
Osteocyte-Derived CaMKK2 Regulates Osteoclasts and Bone Mass in a Sex-Dependent Manner through Secreted Calpastatin
 , ,
, ,
Abstract
:1. Introduction
2. Results
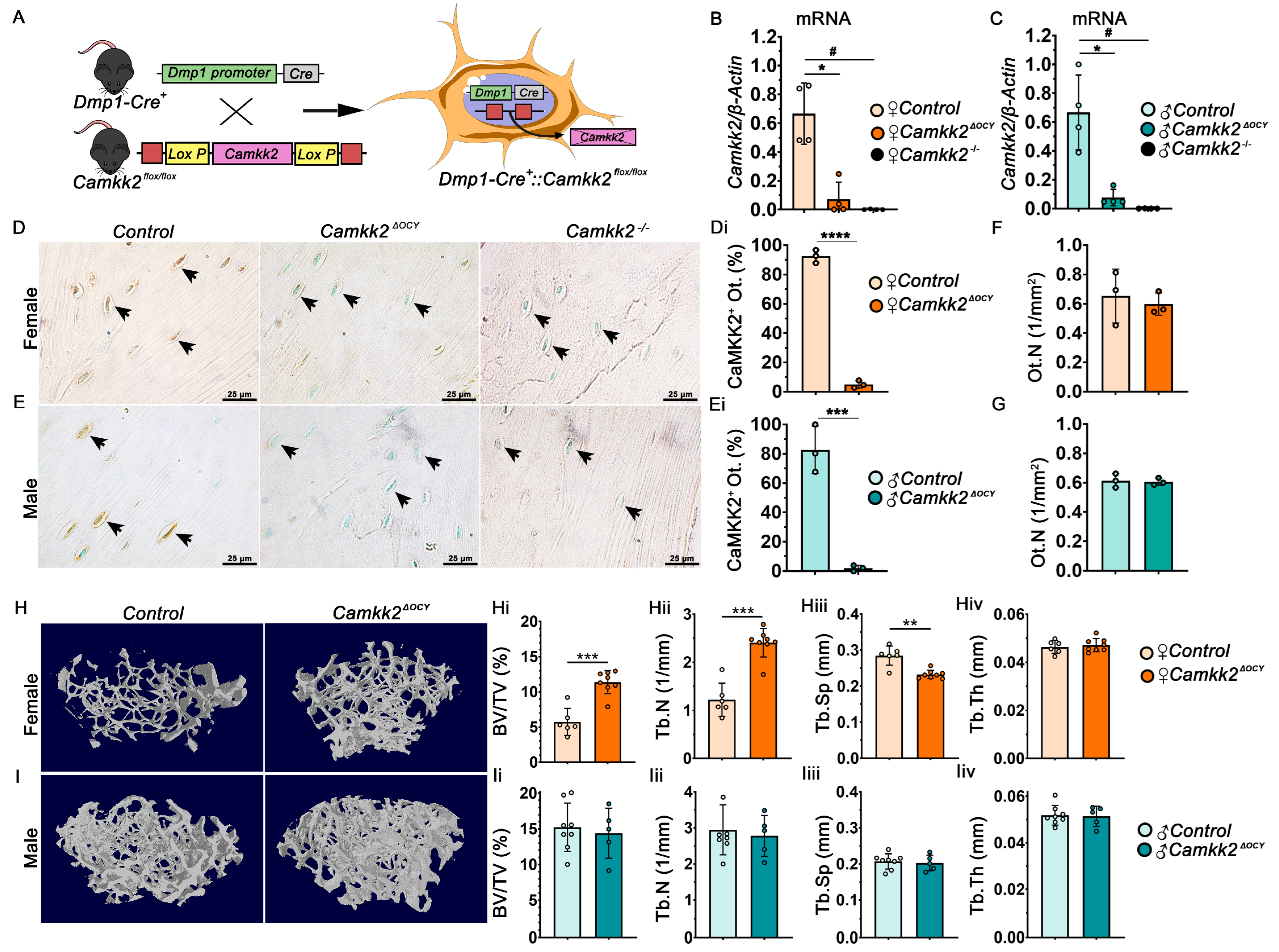
2.1. Conditional Deletion of CaMKK2 in Osteocytes Enhances Trabecular Bone Mass Only in Female Mice
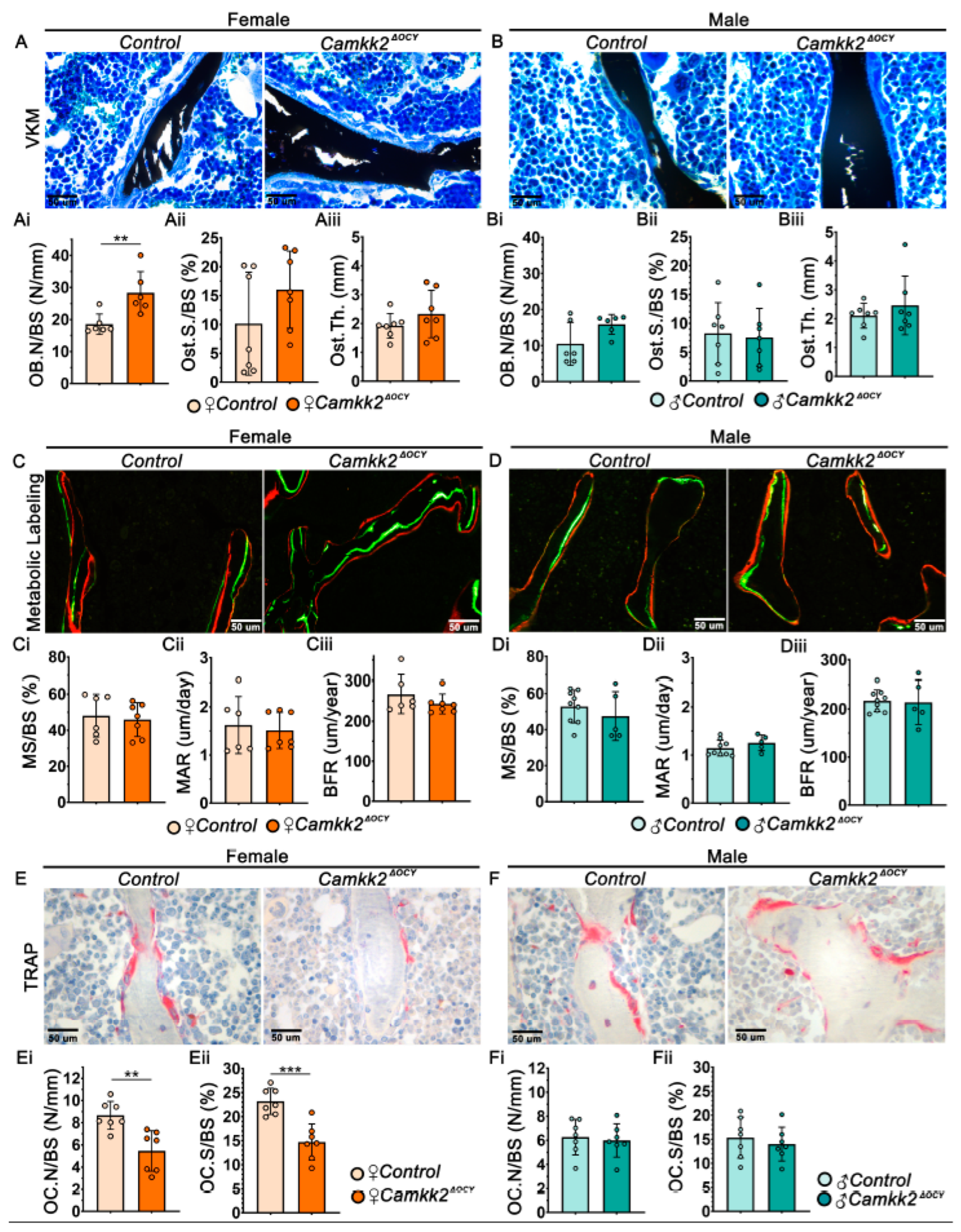
2.2. Ablation of Osteocyte-Derived CaMKK2 Diminishes OCs in Female Mice without Altering OB Function
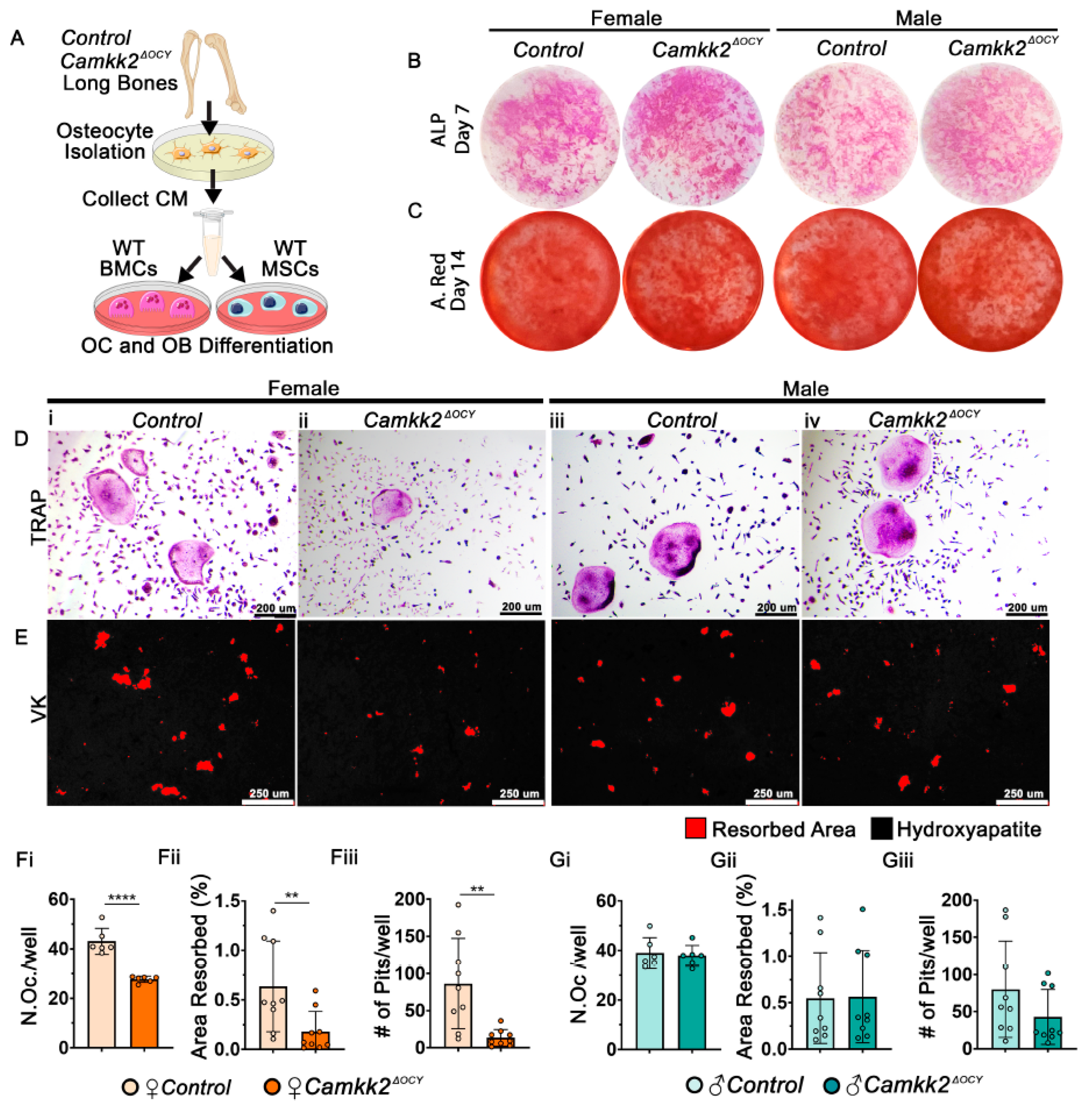
2.3. Conditioned Media from Female Osteocytes Lacking CaMKK2 Inhibits OC Differentiation and Function
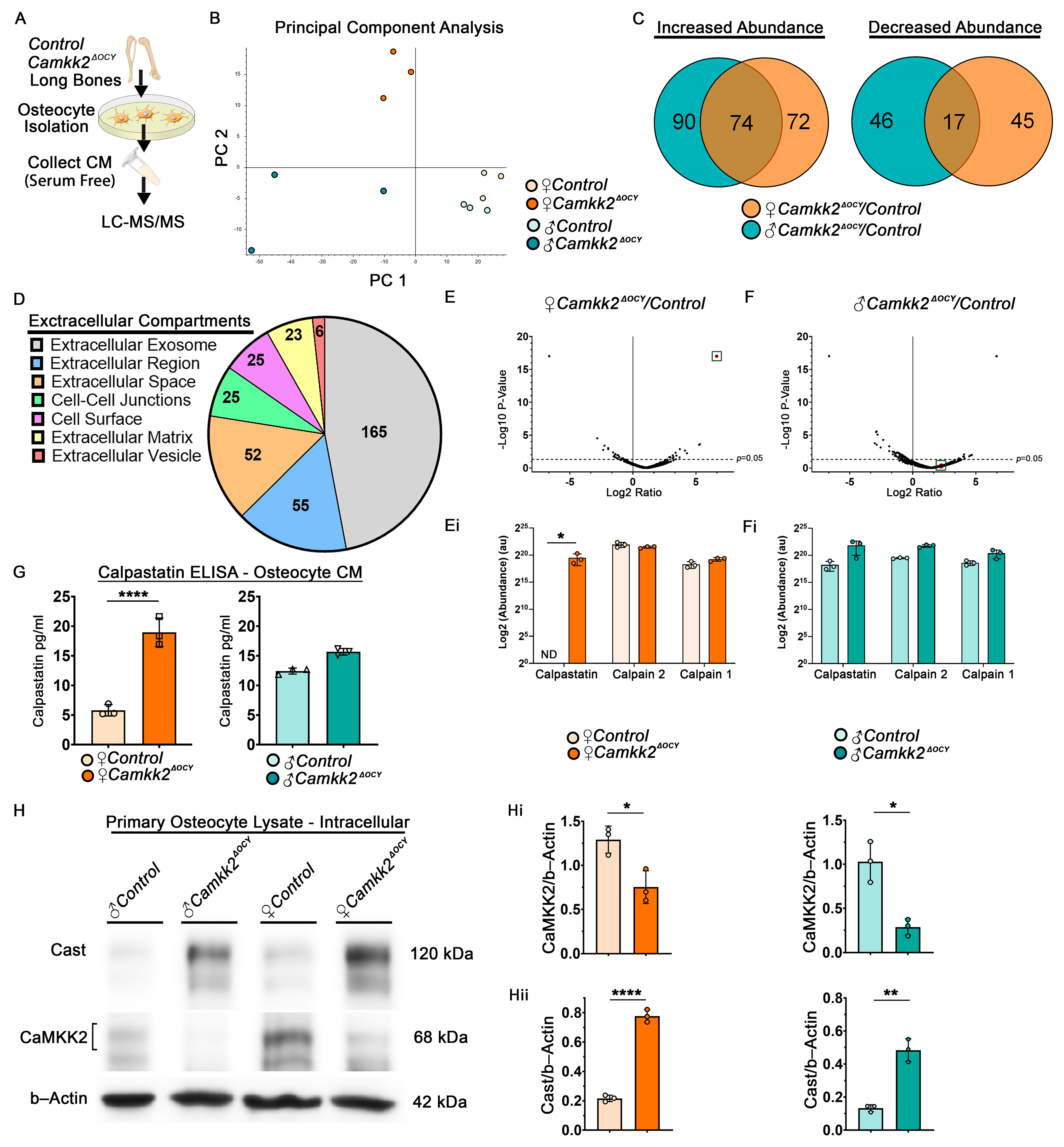
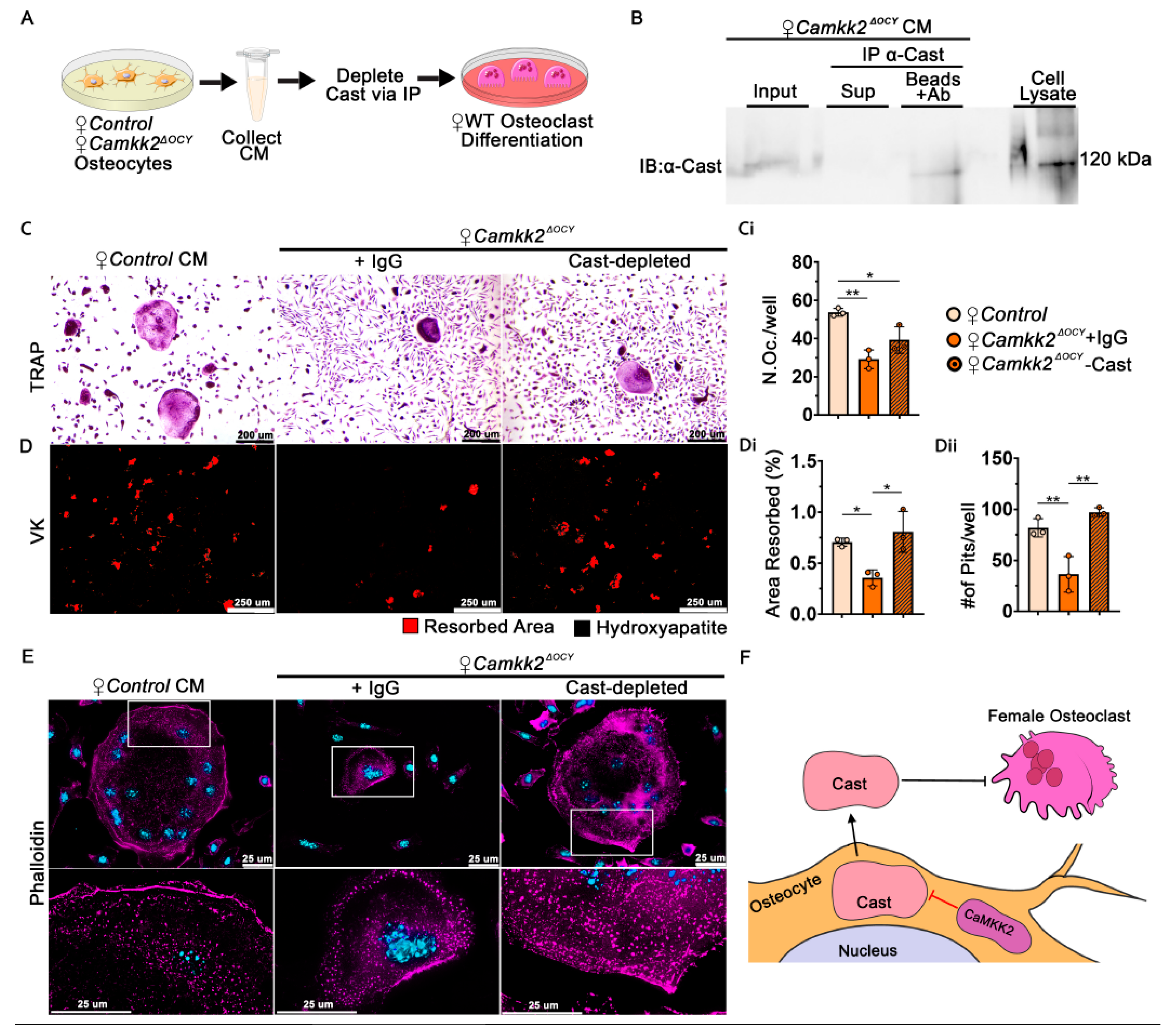
2.4. Extracellular Calpastatin Is Enriched in Female CaMKK2-Deficient Osteocyte Conditioned Media Compared to Control and Its Sequestration Relieves OC Inhibition
2.5. Osteoclast Inhibitory Effects of Recombinant Non-Cell Permeable Calpastatin Is Sex-Dependent
2.6. Extracellular Calpastatin in Female Camkk2ΔOCY CM Regulates Bone Resorption by Osteoclasts
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. µCT Imaging
4.3. Histology
4.4. Immunohistochemistry
4.5. Osteocyte Culture and Media Collection
4.6. Osteoclast Cultures and Media Supplementation:
4.7. qRT-PCR
4.8. Mass Spectrometry
4.9. Proteomics Data Analysis
4.10. Calpastatin Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Immunoblotting
4.12. Immunoprecipitation of Calpastatin from Conditioned Media
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, M.R.; Burr, D.B. Chapter 4—Bone Modeling and Remodeling. In Basic and Applied Bone Biology; Burr, D.B., Allen, M.R., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 75–90. [Google Scholar]
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef]
- Bellido, T.; Plotkin, L.I.; Bruzzaniti, A. Chapter 2—Bone Cells. In Basic and Applied Bone Biology; Burr, D.B., Allen, M.R., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 27–45. [Google Scholar]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell… and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [Green Version]
- Robling, A.G.; Bonewald, L.F. The Osteocyte: New Insights. Annu. Rev. Physiol. 2020, 82, 485–506. [Google Scholar] [CrossRef] [Green Version]
