50]. It has been demonstrated that, following the induction of drought tolerance, the relative water content, total chlorophyll content, CO2 absorbed by net photosynthesis, stomatal conductance, total soluble sugars, and proline concentration of the plant’s leaves increased significantly whereas the relative membrane permeability and transpiration rate decreased significantly, suggesting that GUSP1 may activate some downstream genes in signal transduction pathways in response to drought or other abiotic stresses in order to protect membranes and cells from damage [49]. The results of the mutant assay of heterologous expression of GUSP-2 with a lysine-to-threonine mutation demonstrate that this protein may be directly involved in stress tolerance or may act as a signaling molecule to activate stress adaptation mechanisms [51]. A universal stress protein in M. sieversii (MsUSPA) is involved in the regulation of hormone and secondary metabolite synthesis to reduce transpiration and retain water by altering the cellular structure of leaves to improve drought resistance. In addition, overexpression of MsUspA increases the activity of antioxidant enzymes and improves antioxidant capacity, reducing the accumulation of ROS [46].
Of the 32 members of the S. miltiorrhiza SmUSP protein family, four target mitochondria, four target chloroplasts, and two specifically compete for the secretory pathway [55]. The S. brachiata SbUSP is a membrane-bound cytoplasmic protein that interacts with AMP and exhibits characteristic phosphorylation and glycosylation motifs and ATP-binding sites, suggesting that SbUSP may be directly involved in tolerance mechanisms or act as a molecular switch (signaling molecule) to activate stress adaptation mechanisms [53]. The archaea Sulfolobus acidocaldarius contains SaUSPA, which is an ATP-binding protein that binds to the phosphatase PP2A in vitro and in vivo. Although SaUSPA does not hydrolyze ATP, it stimulates the phosphatase activity of PP2A and may affect many other processes in this way [103]. The leguminous plant Astragalus fridae increased the expression rate of USP in response to exposure to SiO2 nanoparticles [104].
5. Future Perspectives
It is clear that the study of USPs has moved from superficial investigations to in-depth examination of the mechanisms involved. In the future, the study of USPs may focus on the following trends: (1) despite their importance, our understanding of the structure of USPs is still insufficiently detailed, particularly that of eukaryotic USPs; (2) although USPs are able to help cells survive under stressful conditions, these proteins follow two completely different mechanisms, namely ATP-dependent USPs play a role in cellular transport, whereas ATP-non-binding USPs may function in the cell cycle [105]; (3) the application of USP research to agriculture may facilitate the development of stress-tolerant crop varieties, and the creation of new green pesticide formulations, using USPs as targets or lead compounds for therapeutic or protective agents; and (4) in the medical field, the study of USPs in drug-resistance mechanisms in pathogenic bacteria is important for the screening of novel therapeutic drugs [79] and the development of related antibodies and vaccines for the prevention and treatment of human diseases. USPs from different plants, which play different roles in vivo (Table 3), are also worthy of research in the future by genetically engineering plants to adapt them to more complex climatic challenges.
Author Contributions
Conceptualization, X.L.; data curation, D.L. and Z.W.; formal analysis, D.L. and Q.B.; funding acquisition, X.L.; investigation, D.L.; methodology, D.L. and Z.W.; project administration, D.L.; resources, X.L.; software, Z.W.; supervision, X.L.; validation, X.L.; visualization, D.L. and Z.W.; writing—original draft, D.L. and Z.W.; writing—review and editing, X.L., Q.B., Y.Z., M.H. and Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the National Key Research and Development Program of China (Grant No. 2022YFD1400200) and National Natural Science Foundation of China (Grant No. 32172461), Talents of Guizhou Science and Technology Cooperation Platform [Grant No. (2021)5623], Guizhou Science and Technology Cooperation Foundation [Grant No. ZK(2021)140], The Unique Feature Project of Guizhou Provincial Education Department [Grant No. KY(2021)056], 17Program of Introducing Talents of Discipline to Universities of China (Grant No. 111 Program, D20023).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare that they have no known competing financial interest in this work.
References
- Nyström, T.; Neidhardt, F.C. Cloning, mapping and nucleotide sequencing of a gene encoding a universal stress protein in Eschericha coli. Mol. Microbiol. 1992, 6, 3187–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyström, T.; Neidhardt, F.C. Isolation and properties of a mutant of Escherichia coli with an insertional inactivation of the uspA gene, which encodes a universal stress protein. J. Bacteriol. 1993, 175, 3949–3956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyström, T.; Neidhardt, F.C. Expression and role of the universal stress protein, UspA, of Escherichia coli during growth arrest. Mol. Microbiol. 1994, 11, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freestone, P.; Nystrom, T.; Trinei, M.; Norris, V. The universal stress protein, UspA, of Escherichia coli is phosphorylated in response to stasis. J. Mol. Biol. 1997, 274, 318–324. [Google Scholar] [CrossRef]
- Freestone, P.; Grant, S.; Trinei, M.; Onoda, T.; Norris, V. Protein phosphorylation in Escherichia cob L. form NC=7. Microbiology 1998, 144, 3289–3295. [Google Scholar] [CrossRef] [Green Version]
- Phadtare, S.; Inouye, M. Role of CspC and CspE in regulation of expression of RpoS and UspA, the stress response proteins in Escherichia coli. J. Bacteriol. 2001, 183, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.; Bausch, C.; Richmond, C.; Blattner, F.R.; Conway, T. Functional genomics: Expression analysis of Escherichia coli growing on minimal and rich media. J. Bacteriol. 1999, 181, 6425–6440. [Google Scholar] [CrossRef] [Green Version]
