1. Introduction
A nonsense mutation generates an in-frame premature termination codon (PTC) within the open reading frame of an mRNA. A PTC reduces the amount of functional protein translated from an mRNA in two ways. First, a PTC terminates translation before a full-length protein can be generated, resulting in a truncated polypeptide that lacks normal function or is unstable. Second, a PTC can activate nonsense-mediated mRNA decay (NMD), a conserved eukaryotic cellular pathway that targets PTC-containing mRNAs for degradation, reducing the pool of mRNA available for translation. Significantly, 11% of all gene lesions associated with a genetic disease generate an in-frame PTC [
1], indicating that millions of patients carry this type of mutation.
One way to overcome the effects of nonsense mutations on gene expression is to suppress translation termination at PTCs, allowing partial levels of full-length, functional protein to be restored. PTC suppression occurs when an aminoacyl-tRNA that base-pairs with two of the three nucleotides of a PTC, known as a near-cognate aminoacyl-tRNA, becomes accommodated into the ribosomal acceptor site and its amino acid is added to the nascent polypeptide at the PTC position [
2,
3]. This mechanism, often termed “readthrough”, permits translation elongation to continue downstream of the PTC in the correct ribosomal reading frame, generating a full-length polypeptide. The use of small molecules that promote the readthrough of PTCs is currently being explored as a precision/personalized medicine approach to restore the deficient protein in patients who harbor a nonsense mutation. However, the readthrough agents that are currently available for clinical use are unlikely to be effective enough to treat most genetic diseases. Factors that can limit the effectiveness of current readthrough agents include: the mRNA context surrounding a PTC [
4,
5], the nature of the amino acid that becomes incorporated at a PTC during readthrough [
2,
3], the efficiency of NMD that limits mRNA availability for readthrough [
6,
7,
8], and the level of protein function required to alleviate a disease phenotype. More effective and safe readthrough agents are needed that can overcome these limitations and can be applied to diseases for which rescue of partial levels of protein function is sufficient to normalize the phenotype.
One such disease that could benefit from readthrough therapies is Mucopolysaccharidosis I (MPS I). MPS I is caused by a deficiency of the lysosomal enzyme, α-L-iduronidase (encoded by the
IDUA gene), that participates in the catabolism of the glycosaminoglycans (GAGs) dermatan sulfate and heparan sulfate. α-L-iduronidase insufficiency results in the cumulative storage of these partially digested GAGs in lysosomes. Complete loss of α-L-iduronidase leads to the onset of the severe, Hurler form of MPS I (MPS I-H), which is characterized by progressive development of abnormalities in the cartilage, bone, heart, spleen, liver, lung, and neurological tissues, ultimately resulting in a reduced lifespan. MPS I-H represents an excellent candidate for readthrough therapies since the majority of MPS I-H patients carry a nonsense mutation [
9] and even low levels of normal α-L-iduronidase activity (<1%) can significantly attenuate MPS I-H phenotypes [
10].
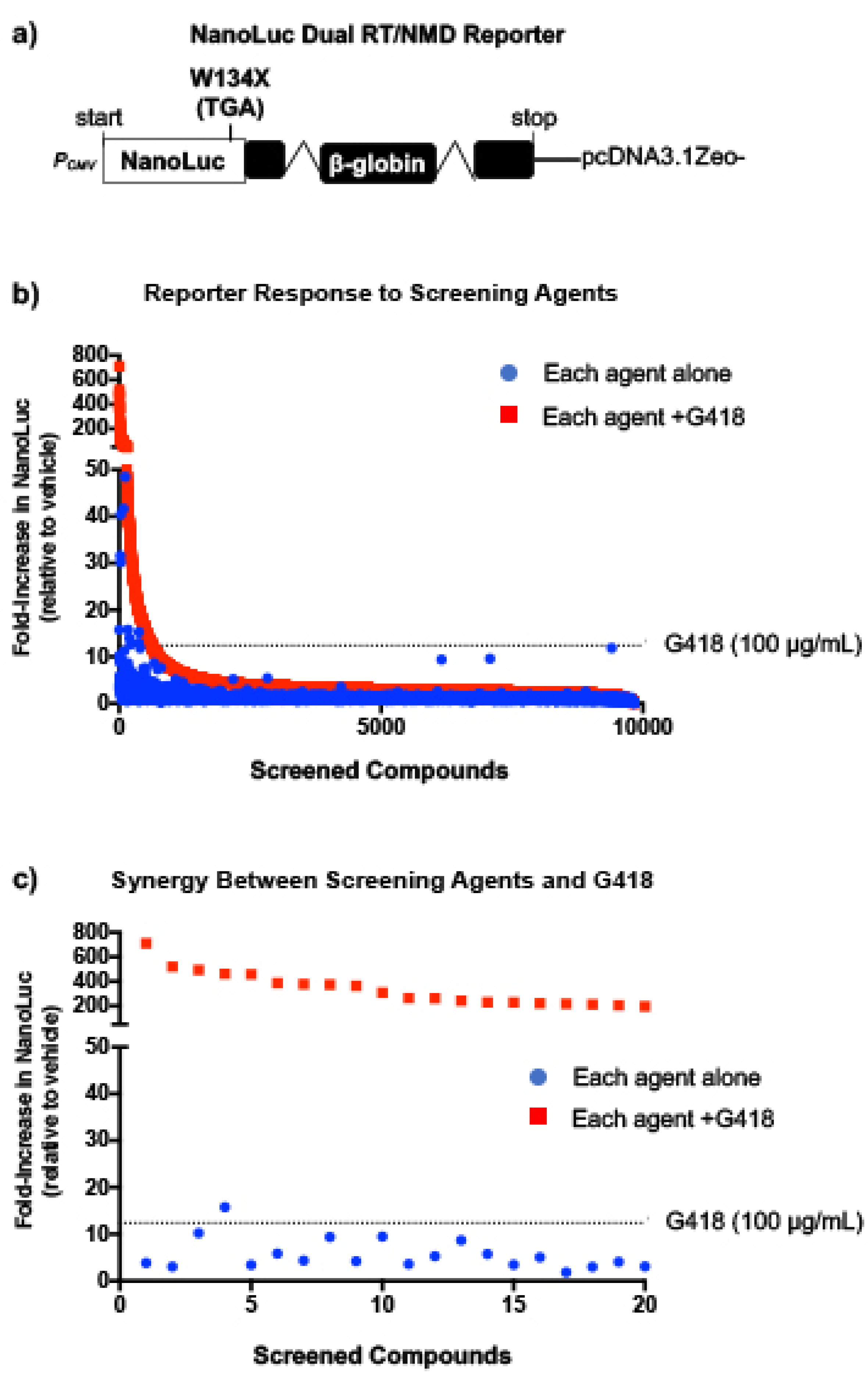
To identify more effective readthrough compounds that could potentially be used to treat MPS I-H (and other diseases), we performed a screen of ~10,000 low-molecular weight compounds for those that suppress PTCs. This screen was performed using NanoLuc reporter cell lines that were previously shown to be sensitive to changes in both readthrough and mRNA abundance, and used to identify eRF1 degrader molecules, a new class of readthrough agent [
11]. Our current screen was performed with each compound alone and in the presence of a low dose of the aminoglycoside, G418. Although G418 is a potent readthrough compound, it cannot be used clinically due to its toxicity. However, it was previously shown that G418 can be used to identify enhancer molecules that do not mediate readthrough themselves but enhance the activity of other readthrough compounds [
12]. In addition, G418, which induces readthrough by increasing ribosomal misreading, has been shown to synergistically increase readthrough when combined with compounds that mediate readthrough via other mechanisms (such as eRF1 [
11] and eRF3 [
13] degraders). We subsequently examined primary hits from this screen for the ability to suppress nonsense mutations associated with MPS I-H.