- Uda, Y.; Azab, E.; Sun, N.; Shi, C.; Pajevic, P.D. Osteocyte Mechanobiology. Curr. Osteoporos. Rep. 2017, 15, 318–325. [Google Scholar] [CrossRef]
- Anderson, K.A.; Means, R.L.; Huang, Q.H.; Kemp, B.E.; Goldstein, E.G.; Selbert, M.A.; Edelman, A.M.; Fremeau, R.T.; Means, A.R. Components of a calmodulin-dependent protein kinase cascade. Molecular cloning, functional characterization and cellular localization of Ca2+/calmodulin-dependent protein kinase kinase beta. J. Biol. Chem. 1998, 273, 31880–31889. [Google Scholar] [CrossRef] [Green Version]
- Green, M.F.; Anderson, K.A.; Means, A.R. Characterization of the CaMKKbeta-AMPK signaling complex. Cell. Signal. 2011, 23, 2005–2012. [Google Scholar] [CrossRef] [Green Version]
- Hurley, R.L.; Anderson, K.A.; Franzone, J.M.; Kemp, B.E.; Means, A.R.; Witters, L.A. The Ca2+/calmodulin-dependent protein kinase kinases are AMP-activated protein kinase kinases. J. Biol. Chem. 2005, 280, 29060–29066. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.A.; Lin, F.; Ribar, T.J.; Stevens, R.D.; Muehlbauer, M.J.; Newgard, C.B.; Means, A.R. Deletion of CaMKK2 from the liver lowers blood glucose and improves whole-body glucose tolerance in the mouse. Mol. Endocrinol. 2012, 26, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.A.; Ribar, T.J.; Lin, F.; Noeldner, P.K.; Green, M.F.; Muehlbauer, M.J.; Witters, L.A.; Kemp, B.E.; Means, A.R. Hypothalamic CaMKK2 Contributes to the Regulation of Energy Balance. Cell Metab. 2008, 7, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Mével, E.; Shutter, J.A.; Ding, X.; Mattingly, B.T.; Williams, J.N.; Li, Y.; Huls, A.; Kambrath, A.V.; Trippel, S.B.; Wagner, D.; et al. Systemic inhibition or global deletion of CaMKK2 protects against post-traumatic osteoarthritis. Osteoarthr. Cartil. 2021, 30, 124–136. [Google Scholar] [CrossRef]
- Racioppi, L.; Noeldner, P.K.; Lin, F.; Arvai, S.; Means, A.R. Calcium/calmodulin-dependent protein kinase kinase 2 regulates macrophage-mediated inflammatory responses. J. Biol. Chem. 2012, 287, 11579–11591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cary, R.L.; Waddell, S.; Racioppi, L.; Long, F.; Novack, D.V.; Voor, M.J.; Sankar, U. Inhibition of Ca2+/calmodulin-dependent