- Forêt, S.; Seneca, F.; De Jong, D.; Bieller, A.; Hemmrich, G.; Augustin, R.; Hayward, D.C.; Ball, E.E.; Bosch, T.C.; Agata, K. Phylogenomics reveals an anomalous distribution of USP genes in metazoans. Mol. Biol. Evol. 2011, 28, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Kvint, K.; Nachin, L.; Diez, A.; Nyström, T. The bacterial universal stress protein: Function and regulation. Curr. Opin. Microbiol. 2003, 6, 140–145. [Google Scholar] [CrossRef]
- Vollmer, A.C.; Bark, S.J. Twenty five years of investigating the universal stress protein: Function, structure, and applications. Adv. Appl. Microbiol. 2018, 102, 1–36. [Google Scholar]
- Gustavsson, N.; Diez, A.; Nyström, T. The universal stress protein paralogues of Escherichia coli are coordinately regulated and cooperate in the defence against DNA damage. Mol. Microbiol. 2002, 43, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Anantharaman, V.; Koonin, E.V. Monophyly of class I aminoacyl tRNA synthetase, USPA, ETFP, photolyase, and PP-ATPase nucleotide binding domains: Implications for protein evolution in the RNA world. Proteins Struct. Funct. Bioinform. 2002, 48, 1–14. [Google Scholar] [CrossRef]
- Nachin, L.; Nannmark, U.; Nystrom, T. Differential roles of the universal stress proteins of Escherichia coli in oxidative stress resistance, adhesion, and motility. J. Bacteriol. 2005, 187, 6265–6272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegele, D.A. Universal stress proteins in Escherichia coli. J. Bacteriol. 2005, 187, 6253–6254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boes, N.; Schreiber, K.; Härtig, E.; Jaensch, L.; Schobert, M. The Pseudomonas aeruginosa universal stress protein PA4352 is essential for surviving anaerobic energy stress. J. Bacteriol. 2006, 188, 6529–6538. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.X.; Wei, X.Y.; Chen, D.S.; Zhou, J.C. A novel nodule-enhanced gene encoding a putative universal stress protein from Astragalus sinicus. J. Plant Physiol. 2007, 164, 764–772. [Google Scholar] [CrossRef]
- Galindo Blaha, C.A.; Schrank, I.S. An Azospirillum brasilense Tn5 mutant with modified stress response and impaired in flocculation. Antonie Van Leeuwenhoek 2003, 83, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Fink, R.C.; Evans, M.R.; Porwollik, S.; Vazquez-Torres, A.; Jones-Carson, J.; Troxell, B.; Libby, S.J.; McClelland, M.; Hassan, H.M. FNR is a global regulator of virulence and anaerobic metabolism in Salmonella enterica serovar Typhimurium (ATCC 14028s). J. Bacteriol. 2007, 189, 2262–2273. [Google Scholar] [CrossRef] [Green Version]
- Sauter, M.; Rzewuski, G.; Marwedel, T.; Lorbiecke, R. The novel ethylene regulated gene OsUsp1 from rice encodes a member of a plant protein family related to prokaryotic universal stress proteins. J. Exp. Bot. 2002, 53, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Coetzer, N.; Gazendam, I.; Oelofse, D.; Berger, D.K. SSHscreen and SSHdb, generic software for microarray based gene discovery: Application to the stress response in cowpea. Plant Methods 2010, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Kerk, D.; Bulgrien, J.; Smith, D.W.; Gribskov, M. Arabidopsis proteins containing similarity to the universal stress protein domain of bacteria. Plant Physiol. 2003, 131, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Merkouropoulos, G.; Andreasson, E.; Hess, D.; Boller, T.; Peck, S.C. An Arabidopsis protein phosphorylated in response to microbial elicitation, AtPHOS32, is a substrate of MAP kinases 3 and 6. J. Biol. Chem. 2008, 283, 10493–10499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maqbool, A.; Zahur, M.; Irfan, M.; Younas, M.; Barozai, K.; Rashid, B.; Husnain, T.; Riazuddin, S. Identification and expression of six drought-responsive transcripts through differential display in desi cotton (Gossypium arboreum). Mol. Biol. 2008, 42, 492–498. [Google Scholar] [CrossRef]
- Wang, H. Construction and Analysis of subtractive cDNA library of phellodendron amurense under drought stress. Chin. J. Biotechnol. 2008, 24, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Sapitnitskaya, M.; Maul, P.; McCollum, G.T.; Guy, C.L.; Weiss, B.; Samach, A.; Porat, R. Postharvest heat and conditioning treatments activate different molecular responses and reduce chilling injuries in grapefruit. J. Exp. Bot. 2006, 57, 2943–2953. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.A.; Raymond, M.J.; Smirnoff, N. Analysis of the root-hair morphogenesis transcriptome reveals the molecular identity of six genes with roles in root-hair development in Arabidopsis. Plant J. 2006, 45, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Drumm, J.E.; Mi, K.; Bilder, P.; Sun, M.; Lim, J.; Bielefeldt-Ohmann, H.; Basaraba, R.; So, M.; Zhu, G.; Tufariello, J.M. Mycobacterium tuberculosis universal stress protein Rv2623 regulates bacillary growth by ATP-Binding: Requirement for establishing chronic persistent infection. PLoS Pathog. 2009, 5, e1000460. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, M.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, S.; Rai, R.; Chatterjee, A.; Rai, S.; Yadav, S.; Agrawal, C.; Rai, L. Molecular characterization of two novel proteins All1122 and Alr0750 of Anabaena PCC 7120 conferring tolerance to multiple abiotic stresses in Escherichia coli. Gene 2019, 685, 230–241. [Google Scholar] [CrossRef]
- Tkaczuk, K.L.; Shumilin, I.A.; Chruszcz, M.; Evdokimova, E.; Savchenko, A.; Minor, W. Structural and functional insight into the universal stress protein family. Evol. Appl. 2013, 6, 434–449. [Google Scholar] [CrossRef]
- Elhosseiny, N.M.; Amin, M.A.; Yassin, A.S.; Attia, A.S. Acinetobacter baumannii universal stress protein A plays a pivotal role in stress response and is essential for pneumonia and sepsis pathogenesis. Int. J. Med. Microbiol. 2015, 305, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; Mahmud, O.; Mbah, A.N.; Simmons, S.S.; Brown, S.D. Developmental regulation of genes encoding universal stress proteins in Schistosoma mansoni. Gene Regul. Syst. Biol. 2011, 5, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinola, S.M.; Cancela, M.P.; Corrêa, L.B.; Zaha, A. Evolutionary fates of universal stress protein paralogs in Platyhelminthes. BMC Evol. Biol. 2018, 18, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekadwad, B.N.; Li, W.J.; Rekha, P.D. The diversity of unique 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxylic acid coding common genes and Universal stress protein in Ectoine TRAP cluster (UspA) in 32 Halomonas species. BMC Res. Notes 2021, 14, 296. [Google Scholar] [CrossRef] [PubMed]