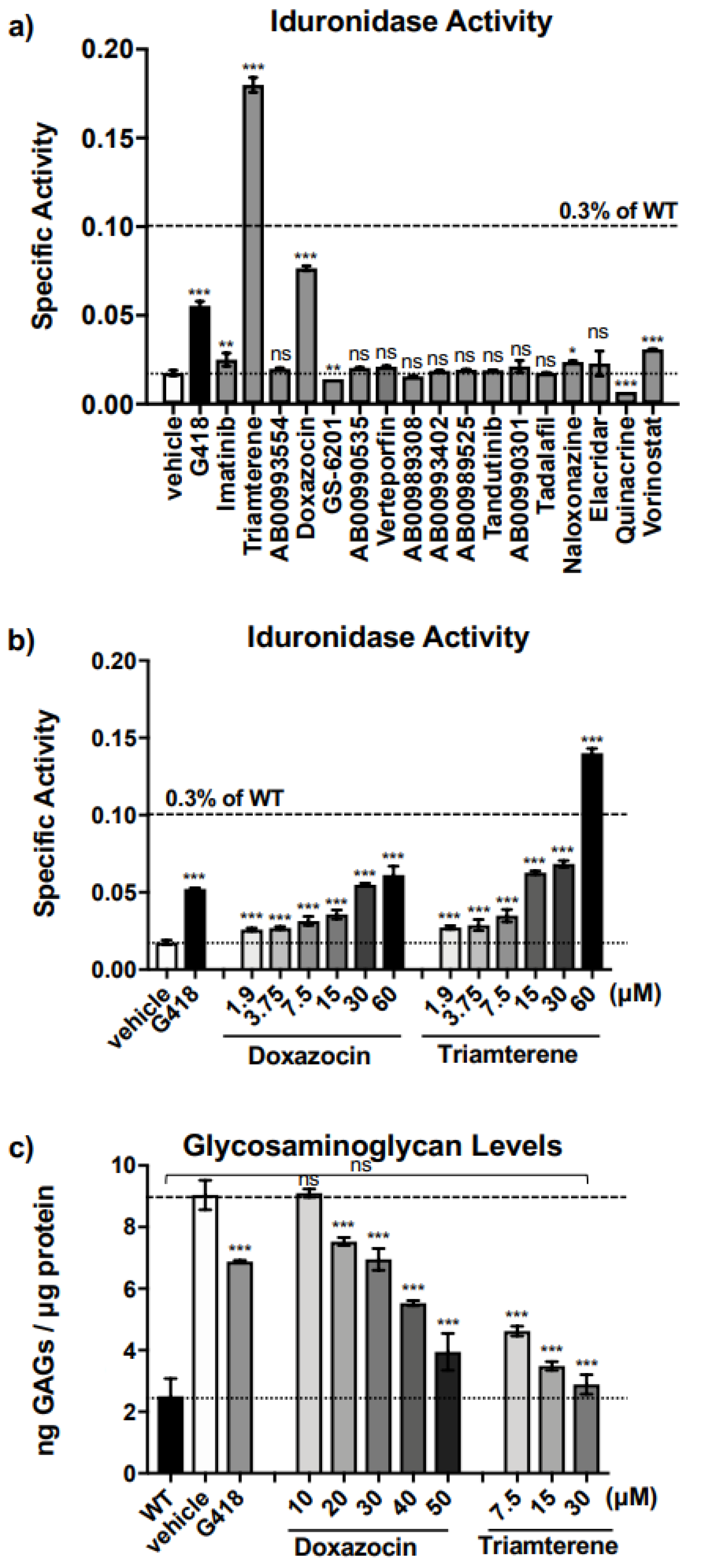
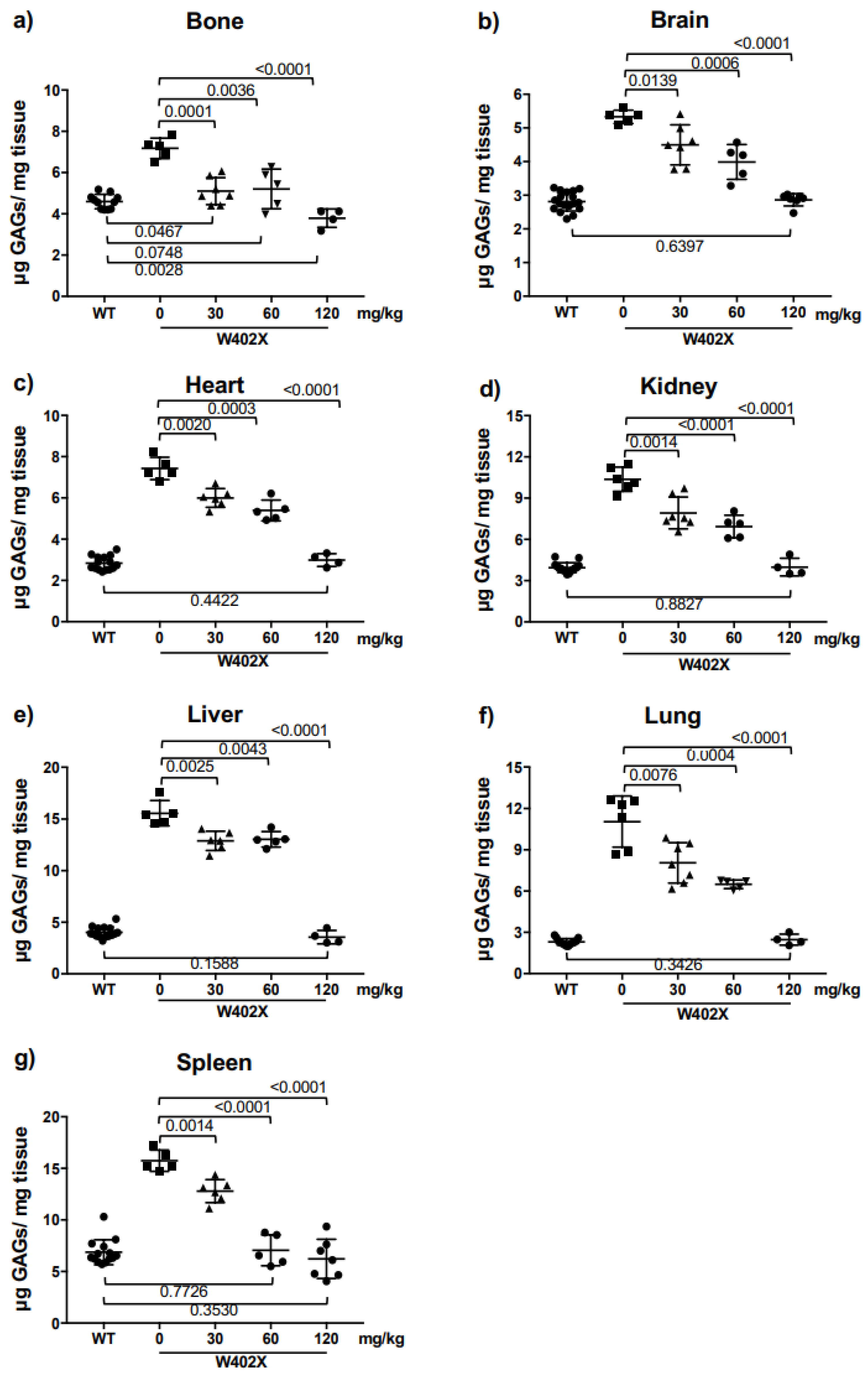
Triamterene, an FDA-approved, potassium-sparing, diuretic used to treat hypertension, was identified as a hit from our screen. Triamterene increased α-L-iduronidase activity in a dose-dependent manner and correspondingly decreased GAG storage in both mouse and human cells. This newly discovered, PTC-associated function of triamterene is independent of the epithelial sodium channel (ENaC) activity, the target of triamterene’s antihypertensive action. Importantly, after two weeks of administering 120 mg/kg triamterene to IDUA-W402X mice, GAGs were completely restored to normal levels in multiple tissues, including the brain and bone. Together, these data suggest that triamterene represents a promising new drug for treating MPS I-H patients who harbor a nonsense mutation.
3. Discussion
Standard of care treatments for MPS I-H includes hematopoietic stem cell transplantation (HSCT), and enzyme replacement therapy (ERT). Both treatments help alleviate some MPS I-H symptoms and extend the patient’s lifespan. However, Aldurazyme
TM, a recombinant form of α-L-iduronidase that is administered as an ERT, cannot cross the blood/brain barrier to prevent the onset of neurological deterioration. While HSCT can halt the progression of neurological phenotypes if administered early (<16-months of age) [
30], it carries significant inherent safety risks. The rate of MPS I-H patients surviving an initial HSCT with efficient engraftment has been reported to be around 70% [
31]. In addition, neither ERT nor HSCT prevent the onset of MPS I-H manifestations in the heart valves, cornea, or bone, which significantly contribute to the morbidity and mortality of MPS I-H patients [
32]. Safer, more effective treatments for MPS I-H are needed.
Nonsense suppression therapy represents a promising approach to treating MPS I-H patients who carry a PTC. Notably, more than 70% of all MPS I-H patients carry a nonsense mutation [
9]. In addition, the threshold for phenotypic improvement of MPS I-H is quite low, with as little as 0.3% of normal α-L-iduronidase activity differentiating between patients with the severe, Hurler form of MPS I from patients from the attenuated, Scheie form [
10], who have a normal intellect and life expectancy. Furthermore, readthrough agents are low molecular weight compounds that may access tissues that are impenetrable to current MPS I-H treatments, such as the brain and bone [
19,
20].