protein kinase kinase 2 stimulates osteoblast formation and inhibits osteoclast differentiation. J. Bone Miner. Res. 2013, 28, 1599–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, Z.J.; Cary, R.L.; Yang, C.; Novack, D.V.; Voor, M.J.; Sankar, U. Inhibition of CaMKK2 reverses age-associated decline in bone mass. Bone 2015, 75, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Tokumitsu, H.; Inuzuka, H.; Ishikawa, Y.; Ikeda, M.; Saji, I.; Kobayashi, R. STO-609, a specific inhibitor of the Ca(2+)/calmodulin-dependent protein kinase kinase. J. Biol. Chem. 2002, 277, 15813–15818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.N.; Kambrath, A.V.; Patel, R.B.; Kang, K.S.; Mevel, E.; Li, Y.; Cheng, Y.; Pucylowski, A.J.; Hassert, M.A.; Voor, M.J.; et al. Inhibition of CaMKK2 Enhances Fracture Healing by Stimulating Indian Hedgehog Signaling and Accelerating Endochondral Ossification. J. Bone Miner. Res. 2018, 33, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Bivi, N.; Pacheco-Costa, R.; Brun, L.R.; Murphy, T.R.; Farlow, N.R.; Robling, A.G.; Bellido, T.; Plotkin, L.I. Absence of Cx43 selectively from osteocytes enhances responsiveness to mechanical force in mice. J. Orthop. Res. 2013, 31, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Bivi, N.; Condon, K.W.; Allen, M.R.; Farlow, N.; Passeri, G.; Brun, L.R.; Rhee, Y.; Bellido, T.; I Plotkin, L. Cell autonomous requirement of connexin 43 for osteocyte survival: Consequences for endocortical resorption and periosteal bone formation. J. Bone Miner. Res. 2012, 27, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Calle, J.; Bellido, T. The osteocyte as a signaling cell. Physiol. Rev. 2022, 102, 379–410. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef]
- Goll, D.E.; Thompson, V.F.; Li, H.; Wei, W.; Cong, J. The Calpain System. Physiol. Rev. 2003, 83, 731–801. [Google Scholar] [CrossRef]
- Sharma, R.K.; Parameswaran, S. Calmodulin-binding proteins: A journey of 40 years. Cell Calcium 2018, 75, 89–100. [Google Scholar] [CrossRef]