- Bhuria, M.; Goel, P.; Kumar, S.; Singh, A.K. Genome-wide identification and expression profiling of genes encoding universal stress proteins (USP) identify multi-stress responsive USP genes in Arabidopsis thaliana. Plant Physiol. Rep. 2019, 24, 434–445. [Google Scholar] [CrossRef]
- Gorshkova, D.; Getman, I.; Voronkov, A.; Chizhova, S.; Kuznetsov, V.V.; Pojidaeva, E. The gene encoding the universal stress protein AtUSP is regulated by phytohormones and involved in seed germination of Arabidopsis thaliana. Dokl. Biochem. Biophys. 2018, 479, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; Wei, Y.M.; Wang, J.R.; Liu, C.J.; Lan, X.J.; Jiang, Q.T.; Pu, Z.E.; Zheng, Y.L. Identification, localization, and characterization of putative USP genes in barley. Theor. Appl. Genet. 2010, 121, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Phan, K.A.T.; Jun, S.E.; Park, J.H.; Paeng, S.K.; Chae, H.B.; Wi, S.D.; Bae, S.B.; Kang, K.R.; Kim, G.T.; et al. Universal Stress Protein (USP) enhances plant growth and development by promoting cell expansion. J. Plant Biol. 2022, 65, 231–239. [Google Scholar] [CrossRef]
- Bhuria, M.; Goel, P.; Kumar, S.; Singh, A.K. The promoter of AtUSP is coregulated by phytohormones and abiotic stresses in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1957. [Google Scholar] [CrossRef] [Green Version]
- Gorshkova, D.; Getman, I.; Sergeeva, L.; Kuznetsov, V.V.; Pojidaeva, E. GRUSP, an universal stress protein, is involved in gibberellin-dependent induction of flowering in Arabidopsis thaliana. Dokl. Biochem. Biophys. 2021, 499, 233–237. [Google Scholar] [CrossRef]
- Bhuria, M.; Goel, P.; Kumar, S.; Singh, A.K. AtUSP17 negatively regulates salt stress tolerance through modulation of multiple signaling pathways in Arabidopsis. Physiol. Plant. 2022, 174, e13635. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhang, P.; Hu, Y.; Chen, C.; Liu, Q.; Guan, P.; Zhang, J. Genome-wide analysis of the universal stress protein A gene family in Vitis and expression in response to abiotic stress. Plant Physiol. Biochem. 2021, 165, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Biswas, P.; Banerjee, A.; Bose, A.; Siddiqui, N.; Nambi, S.; Saini, D.K.; Visweswariah, S.S. A universal stress protein in Mycobacterium smegmatis sequesters the cAMP-regulated lysine acyltransferase and is essential for biofilm formation. J. Biol. Chem. 2020, 295, 1500–1516. [Google Scholar] [CrossRef] [Green Version]
- Loukehaich, R.; Wang, T.; Ouyang, B.; Ziaf, K.; Li, H.; Zhang, J.; Lu, Y.; Ye, Z. SpUSP, an annexin-interacting universal stress protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arabia, S.; Sami, A.A.; Akhter, S.; Sarker, R.H.; Islam, T. Comprehensive in silico characterization of universal stress proteins in rice (Oryza sativa L.) with insight into their stress-specific transcriptional modulation. Front. Plant Sci. 2021, 12, 712607. [Google Scholar] [CrossRef]
- Yang, M.; Che, S.; Zhang, Y.; Wang, H.; Wei, T.; Yan, G.; Song, W.; Yu, W. Universal stress protein in Malus sieversii confers enhanced drought tolerance. J. Plant Res. 2019, 132, 825–837. [Google Scholar] [CrossRef]
- Varshney, R.K.; Chen, W.; Li, Y.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.; Azam, S.; Fan, G.; Whaley, A.M. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2012, 30, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Sinha, P.; Pazhamala, L.T.; Singh, V.K.; Saxena, R.K.; Krishnamurthy, L.; Azam, S.; Khan, A.W.; Varshney, R.K. Identification and validation of selected universal stress protein domain containing drought responsive genes in pigeonpea (Cajanus cajan L.). Front. Plant Sci. 2016, 6, 1065. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.; Ahmad, A.; Batool, F.; Rashid, B.; Husnain, T. Genetic modification of Gossypium arboreum universal stress protein (GUSP1) improves drought tolerance in transgenic cotton (Gossypium hirsutum). Physiol. Mol. Biol. Plants 2021, 27, 1779–1794. [Google Scholar] [CrossRef]
- Hassan, S.; Samiullah, T.R.; Ansari, M.R.; Rashid, B.; Husnain, T. Cloning, antibody production, expression and cellular localization of universal stress protein gene (USP1-GFP) in transgenic cotton. J. Plant Biochem. Biotechnol. 2018, 27, 175–185. [Google Scholar] [CrossRef]
- Hafeez, M.N.; Khan, M.A.; Sarwar, B.; Hassan, S.; Ali, Q.; Husnain, T.; Rashid, B. Mutant Gossypium universal stress protein-2 (GUSP-2) gene confers resistance to various abiotic stresses in E. coli BL-21 and CIM-496-Gossypium hirsutum. Sci. Rep. 2021, 11, 20466. [Google Scholar] [CrossRef] [PubMed]
- Udawat, P.; Jha, R.K.; Sinha, D.; Mishra, A.; Jha, B. Overexpression of a cytosolic abiotic stress responsive universal stress protein (SbUSP) mitigates salt and osmotic stress in transgenic tobacco plants. Front. Plant Sci. 2016, 7, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udawat, P.; Mishra, A.; Jha, B. Heterologous expression of an uncharacterized universal stress protein gene (SbUSP) from the extreme halophyte, Salicornia brachiata, which confers salt and osmotic tolerance to E. coli. Gene 2014, 536, 163–170. [Google Scholar] [CrossRef]
- Shokry, A.M.; Karim, S.A.; Ramadan, A.; Gadallah, N.; Attas, S.G.; Sabir, J.S.; Hassan, S.M.; Madkour, M.A.; Bressan, R.; Mahfouz, M. Detection of a Usp-like gene in Calotropis procera plant from the de novo assembled genome contigs of the high-throughput sequencing dataset. C. R. Biol. 2014, 337, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Su, J.; Na, Y.; Hui, Z.; Cao, X.Y.; Kang, J.F. Functional characterization of selected universal stress protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli. Genes 2017, 8, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanyalakshmi, K.H.; Nataraja, K.N. Universal stress protein-like gene from mulberry enhances abiotic stress tolerance in Escherichia coli and transgenic tobacco cells. Plant Biol. 2021, 23, 1190–1194. [Google Scholar] [CrossRef]