In the current study, we implemented several strategies to identify new, effective readthrough agents. First, we used a NanoLuc-based dual RT/NMD reporter that allowed us to identify not only molecules that stimulate PTC readthrough, but also compounds that stabilize the PTC-containing mRNA by inhibiting NMD or via other mechanisms such as passive mRNA stabilization [
33]. An advantage of this reporter is that it can be used to find compounds (or combinations of compounds) that target multiple PTC suppression mechanisms, a strategy likely to generate more protein function than targeting only a single readthrough mechanism [
6,
8]. Furthermore, we screened each compound alone and combined with a low dose of the aminoglycoside, G418. The presence of G418 during screening enhanced readthrough, allowing us to identify greater numbers of compounds with potential readthrough activity, or with the ability to enhance the activity of other readthrough compounds as previously reported [
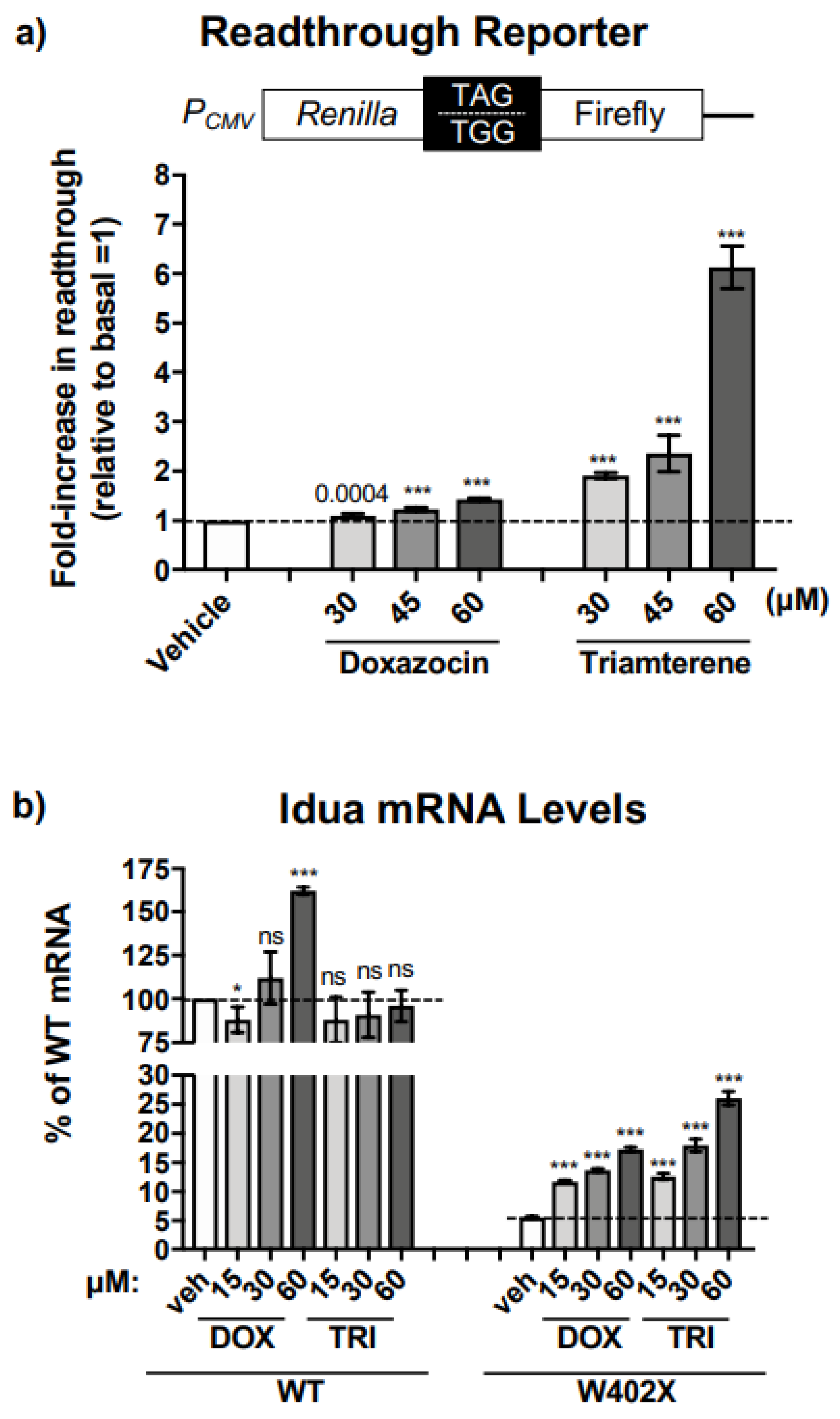
12]. Triamterene was identified as a hit from our screen that alone stimulated readthrough, and in combination with G418, produced a synergistic increase in readthrough in the primary reporter assay screen. However, we did not observe synergy with triamterene and G418 in
IDUA-W402X MEFs. We speculate that certain aspects of translation and/or mRNA turnover might differ between embryonic and differentiated cells which could affect whether synergy occurs. For example, we have found that the extent by which
IDUA-W402X mRNA abundance is reduced by NMD differs between mouse embryonic tissue and tissues from adult mice.