- Hayashi, M.; Koshihara, Y.; Ishibashi, H.; Yamamoto, S.; Tsubuki, S.; Saido, T.C.; Kawashima, S.; Inomata, M. Involvement of Calpain in Osteoclastic Bone Resorption. J. Biochem. 2005, 137, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.Y.; Kim, D.W.; Karmin, J.A.; Hong, D.; Chang, S.S.; Fujisawa, M.; Takayanagi, H.; Bigliani, L.U.; Blaine, T.A.; Lee, H.-J. mu-Calpain regulates receptor activator of NF-kappaB ligand (RANKL)-supported osteoclastogenesis via NF-kappaB activation in RAW 264.7 cells. J. Biol. Chem. 2005, 280, 29929–29936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peltier, J.; Bellocq, A.; Perez, J.; Doublier, S.; Dubois, Y.-C.X.; Haymann, J.-P.; Camussi, G.; Baud, L. Calpain Activation and Secretion Promote Glomerular Injury in Experimental Glomerulonephritis: Evidence from Calpastatin-Transgenic Mice. J. Am. Soc. Nephrol. 2006, 17, 3415–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, R.V.; Goust, J.-M.; Chakrabarti, A.K.; Barbosa, E.; Hogan, E.L.; Banik, N.L. Calpain Expression in Lymphoid Cells: INCREASED mRNA AND PROTEIN LEVELS AFTER CELL ACTIVATION (∗). J. Biol. Chem. 1995, 270, 2497–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, F.; Letavernier, E.; Abid, S.; Houssaini, A.; Czibik, G.; Marcos, E.; Rideau, D.; Parpaleix, A.; Lipskaia, L.; Amsellem, V.; et al. Extracellular Calpain/Calpastatin Balance Is Involved in the Progression of Pulmonary Hypertension. Am. J. Respir. Cell Mol. Biol. 2016, 55, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Frangié, C.; Zhang, W.; Perez, J.; Dubois, Y.-C.X.; Haymann, J.-P.; Baud, L. Extracellular Calpains Increase Tubular Epithelial Cell Mobility: IMPLICATIONS FOR KIDNEY REPAIR AFTER ISCHEMIA*. J. Biol. Chem. 2006, 281, 26624–26632. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Su, Z.; Liu, F. Effects of functionally diverse calpain system on immune cells. Immunol. Res. 2021, 69, 8–17. [Google Scholar] [CrossRef]
- Ono, Y.; Sorimachi, H. Calpains—An elaborate proteolytic system. Biochim. Et Biophys. Acta (BBA)—Proteins Proteom. 2012, 1824, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Marzia, M.; Chiusaroli, R.; Neff, L.; Kim, N.-Y.; Chishti, A.H.; Baron, R.; Horne, W.C. Calpain is required for normal osteoclast function and is down-regulated by calcitonin. J. Biol. Chem. 2006, 281, 9745–9754. [Google Scholar] [CrossRef] [Green Version]