- Zarembinski, T.I.; Hung, L.W.; Mueller, H.J.; Kim, K.K.; Yokota, H.; Kim, R.; Kim, S.H. Structure-based assignment of the biochemical function of a hypothetical protein: A test case of structural genomics. Proc. Natl. Acad. Sci. USA 1998, 95, 15189–15193. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.C.; McKay, D.B. Structure of the Universal Stress Protein of Haemophilus influenzae. Structure 2001, 9, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Adolph, R.S.; Gopalakrishnapai, J.; Kleinboelting, S.; Emmerich, C.; Steegborn, C.; Visweswariah, S.S. A universal stress protein (USP) in Mycobacteria binds cAMP. J. Biol. Chem. 2015, 290, 12731–12743. [Google Scholar] [CrossRef] [Green Version]
- Bangera, M.; Panigrahi, R.; Sagurthi, S.R.; Savithri, H.S.; Murthy, M. Structural and functional analysis of two universal stress proteins YdaA and YnaF from Salmonella typhimurium: Possible roles in microbial stress tolerance. J. Struct. Biol. 2015, 189, 238–250. [Google Scholar] [CrossRef]
- Dutta, A.; Batish, M.; Parashar, V. Structural basis of KdpD histidine kinase binding to the second messenger c-di-AMP. J. Biol. Chem. 2021, 296, 100771. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Quan, C.-S.; Jin, X.; Zhao, J.; Li, X.; Zheng, W.; Jin, L.; Liu, D.; Fan, S.; Ha, N.-C. Crystallization and preliminary X-ray diffraction analysis of UspE from Escherichia coli. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 1640–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Guo, J.; Jin, X.; Kim, J.S.; Ji, Y.; Fan, S.; Ha, N.C.; Quan, C.S. Crystal structure and functional implications of the tandem-type universal stress protein UspE from Escherichia coli. BMC Struct. Biol. 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.; Eduard, K.; Craig, B.; Bingman, A.; Kim, H.J. Crystal structure of the protein At3g01520, a eukaryotic universal stress protein-like protein from Arabidopsis thalianain complex with AMP. Proteins Struct. Funct. Bioinform. 2015, 83, 1368–1373. [Google Scholar]
- Havis, S.; Bodunrin, A.; Rangel, J.; Zimmerer, R.; Murphy, J.; Storey, J.D.; Duong, T.D.; Mistretta, B.; Gunaratne, P.; Widger, W.R. A universal stress protein that controls bacterial stress survival in Micrococcus luteus. J. Bacteriol. 2019, 201, e00497-19. [Google Scholar] [CrossRef]
- Iino, H.; Shimizu, N.; Goto, M.; Ebihara, A.; Fukui, K.; Hirotsu, K.; Kuramitsu, S. Crystal structure of the tandem-type universal stress protein TTHA0350 from Thermus thermophilus HB8. J. Biochem. 2011, 150, 295–302. [Google Scholar] [CrossRef]
- Averaimo, S.; Milton, R.H.; Duchen, M.R.; Mazzanti, M. Chloride intracellular channel 1 (CLIC1): Sensor and effector during oxidative stress. FEBS Lett. 2010, 584, 2076–2084. [Google Scholar] [CrossRef] [Green Version]
- Suh, K.S.; Mutoh, M.; Nagashima, K.; Fernandez-Salas, E.; Edwards, L.E.; Hayes, D.D.; Crutchley, J.M.; Marin, K.G.; Dumont, R.A.; Levy, J.M. The organellular chloride channel protein CLIC4/mtCLIC translocates to the nucleus in response to cellular stress and accelerates apoptosis. J. Biol. Chem. 2004, 279, 4632–4641. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, R.; Williams, H.D. Universal stress proteins and Mycobacterium tuberculosis. Res. Microbiol. 2003, 154, 387–392. [Google Scholar] [CrossRef]
- Chakraborti, S.; Chakraborty, M.; Bose, A.; Srinivasan, N.; Visweswariah, S.S. Identification of potential binders of Mtb universal stress protein (Rv1636) through an in silico approach and insights into compound selection for experimental validation. Front. Mol. Biosci. 2021, 8, 177. [Google Scholar] [CrossRef]
- Glass, L.N.; Swapna, G.; Chavadi, S.S.; Tufariello, J.M.; Mi, K.; Drumm, J.E.; Lam, T.T.; Zhu, G.; Zhan, C.; Vilcheze, C. Mycobacterium tuberculosis universal stress protein Rv2623 interacts with the putative ATP binding cassette (ABC) transporter Rv1747 to regulate mycobacterial growth. PLoS Pathog. 2017, 13, e1006515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Q.; Hu, X.; Shi, D.; Zhang, Y.; Sun, M.; Wang, J.; Mi, K.; Zhu, G. Universal stress protein Rv2624c alters abundance of arginine and enhances intracellular survival by ATP binding in mycobacteria. Sci. Rep. 2016, 6, 35462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Li, X.; Huang, L.; Chan, J.; Chen, Y.; Deng, H.; Mi, K. Quantitative proteomics reveals novel insights into isoniazid susceptibility in mycobacteria mediated by a universal stress protein. J. Proteome Res. 2015, 14, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; McInnis, D.S.; Destefano, A.M.; Johnson, G.S.; Walker, A.D.; Hall, Y.A.; Mapp, B.W.; Johnson, M.O.; Simmons, S.S. Bioinformatics investigations of universal stressproteins from mercury methylating desulfovibrionaceae. Microorganisms 2021, 9, 1780. [Google Scholar] [CrossRef]
- O’Connor, A.; Berisio, R.; Lucey, M.; Schaffer, K.; Clean, S.M. A Universal Stress Protein upregulated by hypoxia may contribute to chronic lung colonisation and intramacrophage survival in cystic fibrosis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Tremonte, P.; Succi, M.; Coppola, R.; Sorrentino, E.; Tipaldi, L.; Picariello, G.; Pannella, G.; Fraternali, F. Homology-based modeling of universal stress protein from Listeria innocua up-regulated under acid stress conditions. Front. Microbiol. 2016, 7, 1998. [Google Scholar] [CrossRef]
- De Souza, C.S.; Torres, A.G.; Caravelli, A.; Silva, A.; Polatto, J.M.; Piazza, R.M.F. Characterization of the universal stress protein F from atypical enteropathogenic Escherichia coli and its prevalence in Enterobacteriaceae. Protein Sci. 2016, 25, 2142–2151. [Google Scholar] [CrossRef] [Green Version]
- Masamba, P.; Kappo, A.P. Parasite survival and disease persistence in cystic fibrosis, schistosomiasis and pathogenic bacterial diseases: A role for universal Stress Proteins? Int. J. Mol. Sci. 2021, 22, 10878. [Google Scholar] [CrossRef]
- Bandyopadhyay, D.; Singh, G.; Mukherjee, M.; Akhter, Y. Computational approach towards the design of novel inhibitor against universal stress protein A to combat multidrug resistant uropathogenic Escherichia coli. J. Mol. Struct. 2021, 1238, 130379. [Google Scholar] [CrossRef]
- Fang, Q.j.; Han, Y.X.; Shi, Y.J.; Huang, H.Q.; Fang, Z.G.; Hu, Y.H. Universal stress proteins contribute Edwardsiella piscicida adversity resistance and pathogenicity and promote blocking host immune response. Fish Shellfish. Immunol. 2019, 95, 248–258. [Google Scholar] [CrossRef]