IDUA-W402X mRNA abundance is reduced to around 5% of wild-type levels in MEFs, while in tissues from adult mice,
IDUA-W402X mRNA abundance is reduced to 25–50% of wild-type [
8]. This may be at least one factor that contributes to the lack of synergy in MEFs.
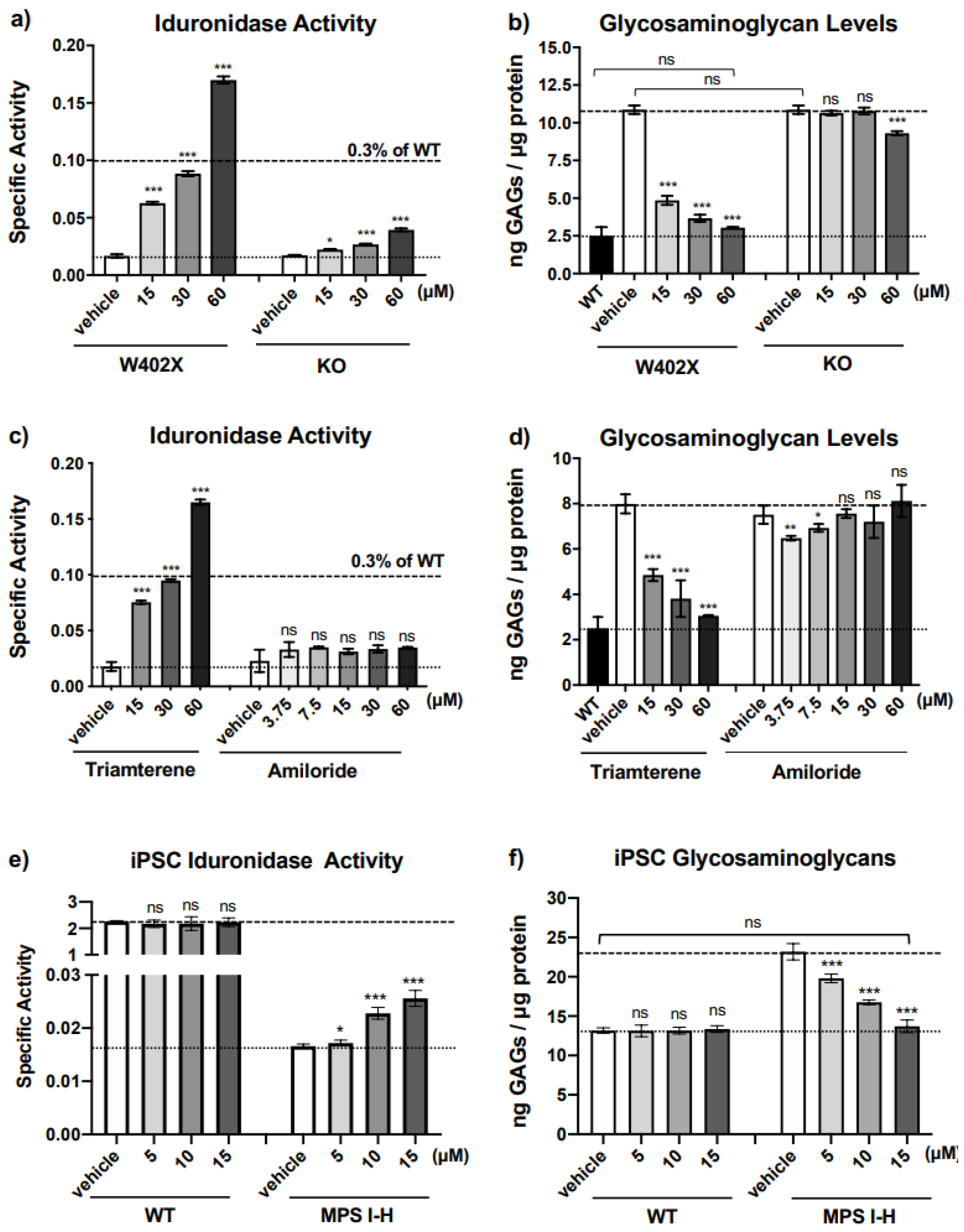
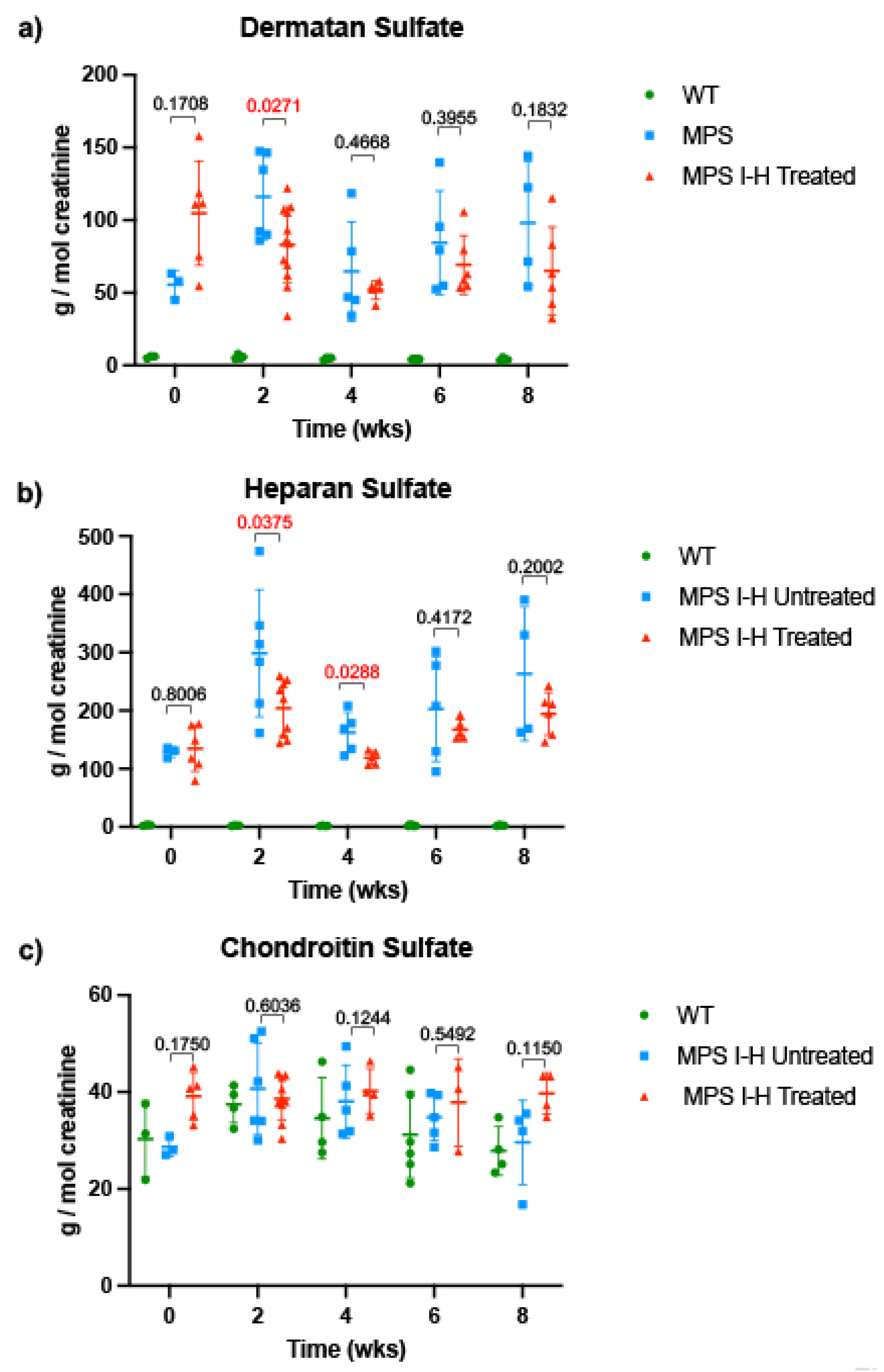
In both in vitro and in vivo
IDUA-W402X MPS I-H models, triamterene restored α-L-iduronidase activity and decreased GAG storage in a dose-dependent manner. Unlike ataluren [
34] or the synthetic aminoglycoside NB84 [
19,
20], which only partially attenuated GAG accumulation in most
IDUA-W402X mouse tissues, a 120 mg/kg dose of triamterene for 2-weeks completely normalized GAG levels in all the tissues examined and significantly reduced DS and HS in urine samples from
IDUA-W402X mice. Even the partial GAG reductions previously observed in NB84-treated
IDUA-W402X mice significantly attenuated the progression of MPS I-H [
19]. Given that triamterene normalized GAG levels in all
IDUA-W402X mouse tissues, it was anticipated that triamterene would also prevent or greatly attenuate the progression of MPS I-H in
IDUA-W402X mice. Unfortunately, this result was not observed; microsome stability assays suggested that triamterene likely undergoes extensive metabolism in mouse liver (
Supplementary Figure S6a), which appears to inactivate its PTC suppression activity. This finding precluded examination of the effects of long-term triamterene treatment in
IDUA-W402X mice. However, this should not be the case in humans since triamterene is much more stable in human microsomal extracts and is routinely used as a diuretic in patients for extended periods of time.