- Murray, S.S.; Grisanti, M.S.; Bentley, G.V.; Kahn, A.J.; Urist, M.R.; Murray, E.J.B. The Calpain–Calpastatin System and Cellular Proliferation and Differentiation in Rodent Osteoblastic Cells. Exp. Cell Res. 1997, 233, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Coury, F.; Zenger, S.; Stewart, A.K.; Stephens, S.; Neff, L.; Tsang, K.; Shull, G.E.; Alper, S.L.; Baron, R.; Aliprantis, A.O. SLC4A2-mediated Cl−/HCO3− exchange activity is essential for calpain-dependent regulation of the actin cytoskeleton in osteoclasts. Proc. Natl. Acad. Sci. USA 2013, 110, 2163–2168. [Google Scholar] [PubMed] [Green Version]
- Yamamoto, S.; Shimizu, K.; Shimizu, K.; Suzuki, K.; Nakagawa, Y.; Yamamuro, T. Calcium-dependent cysteine proteinase (calpain) in human arthritic synovial joints. Arthritis Rheum. 1992, 35, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Shimizu, K.; Hamamoto, T.; Suzuki, K.; Ueda, M.; Yamamuro, T. Calcium-dependent neutral proteinase (calpain) in fracture healing in rats. J. Orthop. Res. 1994, 12, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Hamamoto, T.; Hamakubo, T.; Lee, W.J.; Suzuki, K.; Nakagawa, Y.; Murachi, T.; Yamamuro, T. Immunohistochemical and biochemical demonstration of calcium-dependent cysteine proteinase (calpain) in calcifying cartilage of rats. J. Orthop. Res. 1991, 9, 26–36. [Google Scholar] [CrossRef]
- Adachi, E.; Mukaiyama, T.; Sasai, K.; Hayashi, T.; Kawashima, S.; Kasai, Y.; Hayashi, M.; Hashimoto, P.M. Immunohistochemical Evidence of the Extracellular Localization of Calcium-Activated Neutral Protease (CANP) in Rabbit Skeletal Muscle, Lung and Aorta. Arch. Histol. Cytol. 1990, 53, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Badalamente, M.A.; Hurst, L.C.; Stracher, A. Localization and inhibition of calcium-activated neutral protease (CANP) in primate skeletal muscle and peripheral nerve. Exp. Neurol. 1987, 98, 357–369. [Google Scholar] [CrossRef]
- Wang, Q.; Da’dara, A.A.; Skelly, P.J. The human blood parasite Schistosoma mansoni expresses extracellular tegumental calpains that cleave the blood clotting protein fibronectin. Sci. Rep. 2017, 7, 12912. [Google Scholar] [CrossRef] [Green Version]