- Heermann, R.; Weber, A.; Mayer, B.; Ott, M.; Hauser, E.; Gabriel, G.; Pirch, T.; Jung, K. The universal stress protein UspC scaffolds the KdpD/KdpE signaling cascade of Escherichia coli under salt stress. J. Mol. Biol. 2009, 386, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Heermann, R.; Jung, K. The complexity of the ‘simple’two-component system KdpD/KdpE in Escherichia coli. FEMS Microbiol. Lett. 2010, 304, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isokpehi, R.D.; Mahmud, O.; Awofolu, O.R.; Mbah, A.N. Inferences on the biochemical and environmental regulation of Universal Stress Proteins from Schistosomiasis parasites. Adv. Appl. Bioinform. Chem. 2013, 6, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masamba, P.; Adenowo, A.F.; Oyinloye, B.E.; Kappo, A.P. Universal stress proteins as new targets for environmental and therapeutic interventions of Schistosomiasis. Int. J. Environ. Res. Public Health 2016, 13, 972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.J.; Melencion, S.M.; Lee, E.S.; Park, J.H.; Alinapon, C.V.; Oh, H.T.; Yun, D.J.; Chi, Y.H.; Lee, S.Y. Universal stress protein exhibits a redox-dependent chaperone function in arabidopsis and enhances plant tolerance to heat shock and oxidative stress. Front. Plant Sci. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verslues, P.E.; Lasky, J.R.; Juenger, T.E.; Liu, T.W.; Kumar, M.N. Genome-wide association mapping combined with reverse genetics identifies new effectors of low water potential-induced proline accumulation in Arabidopsis. Plant Physiol. 2014, 164, 144–159. [Google Scholar] [CrossRef] [Green Version]
- Gonzali, S.; Loreti, E.; Cardarelli, F.; Novi, G.; Parlanti, S.; Pucciariello, C.; Bassolino, L.; Banti, V.; Licausi, F.; Perata, P. Universal stress protein HRU1 mediates ROS homeostasis under anoxia. Nat. Plants 2015, 1, 15151. [Google Scholar] [CrossRef] [PubMed]
- Melencion, S.M.B.; Chi, Y.H.; Pham, T.T.; Paeng, S.K.; Wi, S.D.; Lee, C.; Ryu, S.W.; Koo, S.S.; Lee, S.Y. RNA chaperone function of a universal stress protein in Arabidopsis confers enhanced cold stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Park, S.J.; Kwak, K.J. Plant RNA chaperones in stress response. Trends Plant Sci. 2013, 18, 100–106. [Google Scholar] [CrossRef]
- Chi, Y.H.; Koo, S.S.; Oh, H.T.; Lee, E.S.; Park, J.H.; Phan, K.A.T.; Wi, S.D.; Bae, S.B.; Paeng, S.K.; Chae, H.B. The physiological functions of universal stress proteins and their molecular mechanism to protect plants from environmental stresses. Front. Plant Sci. 2019, 10, 750. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.H.; Paeng, S.K.; Kim, M.J.; Hwang, G.Y.; Melencion, S.M.; Oh, H.T.; Lee, S.Y. Redox-dependent functional switching of plant proteins accompanying with their structural changes. Front. Plant Sci. 2013, 4, 277. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Jung, Y.J.; Lee, Y.; Kim, I.R.; Seol, M.-A.; Kim, E.-J.; Jang, M.-K.; Lee, J.R. Functional characterization of the Arabidopsis universal stress protein AtUSP with an antifungal activity. Biochem. Biophys. Res. Commun. 2017, 486, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Thormählen, I.; Daloso, D.M.; Fernie, A.R. The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Bosch, C.G. Priming plant resistance by activation of redox-sensitive genes. Free. Radic. Biol. Med. 2018, 122, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 8, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Mata-Pérez, C.; Spoel, S.H. Thioredoxin-mediated redox signalling in plant immunity. Plant Sci. 2019, 279, 27–33. [Google Scholar] [CrossRef]
- Hinoux, V.D.; Bangash, S.A.; Meyer, A.J.; Reichheld, J.P. Nuclear thiol redox systems in plants. Plant Sci. 2016, 243, 84–95. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Gutsche, N.; Thurow, C.; Zachgo, S.; Gatz, C. Plant specific CC-type glutaredoxins: Functions in developmental processes and stress responses. Biol. Chem. 2015, 396, 495–509. [Google Scholar] [CrossRef]
- Nikkanen, L.; Toivola, J.; Diaz, M.G.; Rintamäki, E. Chloroplast thioredoxin systems: Prospects for improving photosynthesis. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160474. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, E.G.; Personat, J.M.; Torre, F.D.; Pozo, O.D. A universal stress protein involved in oxidative stress is a phosphorylation target for protein kinase CIPK6. Plant Physiol. 2017, 173, 836–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Does, C.V.; Albers, S.V. SaUspA, the universal stress protein of Sulfolobus acidocaldarius stimulates the activity of the PP2A phosphatase and is involved in growth at high salinity. Front. Microbiol. 2020, 11, 2872. [Google Scholar] [CrossRef] [PubMed]
- Moghanloo, M.; Iranbakhsh, A.; Ebadi, M.; Ardebili, Z.O. Differential physiology and expression of phenylalanine ammonia lyase (PAL) and universal stress protein (USP) in the endangered species Astragalus fridae following seed priming with cold plasma and manipulation of culture medium with silica nanoparticles. 3 Biotech 2019, 9, 288. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, D.; Mukherjee, M. Systematic comparison of the protein-protein interaction network of bacterial universal stress protein A (UspA): An insight into its discrete functions. Biologia 2022, 77, 2631–2642. [Google Scholar] [CrossRef]
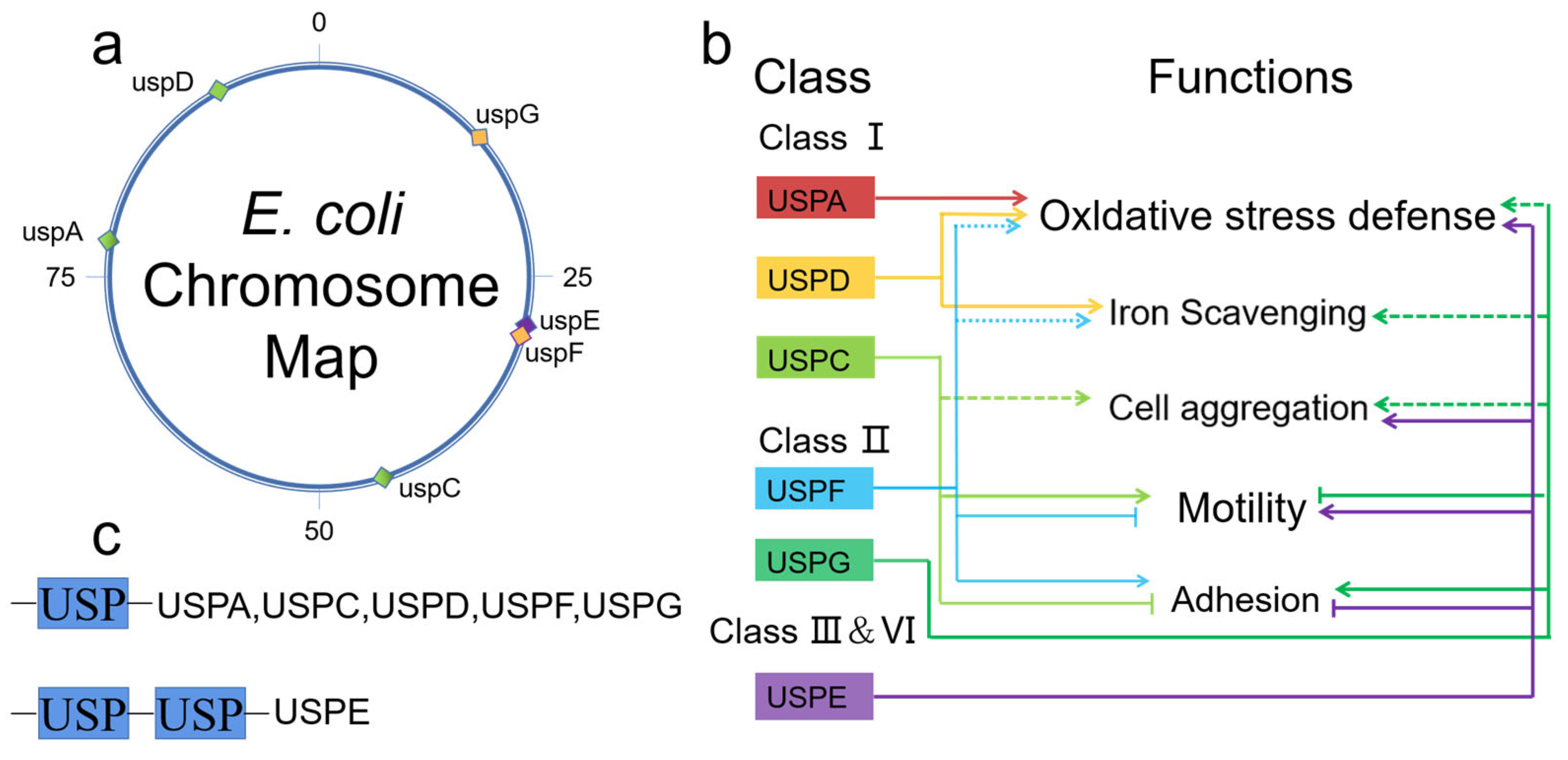
Figure 1.
E. coli genes and functions. (a) Location of USP family genes on E. coli chromosomes. uspA, uspC and uspD belong to a category represented by green squares, uspF and uspG belong to another category represented by yellow squares, and uspE is represented by purple; (b) the role of six E. coli USPs in oxidative stress defense, iron metabolism, and cell surface properties; (c) classification of universal stress proteins of E. coli.
Figure 1.
E. coli genes and functions. (a) Location of USP family genes on E. coli chromosomes. uspA, uspC and uspD belong to a category represented by green squares, uspF and uspG belong to another category represented by yellow squares, and uspE is represented by purple; (b) the role of six E. coli USPs in oxidative stress defense, iron metabolism, and cell surface properties; (c) classification of universal stress proteins of E. coli.