Additional in vitro studies in our IDUA-W402X MEFs indicated that triamterene restores α-L-iduronidase primarily through PTC-dependent mechanisms that at least entail promoting readthrough of the IDUA-W402X PTC and stabilizing the IDUA-W402X mRNA. Furthermore, the PTC suppression activity mediated by triamterene is independent of ENaC function. The ability of triamterene to suppress PTCs in human cells was demonstrated in HEK293 readthrough reporter cells and in MPS I-H patient iPSCs, where triamterene restored sufficient α-L-iduronidase activity to normalize GAG accumulation.
Together, this evidence suggests that we have identified a novel function for triamterene as a PTC suppression agent. Because triamterene is already an FDA-approved drug, repurposing triamterene as a nonsense suppression compound could quickly allow triamterene to enter clinical trials as a readthrough therapy for MPS I-H patients. While the 120 mg/kg dose administered to
IDUA-W402X mice is significantly higher than the clinical dose normally administered to patients, triamterene is metabolized much more extensively in mouse liver microsomes than in human liver microsomes (
Supplementary Figure S6a), suggesting that the triamterene dose that normalized GAGs in
IDUA-W402X mice may be comparable to the range of triamterene dosing that is used clinically. Since ENaC function does not influence the readthrough of the
IDUA-W402X mutation, it may also be possible that modifying the structure of triamterene to increase its potency as a readthrough agent while eliminating its effect on ENaC can reduce potential side effects associated with increased potassium levels. Similar medicinal chemistry approaches have generated safer, more effective aminoglycosides for readthrough [
20,
35,
36,
37]. Altogether, these data suggest that triamterene (or triamterene-like) molecules may be effective readthrough agents for treating MPS I-H. Triamterene could potentially be used as a readthrough agent for other disease models as well. While we mainly focused on the
IDUA-W402X context, it is important to note that triamterene alone increased readthrough by 13-fold in the NanoLuc reporter context. The NanoLuc context (UGA) is significantly different than the
IDUA-W402X context (UAG), suggesting that triamterene likely induces readthrough in a variety of different contexts.
4. Materials and Methods
4.1. Cell Culture
Mouse embryonic fibroblasts (MEFs) were immortalized using the SV40 large T antigen as previously described [
8,
20]. MEFs utilized in this study were derived from homozygous wild-type mice (WT), knock-in MPS I-H mice homozygous for a genomic point mutation that generates a PTC homologous to the
IDUA p.W402X nonsense mutation (
IDUA-W402X) [
18], and knock-out MPS I-H mice carrying an insertional mutation in the mouse
IDUA locus (
IDUA-KO) [
22]. MEF cell lines were cultured at 37 °C with 6.5% CO
2 in Dulbecco’s Modification of Eagle’s Medium containing 4.5 g/L glucose, L-glutamine and sodium pyruvate (Corning Cellgro 10-013-CV, Manassas, VA, USA). This media was supplemented with 100 units/mL penicillin/streptomycin (Corning Cellgro 30-002-Cl), MEM non-essential amino acids (Corning Cellgro 25-025-Cl) at a final concentration of 1% (
v/
v), and fetal bovine serum (Atlanta Biologicals S11150, Flowery Branch, GA, USA) at a final concentration of 10% (
v/
v). HEK293 cells were cultured similarly. Fisher rat thyroid (FRT) cells were cultured in Nutrient Mixture F-12 Coon’s modification media (Sigma F6636, St. Louis, MO, USA) supplemented with 5% fetal bovine serum. For stably transfected HEK293 and FRT cells, 100 units/mL penicillin/streptomycin was added to the media in the absence of zeocin to prevent bacterial contamination. iPSCs were reprogrammed from MPS I-H patient skin fibroblasts [
25] as previously described [
38,
39]. iPSCs were cultured in mTeSR Plus media (Stemcell Technologies #100-0276, Vancouver, BC, Canada) on plates coated with Geltrex (Gibco #A14133-02, Waltham, MA, USA) according to manufacturer instructions.
4.2. Construction of the Readthrough Reporters
The NanoLuc readthrough/NMD (RT/NMD) reporter was constructed as previously described [
11]. The NanoLuc open reading frame was obtained from the pFN[Nluc/CMV/neo] plasmid (Promega CS181701, Madison, WI, USA). Initially, a multi-cloning site was placed into the Xho I/Not I sites of this plasmid using the annealed oligos DB4078 [5′-tcgagccaag cttgcatgcct gcaggtcgact ctagaggatcc ccggggaattcgc-3′] and DB4079 [5′-ggccgcgaat tccccgggga tcctctagag tcgacctgcag gcatgcaagcttggc-3′] to remove the barnase sequence and simplify subsequent cloning. This new construct (pDB1333) was used as a template to generate the W134X (UGA) premature termination codon in the NanoLuc gene (pDB1345) using site-directed mutagenesis with the forward primer DB4144, 5′-gggaccctgt gaaacggcaac-3′ and the reverse primer DB4145, 5′-gttgccgttt cacagggtccc-3′. The final W134X NanoLuc RT/NMD reporter was generated by replacing the
Renilla gene within the
Renilla-β-globin/pcDNA3.1Zeo(−) plasmid (pDB1329) with the W134X NanoLuc gene from pDB1345 using Nhe I/Xho I sites such that the NanoLuc gene was fused in-frame with exon 1 of β-globin.
The construction of the
IDUA-W402X dual-luciferase reporters has been previously described [
20]. The original p2luc dual luciferase readthrough constructs were a gift from Dr. John Atkins [
40]. The p2luc construct was modified to express either the wild-type (WT) mouse
IDUA codon (UGG) that is homologous to the W402 codon in the human
IDUA cDNA, or the
IDUA W402X premature stop codon (UAG), along with three codons of upstream and downstream mouse
IDUA sequence. Complementary oligonucleotides for generating the
IDUA-W402X construct: [5′-tcgacg gaacaa ctctag gcagag gtcg-3′; 5′-gatccg acctct gcctag agttgt tccg-3′], and the WT
IDUA construct: [5′-tcgacg gaacaa ctctgg gcagag gtcg-3′; 5′-gatccg acctct gcccag agttgt tccg-3′] were annealed to generate double-stranded DNA fragments that were ligated into the Sal I and BamH I restriction sites of the p2luc vector, yielding the
IDUA-W402X (pDB1134), and
IDUA-WT (pDB1133) p2luc constructs. Using the Not I and Nhe I restriction sites, the
IDUA-W402X and
IDUA-WT dual luciferase constructs were subcloned into pcDNA3.1Zeo(+) to generate pDB1325 and pDB1326, respectively, for stable expression in mammalian cells.