- Nishihara, H.; Nakagawa, Y.; Ishikawa, H.; Ohba, M.; Shimizu, K.; Nakamura, T. Matrix Vesicles and Media Vesicles as Nonclassical Pathways for the Secretion of m-Calpain from MC3T3-E1 Cells. Biochem. Biophys. Res. Commun. 2001, 285, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Burns, G.; Brooks, K.; Wildung, M.; Navakanitworakul, R.; Christenson, L.K.; Spencer, T.E. Extracellular vesicles in luminal fluid of the ovine uterus. PLoS ONE 2014, 9, e90913. [Google Scholar] [CrossRef] [Green Version]
- Fukui, I.; Tanaka, K.; Murachi, T. Extracellular appearance of calpain and calpastatin in the synovial fluid of the knee joint. Biochem. Biophys. Res. Commun. 1989, 162, 559–566. [Google Scholar] [CrossRef]
- Storr, S.J.; Carragher, N.O.; Frame, M.C.; Parr, T.; Martin, S.G. The calpain system and cancer. Nat. Rev. Cancer 2011, 11, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Nian, H.; Ma, B. Calpain–calpastatin system and cancer progression. Biol. Rev. 2021, 96, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ciani, C.; Doty, S.B.; Fritton, S.P. Delineating bone’s interstitial fluid pathway in vivo. Bone 2004, 34, 499–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knothe Tate, M.L.; Niederer, P.; Knothe, U. In Vivo Tracer Transport Through the Lacunocanalicular System of Rat Bone in an Environment Devoid of Mechanical Loading. Bone 1998, 22, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Letavernier, B.; Zafrani, L.; Nassar, D.; Perez, J.; Levi, C.; Bellocq, A.; Mesnard, L.; Sachon, E.; Haymann, J.-P.; Aractingi, S.; et al. Calpains Contribute to Vascular Repair in Rapidly Progressive Form of Glomerulonephritis: Potential Role of Their Externalization. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Müller, R. Guidelines for assessment of bone microstructure in rodents using micro–computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Stern, A.R.; Stern, M.M.; Van Dyke, M.E.; Jahn, K.; Prideaux, M.; Bonewald, L.F. Isolation and culture of primary osteocytes from the long bones of skeletally mature and aged mice. Biotechniques 2012, 52, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Bland, J.M.; Altman, D.G. Multiple significance tests: The Bonferroni method. BMJ 1995, 310, 170. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Increased Protein Abundance (Camkk2ΔOCY/Control) | Female | Male | |||