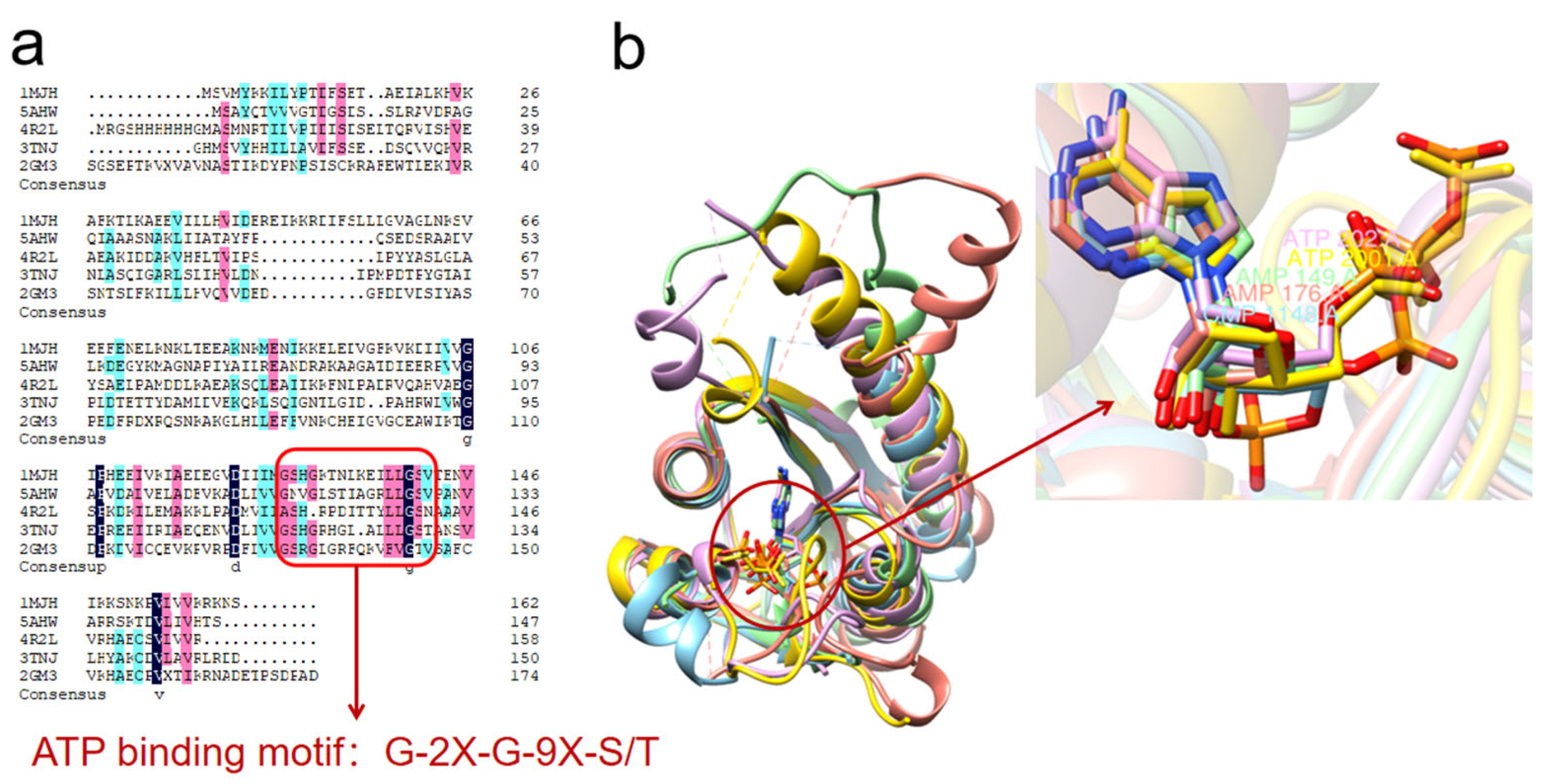
Figure 2.
USPs binding analysis. Crystalline small molecules containing ATP or ATP analogs (AMP, ANP, etc.) were selected for sequence comparison and structure superimposition. (a) Sequence comparison. All except 4R2L contain ATP-binding motifs: G-2X-G-9X-S/T. (b) Superposition. 1MJH, 5AHW, 4R2L, 3TNJ and 2GM3 are gold, blue, purple, green and red, respectively. Superposition of 1MJH as the reference structure revealed that 4R2L still binds ATP at the same position even without the typical ATP binding pattern.
Figure 2.
USPs binding analysis. Crystalline small molecules containing ATP or ATP analogs (AMP, ANP, etc.) were selected for sequence comparison and structure superimposition. (a) Sequence comparison. All except 4R2L contain ATP-binding motifs: G-2X-G-9X-S/T. (b) Superposition. 1MJH, 5AHW, 4R2L, 3TNJ and 2GM3 are gold, blue, purple, green and red, respectively. Superposition of 1MJH as the reference structure revealed that 4R2L still binds ATP at the same position even without the typical ATP binding pattern.

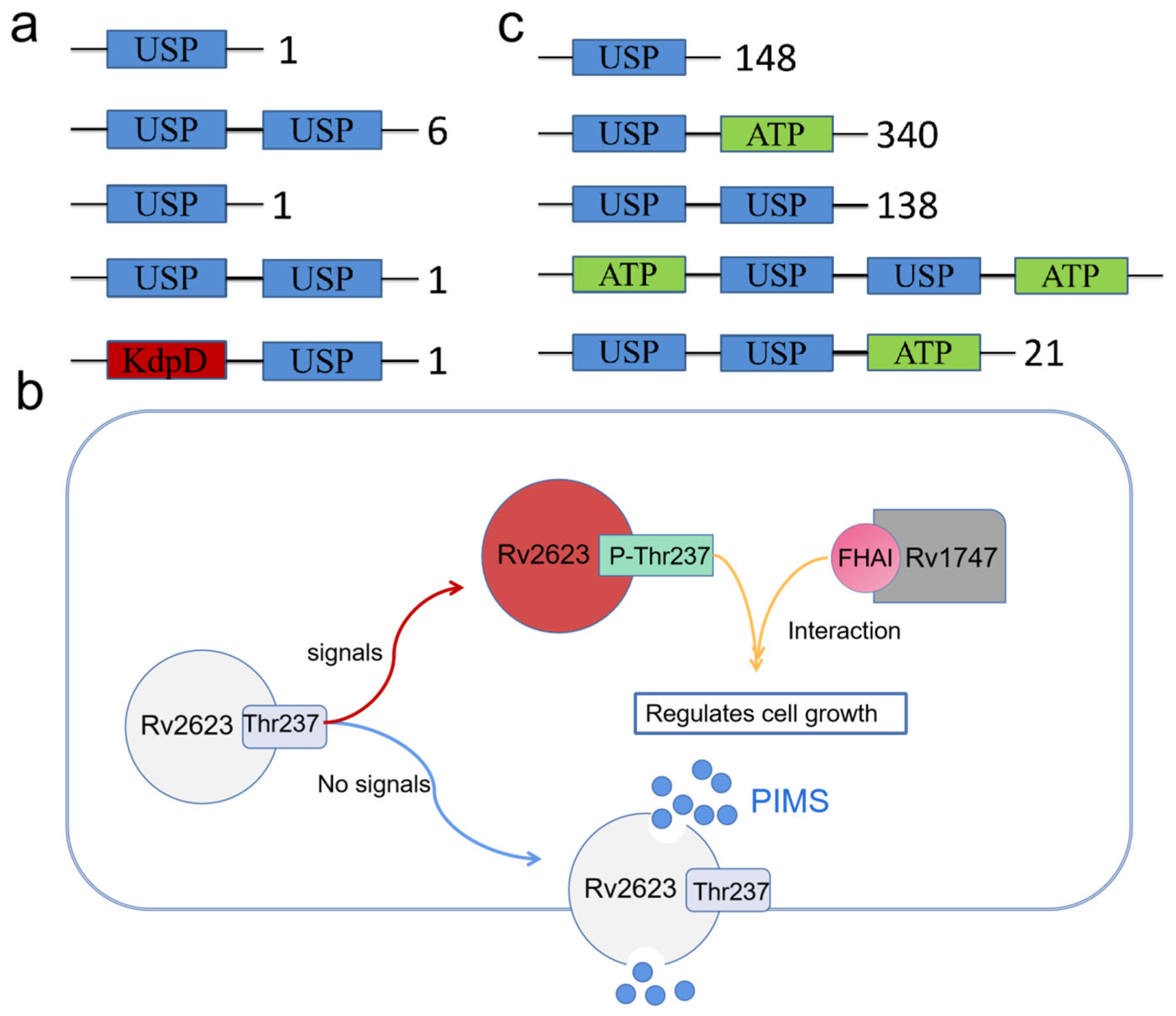
Figure 3.