4.3. Generation of Readthrough Reporter Cells
The W134X NanoLuc RT/NMD reporter was linearized with BgI II and transfected into Fisher rat thyroid (FRT) cells using Lipofectamine 2000 (Invitrogen 11668, Austin, TX, USA). Stably transfected FRT cells were selected by the addition of 800 μg/mL zeocin to the growth media for 2–3 weeks. Stable FRT reporter cell lines were maintained by the addition of 200 μg/mL of zeocin to the media. Monoclonal FRT cell lines were established by collecting cells from single-cell-derived colonies and expanding them.
The IDUA-W402X and IDUA-WT dual luciferase constructs (pDB1325 and pDB1326, respectively) were linearized with Bgl II and transfected into human embryonic kidney (HEK) 293 cells, which were cultured in the presence of 200 μg/mL of zeocin for 2–3 weeks to select for cells stably expressing the reporters. Reporter HEK293 cells were also maintained in 200 μg/mL of zeocin.
4.4. High Throughput Screening Assay
This screen was similarly conducted with the W134X NanoLuc RT/NMD-FRT Monoclonal #14 cells as previously described [
11]. The reporter cells were cultured in Coon’s F-12 Medium (Cedar Lanes Labs Catalog #F 0855, Burlington, ON, Canada) supplemented with 5% heat-inactivated fetal bovine serum (Omega Scientific Catalog #FB-11, Tarzana, CA, USA) and 1% L-glutamine, harvested, and then re-suspended at 400,000 cells/mL in media supplemented with 5% heat-inactivated fetal bovine serum, 1% Pen/Strep/Glut (Corning Catalog #30-009CI) and 1% HEPES (Gibco Catalog #15630-080). Using Beckman Coulter FX, 5 µL of compounds/controls were added to wells of 384-well white, opaque bottom plates (Corning Catalog #3570BC). Control and screening compounds were diluted in media with DMSO (Sigma-Aldrich Catalog #D8418) at 6× concentration. The wells of columns 1 & 2 were used as the negative (cell) control and contained 5 µL of media with DMSO. The wells of columns 23 & 24 were used as the positive control and contained 5 µL of G418 (Corning Catalog #30-234-CI) diluted in media with DMSO. 5 µL of screening compounds were added to the wells of columns 3–22. Using Thermo Matrix wellmate dispenser, 25 µL of W134X cells were added to the wells of columns 1, 2, 23, & 24 of the assay plates. For test wells, 25 µL of W134X cells were added to the wells of columns 3–22. For the screening condition in the presence of G418, this reagent was added to the cells before dispensing to give a final concentration of 100 µg/mL. The final concentration of cells was 10,000 cells per well with a final assay volume of 30 µL and a final concentration of DMSO concentration of 0.6%. G418 was used as the positive control condition with a final concentration of 300 µg/mL. Compounds from 2 libraries were tested on this screen. Library compounds purchased from Enamine (Monmouth Junction, NJ, USA) were screened at 30 µg/mL final concentration. A library of FDA-approved compounds was screened at 60 µM final concentration. The assay plates were gently tapped and placed in a 37 °C with 5% CO
2, high humidity incubator for 48 hrs. After incubation, the plates were cooled to room temperature. Using the Biomek Benchtrak robot, 30 µL of room temperature NanoGlo reagent (Promega Catalog #N1150) diluted 1:50 was added to all the wells of the assay plate. After 10 min, the luminescence was measured using a Perkin Elmer Envision reader. Cell toxicity was monitored using a Celltiter-Glo Luminescent Cell Viability Assay (Promega #G7570).
4.5. Luciferase Readthrough Assays
Equal numbers of the W134X NanoLuc RT/NMD monoclonal reporter cells were seeded into 96-well plates and cultured without zeocin for 24 h. The cells were then treated with screening compounds for 24 h. Dimethyl sulfoxide (DMSO) served as the vehicle in which compounds were administered at a final concentration of 60 μM or 30 μg/mL. After treatment, the cells were washed once with phosphate-buffered saline (PBS), followed by incubation with passive lysis buffer (PLB) (Promega, E1941) for 20 min at room temperature. NanoLuc activity in cell lysates was assayed with the Nano-Glo Luciferase Assay System (Promega, N1120) using the GloMax® Discover System (Promega). NanoLuc activity was normalized to the lysate protein concentration, which was determined by the Bio-Rad Protein Assay Reagent (Bio-Rad 500-006, Hercules, CA, USA).
HEK293 dual luciferase reporter cells were similarly cultured, treated with screening compounds, and lysed as described above. Dual luciferase activity was assayed with the Dual Luciferase Assay System (Promega) using the GloMax Multi Detection System (Promega). The percent readthrough was calculated as the ratio of firefly/Renilla luciferase units expressed from the W402X construct relative to the WT construct × 100.
4.6. α-L-iduronidase Activity Assay
MEFs or iPSCs were seeded into 6-well culture dishes at a density of 1.5 × 10
5 cells per well. Cells were grown to 50% confluency and then treated with screening agents for 48 h. Cells were subsequently washed with PBS and lysed in Mammalian Protein Extraction Reagent (Thermo Scientific P178501, Waltham, MA, USA) containing a protease inhibitor cocktail (Complete Roche 11873580001, Indianapolis, IN, USA). The total protein concentration was determined using the Bio-Rad Protein Assay. Approximately 75 μg of total lysate protein were incubated in a 70 μL reaction containing 0.12 mM 4-methyl-umbelliferyl-α-L-iduronide (Gold Biotech M-570-5, Saint Louis, MO, USA) and 0.42 mg/mL of D-saccharic acid 1,4-lactone monohydrate (a β-glucuronidase inhibitor) (Sigma S0375) in 130 mM sodium formate buffer, pH 3.5. The reaction was incubated for 72 h at 37 °C and then quenched with 300 μL of glycine buffer, pH 10.8. 200 μL of each sample was transferred to a 96-well plate (Corning 3631) and fluorescence was measured at an excitation = 365 nm and an emission = 450 nm using the GloMax
® Discover System (Promega). Free acid 4-methylumbelliferone (FMU) (Sigma M1381) in glycine buffer was used to generate a standard curve. Specific activity was calculated as nanomoles of FMU released per microgram of protein per hour. α-L-iduronidase activity remained linear over the 72-h incubation time with the substrate concentration remaining in excess over the incubation period. As shown in
Supplementary Figure S1, a standard curve was also established with the fluorescent 4-methyl-umbellyferone substrate in the presence of triamterene. Fluorescence associated with triamterene occurred only at high concentrations, which was not expected to be present in cell or tissue lysates with the concentrations used to treat MEFs or mice.