| Biological Process | Reactome Pathway | Fold Enrichment | p-Value | Fold Enrichment | p-Value |
| Cell Cycle | Golgi Cisternae Pericentriolar Stack Reorganization | 6.7 | 0.082 | 10.5 | 0.092 |
| CDK-mediated phosphorylation and removal of Cdc6 | 6.7 | 0.021 | 10.5 | 0.000046 | |
| SCF(Skp2)-mediated degradation of p27/p21 | N/A | N/A | 10.3 | 0.00001 | |
| Autodegradation of the E3 ubiquitin ligase COP1 | 6.6 | 0.022 | 10.3 | 0.000051 | |
| Ubiquitin Mediated Degradation of Phosphorylated Cdc25A | 6.5 | 0.023 | 10.1 | 0.000056 | |
| AUF1 binds and destabilizes mRNA | 6.3 | 0.025 | 9.8 | 0.000069 | |
| Cdc20:Phospho-APC/C mediated degradation of Cyclin A | 6.1 | 0.0087 | N/A | N/A | |
| Extracellular Matrix Organization | Degradation of the extracellular matrix | 6 | 0.028 | N/A | N/A |
| Hemostasis | Cell surface interactions at the vascular wall | 7.4 | 0.016 | N/A | N/A |
| Immune System | Cross-presentation of soluble endosomes | 7.4 | 0.016 | 11.5 | 0.000026 |
| Activation of NF-kappaB | N/A | N/A | 9.4 | 0.000019 | |
| Metabolism | Regulation of ornithine decarboxylase (ODC) | 8.6 | 0.0026 | 10.7 | 0.000041 |
| Signal Transduction | Degradation of GLI1 by the proteasome | 6.1 | 0.027 | 10.8 | 0.0000074 |
| GLI3 is processed to GLI3R by the proteasome | N/A | N/A | 10.6 | 0.0000082 | |
| Dectin-1 mediated noncanonical NF-kB signaling | N/A | N/A | 10.5 | 0.0000092 | |
| NIK noncanonical NF-kB signaling | N/A | N/A | 10.5 | 0.0000092 | |
| Degradation of beta-catenin by the destruction complex | 6.5 | 0.0069 | 10.4 | 0.0000018 | |
| Degradation of AXIN | 6.3 | 0.025 | 9.8 | 0.000069 | |
| Degradation of DVL | 6.1 | 0.027 | 9.4 | 0.000083 | |
| Hedgehog ligand biogenesis | N/A | N/A | 9.3 | 0.000091 | |
| Vesicle-Mediated Transport | Golgi Associated Vesicle Biogenesis | 7.1 | 0.018 | N/A | N/A |
| COPI-mediated anterograde transport | 6.5 | 0.0069 | N/A | N/A | |
| Decreased Protein Abundance (Camkk2ΔOCY/Control) | Female | Male | |||
| Biological Process | Reactome Pathway | Fold Enrichment | p-Value | Fold Enrichment | p-Value |
| Metabolism of Proteins | Formation of a pool of free 40S subunits | 8.1 | 0.0012 | N/A | N/A |