Classification of M. tuberculosis, Vibrio desulfuricans and interaction of Rv2623 with Rv1747. (a) There are ten USPs of Mycobacterium nucleatum, which are classified into five categories. (b) Rv2623 regulates M. tuberculosis growth. When certain signals are present, Rv2623f Thr237 is phosphorylated and interacts with the FHA structure of Rv1747; when there is no signal from Mycobacterium tuberculosis, this protein transports phosphatidylinositol mannoside (PIM), making M. tuberculosis, possess more virulence. (c) Classification of the USP of Vibrio desulfuricans. The USPs can be classified into five categories according to the number of USPs and bound ATPs, i.e., single USP, one USP bound to one ATP, two USPs alone, two USPs connected to one ATP and two ATPs.
Figure 3.
Classification of M. tuberculosis, Vibrio desulfuricans and interaction of Rv2623 with Rv1747. (a) There are ten USPs of Mycobacterium nucleatum, which are classified into five categories. (b) Rv2623 regulates M. tuberculosis growth. When certain signals are present, Rv2623f Thr237 is phosphorylated and interacts with the FHA structure of Rv1747; when there is no signal from Mycobacterium tuberculosis, this protein transports phosphatidylinositol mannoside (PIM), making M. tuberculosis, possess more virulence. (c) Classification of the USP of Vibrio desulfuricans. The USPs can be classified into five categories according to the number of USPs and bound ATPs, i.e., single USP, one USP bound to one ATP, two USPs alone, two USPs connected to one ATP and two ATPs.

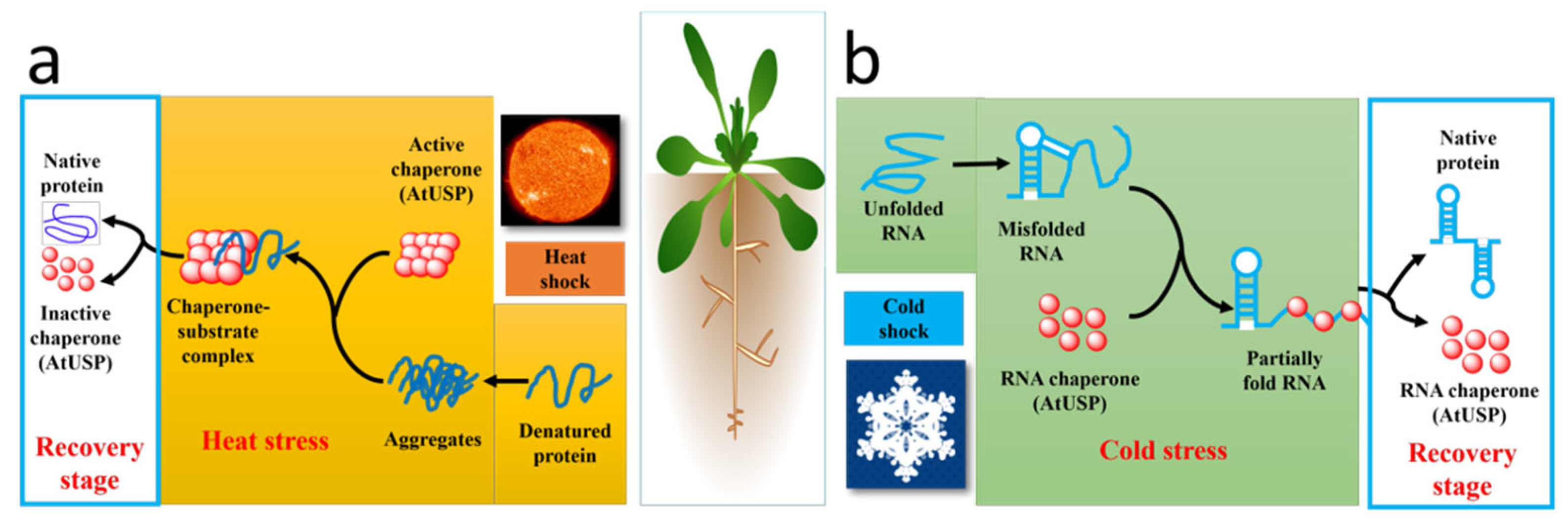
Figure 4.
Role of Arabidopsis USPs in different stresses. (a) When the environment gives Arabidopsis high temperature stress, AtUSP functions as a protein chaperone in the cell, it changes from a low molecular weight complex (LMW) to a high molecular compound (HMW) and binds to proteins that cannot function properly because of high temperature aggregation, helping it to restore its active state after AtUSP reverts back to the LMW state. (b) When subjected to cold stress, AtUSP binds as an RNA chaperone to RNA fragments misfolded by low temperature and helps them to open the misfold and restore the correct fold.
Figure 4.
Role of Arabidopsis USPs in different stresses. (a) When the environment gives Arabidopsis high temperature stress, AtUSP functions as a protein chaperone in the cell, it changes from a low molecular weight complex (LMW) to a high molecular compound (HMW) and binds to proteins that cannot function properly because of high temperature aggregation, helping it to restore its active state after AtUSP reverts back to the LMW state. (b) When subjected to cold stress, AtUSP binds as an RNA chaperone to RNA fragments misfolded by low temperature and helps them to open the misfold and restore the correct fold.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of AtUSP genes in Arabidopsis thaliana along with their corresponding proteins, locus ID, CDS length, protein length, domain and Pfam ID. (Adapted with permission from Ref. [35]. 2023, copyright Springer Nature. and NCBI: https://www.ncbi.nlm.nih.gov/genome/ (accessed on 29 January 2023).
Table 1.
List of AtUSP genes in Arabidopsis thaliana along with their corresponding proteins, locus ID, CDS length, protein length, domain and Pfam ID. (Adapted with permission from Ref. [35]. 2023, copyright Springer Nature. and NCBI: https://www.ncbi.nlm.nih.gov/genome/ (accessed on 29 January 2023).
| Gene | Protein | Locus | CDS Length (bp) | Protein Length (aa) | Domain | Pfam ID |
|---|---|---|---|---|---|---|
| AtUSP1 | AtUSP1 | At1G01680 | 927 | 308 | USP | PF00582 |
| AtUSP2 | AtUSP2 | At1G09740 | 516 | 171 | USP | PF00582 |
| AtUSP3 | AtUSP3 | At1G11360 | 729 | 242 | USP | PF00582 |
| AtUtyK1 | AtUtyK1 | At1G16760 | 2277 | 758 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUtyK2 | AtUtyK2 | At1G17540 | 2187 | 728 | USP | PF00582 |
| Pkinase-Tyr | PF00069 | |||||
| AtUK1 | AtUK1 | At1G21590 | 2271 | 756 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP4 | AtUSP4 | At1G44760 | 642 | 213 | USP | PF00582 |
| AtUSP5 | AtUSP5 | At1G48960 | 660 | 219 | USP | PF00582 |
| AtUSP6 | AtUSP6 | At1G68300 | 483 | 160 | USP | PF00582 |
| AtUSP7 | AtUSP7.1 | At1G69080.1 | 672 | 223 | USP | PF00582 |
| AtUSP7.2 | At1G69080.2 | 630 | 209 | USP | PF00582 | |
| AtUK2 | AtUK2 | At1G77280 | 2385 | 753 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUtyK3 | AtUtyK3 | At1G78940.2 | 2265 | 754 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUSP8 | AtUSP8 | At2G03720 | 498 | 165 | USP | PF00582 |
| AtUK3 | AtUK3 | At2G07020 | 2103 | 700 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP9 | AtUSP9.1 | At2G21620.1 | 564 | 187 | USP | PF00582 |
| AtUSP9.2 | At2G21620.2 | 582 | 193 | USP | PF00582 | |
| AtUK4 | AtUK4 | At2G24370 | 2367 | 788 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP10 | AtUSP10 | At2G47710 | 489 | 162 | USP | PF00582 |
| AtUSP11 | AtUSP11 | At3G01520 | 528 | 175 | USP | PF00582 |
| AtUSP12 | AtUSP12 | At3G03270.1 | 606 | 201 | USP | PF00582 |
| AtUSP13 | AtUSP13 | At3G03290 | 825 | 274 | USP | PF00582 |
| AtUSPUSP1 | AtUSPUSP1.1 | At3G11930.1 | 600 | 199 | USP | PF00582 |
| AtUSPUSP1.2 | At3G11930.2 | 603 | 200 | USP | PF00582 | |
| AtUSPUSP1.3 | At3G11930.3 | 681 | 226 | USP, USP | PF00582 | |
| AtUSPUSP1.4 | At3G11930.4 | 606 | 201 | USP | PF00582 | |
| AtUSP14 | AtUSP14 | At3G17020 | 492 | 163 | USP | PF00582 |
| AtUtyK4 | AtUtyK4 | At3G20200 | 2343 | 780 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP15 | AtUSP15 | At3G21210 | 2415 | 804 | USP | PF00582 |
| AtUSP16 | AtUSP16 | At3G25930 | 465 | 154 | USP | PF00582 |
| AtUSP17 | AtUSP17.1 | At3G53990.1 | 483 | 160 | USP | PF00582 |
| AtUSP17.2 | At3G53990.2 | 381 | 126 | USP | PF00582 | |
| AtUSP18 | AtUSP18.1 | At3G58450.1 | 615 | 204 | USP | PF00582 |
| AtUSP18.2 | At3G58450.2 | 594 | 197 | USP | PF00582 | |
| AtUSP19 | AtUSP19 | At3G62550 | 489 | 162 | USP | PF00582 |
| AtUtyK5 | AtUtyK5 | At4G25160 | 2508 | 835 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUSP20 | AtUSP20 | AT4G13450.1 | 660 | 219 | USP | PF00582 |
| AtUSP21 | AtUSP21 | At4G27320 | 783 | 260 | USP | PF00582 |
| AtUK5 | AtUK5 | At4G31230 | 2295 | 764 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUtyK6 | AtUtyK6 | At5G12000 | 2106 | 701 | USP | PF00582 |
| Pkinase-Tyr | PF07714 | |||||
| AtUSP22 | AtUSP22 | At5G14680 | 528 | 175 | USP | PF00582 |
| AtUSP23 | AtUSP23 | At5G17390 | 858 | 285 | USP | PF00582 |
| AtUSP24 | AtUSP24 | At5G20310 | 1185 | 394 | USP | PF00582 |
| AtUK6 | AtUK6 | At5G35380 | 2196 | 731 | USP | PF00582 |
| Pkinase | PF00069 | |||||
| AtUSP25 | AtUSP25.1 | At5G47740.1 | 735 | 244 | USP | PF00582 |
| AtUSP25.2 | At5G47740.2 | 741 | 246 | USP | PF00582 | |
| AtUSP26 | AtUSP26 | At5G49050 | 453 | 150 | USP | PF00582 |
| AtUSP27 | AtUSP27 | At5G54430 | 729 | 242 | USP | PF00582 |
| AtUK7 | AtUK7 | At5G63940 | 2118 | 705 | USP | PF00582 |
| Pkinase | PF00069 |
Table 2.
Summary of crystal structures of USP family proteins.
| Organism | PDB Code | Chains | Mutation(s) | Small Molecules | References |
|---|---|---|---|---|---|
| Methanocaldococcus jannaschii | 1MJH | 2 | 0 | ATP, Mn2+ | [57] |
| Haemophilus influenzae | 1JMV | 4 | 0 | SO42− | [58] |
| Methanocaldococcus jannaschii | 5AHW | 6 | 0 | CMP, SO42−, POG, Cl− | [59] |
| Salmonella enterica subsp. enterica serovar Typhimurium str. LT2 | 4R2J | 1 | 0 | GLC, PO43−, EDO, Zn2+ | [60] |
| 4R2K | 1 | 1 (4R2J Mutation) | SO42−, EDO, OXD | ||
| 4R2L | 2 | 0 | EDO, Cl−, ATP, Mg2+ | ||
| 4R2M | 2 | 1 (4R2L Mutation) | ANP, Mg2+ | ||
| Nitrosomonas europaea ATCC 19718 | 3TNJ | 1 | 0 | AMP | [61] |
| 2PFS | 1 | 3 | MSE, Cl− | ||
| Archaeoglobus fulgidus | 3QTB | 2 | 0 | D5M, ACT, MSE | |
| 6HCD | 4 | 0 | ACT, MSE, Cl− | ||
| Escherichia coli K-12 | 5CB0 | 2 | 0 | Z6X | [63] |
| Arabidopsis thaliana | 2GM3 | 6 | 3 | AMP, MSE | [64] |
Table 3.
Summary of the functions of USPs in different plants.
| Plant Species | Name of USPs | Functions | References |
|---|---|---|---|
| Arabidopsis thaliana | AtUSP | Protein chaperone | [85] |
| RNA chaperone | [90] | ||
| Inherently antifungal activity | [55] | ||
| AT5G35380 | Related to resistance to flooding | [86] | |
| GRUSP | Promote seed germination | [36] | |
| Novel regulatory components of the flowering signal transduction pathway | [40] | ||
| HRU1 | Coordination of oxygen sensing and ROS signaling under hypoxic conditions | [87] | |
| AtUSP17 | Related to the salt tolerance of the plant | [42] | |
| Gossypium arboretum | GUSP1 | Activates downstream genes in response to drought | [49,50] |
| GUSP2 | Directly involved in stress tolerance or as signaling molecules to activate stress adaptation mechanisms | [51] | |
| Malus sieversii | MsUSPA | Reducing transpiration and retaining water by altering the cellular structure of the leaves to improve drought resistance | [46] |
| Increase the activity of antioxidant enzymes, reduce the accumulation of Ros and improve the antioxidant capacity | |||
| Solanum lycoperiscus | SlRd2 | Regulation of SlCipk6-mediated ROS generation | [102] |
| Solanum pennellii | SpUSP | Improving tolerance to oxidative stress | [44] |
| Salicornia brachiata | SbUSP | Participate in tolerance mechanisms or act as molecular switches (signaling molecules) to activate stress adaptation mechanisms | [53] |
| Sulfolobus acidocaldarius | SaUspA | Binds phosphatase and alters phosphatase activity | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Luo, D.; Wu, Z.; Bai, Q.; Zhang, Y.; Huang, M.; Huang, Y.; Li, X. Universal Stress Proteins: From Gene to Function. Int. J. Mol. Sci. 2023, 24, 4725. https://doi.org/10.3390/ijms24054725
AMA Style
Luo D, Wu Z, Bai Q, Zhang Y, Huang M, Huang Y, Li X. Universal Stress Proteins: From Gene to Function. International Journal of Molecular Sciences. 2023; 24(5):4725. https://doi.org/10.3390/ijms24054725
Chicago/Turabian StyleLuo, Dan, Zilin Wu, Qian Bai, Yong Zhang, Min Huang, Yajiao Huang, and Xiangyang Li. 2023. "Universal Stress Proteins: From Gene to Function" International Journal of Molecular Sciences 24, no. 5: 4725. https://doi.org/10.3390/ijms24054725
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.