| Formation of the ternary and 43S complex | 6.5 | 0.0015 | N/A | N/A | |
| L13a-mediated translational silencing of Ceruloplasmin expression | 8.1 | 0.0017 | N/A | N/A | |
| GTP hydrolysis and joining of the 60S ribosomal subunit | 6.5 | 0.0021 | N/A | N/A | |
| Translation initiation complex formation | 6.5 | 0.0022 | N/A | N/A | |
| Ribosomal scanning and start codon recognition | 6.5 | 0.009 | N/A | N/A | |
| SRP-dependent co-translational protein targeting to membrane | 6.5 | 0.0093 | N/A | N/A | |
| Metabolism of RNA | Nonsense Mediated Decay enhanced by the Exon Junction Complex | 6.5 | 0.016 | N/A | N/A |
| Signal Transduction | Signaling by EGFR | N/A | N/A | 77.3 | 0.025 |
| EGFR Transactivation by Gastrin | N/A | N/A | 60.1 | 0.032 | |
| Cell-Cell Communication | Type I hemidesmosome assembly | N/A | N/A | 49.2 | 0.039 |
| Adherens junctions interactions | N/A | N/A | 26.2 | 0.0053 | |
| Extracellular matrix Organization | Collagen degradation | N/A | N/A | 15.9 | 0.0017 |
| Degradation of the extracellular matrix | N/A | N/A | 13.1 | 0.02 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, J.N.; Irwin, M.; Li, Y.; Kambrath, A.V.; Mattingly, B.T.; Patel, S.; Kittaka, M.; Collins, R.N.; Clough, N.A.; Doud, E.H.; et al. Osteocyte-Derived CaMKK2 Regulates Osteoclasts and Bone Mass in a Sex-Dependent Manner through Secreted Calpastatin. Int. J. Mol. Sci. 2023, 24, 4718. https://doi.org/10.3390/ijms24054718
Williams JN, Irwin M, Li Y, Kambrath AV, Mattingly BT, Patel S, Kittaka M, Collins RN, Clough NA, Doud EH, et al. Osteocyte-Derived CaMKK2 Regulates Osteoclasts and Bone Mass in a Sex-Dependent Manner through Secreted Calpastatin. International Journal of Molecular Sciences. 2023; 24(5):4718. https://doi.org/10.3390/ijms24054718
Chicago/Turabian StyleWilliams, Justin N., Mavis Irwin, Yong Li, Anuradha Valiya Kambrath, Brett T. Mattingly, Sheel Patel, Mizuho Kittaka, Rebecca N. Collins, Nicholas A. Clough, Emma H. Doud, and et al. 2023. "Osteocyte-Derived CaMKK2 Regulates Osteoclasts and Bone Mass in a Sex-Dependent Manner through Secreted Calpastatin" International Journal of Molecular Sciences 24, no. 5: 4718. https://doi.org/10.3390/ijms24054718