4.7. Glycosaminoglycan Quantitation in Cultured Cells
MEFs or iPSCs were seeded into 6-well culture dishes at a density of 5 × 10
4 cells per well. Cells were grown to 50% confluency and then treated with screening agents for 48 h. Subsequently, the media was removed and the cells were scraped into 500 µL of 0.2 mg/mL papain in pH 6.5 phosphate buffer containing 0.6 mg/mL cysteine. The cell lysates were incubated in the papain solution at 65 °C with gentle agitation for 3 h. Samples were briefly microfuged for 10 min at 10,000×
g at room temperature to remove debris. GAG levels were determined using the Blyscan Sulfated GAG Assay (Biocolor Ltd., Carrickfergus, UK). Briefly, 50 µL of each supernatant was mixed with 500 µL of the Blyscan Dye Reagent to bind sulfated GAGs. The dye-bound GAGs were pelleted by microfuging for 10 min at 10,000×
g at room temperature. 500 µL of the Blyscan Dye Dissociation Reagent was added to each sample to dissociate the GAGs from the dye. 200 µL of each sample was transferred to a 96-well plate (Corning 3631) and the absorbance was measured at a wavelength of 650 nm absorbance using a GloMax
® Discover System (Promega). The total amount of sulfated GAGs precipitated from each sample was determined from a chondroitin 4-sulfate (Sigma C9819) standard curve. The total protein concentration in each lysate was determined using the Bio-Rad Protein Assay (5000006) from a standard curve generated using bovine serum albumin. The GAG levels are expressed as nanograms of GAGs per microgram of total protein. Standard curves for the GAG dye-binding assay using chondroitin 6-sulfate (Sigma C4384) (
Supplementary Figure S2a), dermatan sulfate (EMD Biosciences 263301) (
Supplementary Figure S2b), and heparan sulfate (Sigma-Aldrich H7640) (
Supplementary Figure S2c) were performed in the absence and presence of triamterene (at concentrations 10- to 20-fold greater than the typical patient target serum concentrations with oral dosing [
41]) to confirm that triamterene does not interfere with the binding of any of the different types of GAGs in the dye-binding assay.
4.8. Quantitation of Steady-State IDUA mRNA Levels
Total RNA was isolated from MEFs using the Ambion RiboPure kit (Fisher Scientific AM1924) and DNase-treated using the Turbo DNA-Free kit (Fisher Scientific AM1907). Polyadenylated RNA was reverse transcribed into cDNA in a 50 μL reaction containing 1 μg of total RNA; 0.5 mg/mL oligo dT; 1.2 mM dNTPs; 40U RNasin (Promega N2511); 10 mL of 5× AMV RT buffer and 40U AMV reverse transcriptase (Promega PM-M9004). RT reactions were incubated at 42 °C for 1.5 h, and then heat-inactivated at 65 °C for 15 min. The cDNA was ethanol precipitated and subjected to qPCR in a 25 μL reaction containing 12.5 μL iQ SYBR Green Supermix (Bio-Rad 1708882); 0.2 mM of each forward and reverse primer; and 2 μg of cDNA. The following primer sets (forward = Pf and reverse = Pr) were used:
IDUA Pf: 5′-tgacaa tgcctt cctgag ctacca-3′ and
IDUA Pr: 5′-tgactg tgagta ctggct ttcgca-3′;
Gapdh Pf: 5′-ttccag tatgac tccact cacgg-3′ and
Gapdh Pr: 5′-tgaaga caccag tagact ccacgac-3′;
Rpl13a Pf: 5′-atgaca agaaaa agcggatg-3′ and
Rpl13a Pr: 5′-cttttc tgcctg tttccgta-3′. The qPCR was performed using the CFX96 Real-Time PCR Detection System (Bio-Rad) using a program that included an initial 3-min denaturation step at 95 °C followed by 40 repeated cycles of a 10 s denaturation step at 95 °C and a 30 s annealing/extension step at 55 °C. Melt curve analysis was initially performed with each primer set to verify that only one gene product was generated from the PCR reactions. A standard curve was also performed using each primer set to ensure that under the PCR conditions used, the efficiency ranged between 90–110%. The average quantification cycle (Cq) was determined for each mRNA, and mRNA quantification was performed using the Livak (DDCq) method [
21], where
Gapdh and
Rpl13a served as normalization controls. Cq values among the different samples for the various transcripts ranged from 10–34. The qPCR was performed using at least 8–12 replicates for each gene product from each sample.
4.9. Microsome Stability Assay
The amount of unaltered compound was determined in human and mouse liver microsomes (0.5 mg/mL protein) at 37 °C with 1.0 µM analyte. The reaction was quenched with acetonitrile after 0, 5, 10, 20, 30, and 60 min. The samples were then analyzed by LC-MS/MS methodology as previously described, with diclofenac and midazolam used as positive controls [
42,
43].
4.10. Animal Treatment
Triamterene was suspended in a vehicle composed of 0.5% methylcellulose in an artificially sweetened Kool-Aid solution. This mixture was administered orally to 8- to 10-week-old mice via once-daily gavage for a total of 14 days. Doses ranged from 30–120 mg/kg. Control mice were administered the Kool-Aid vehicle alone. All animals were provided standard chow and water ad libritum. All animal work was conducted according to relevant national and international guidelines. All animal protocols used in this study were reviewed and approved by the UAB IACUC (protocol IACUC-20364).
4.11. GAG Quantitation in Mouse Tissues
This assay was performed as previously described [
8,
19,
20]. Tissues were homogenized using a Tissue Tearor homogenizer in chloroform: methanol (2:1
v/
v). Defatted tissue was dried in a speedvac, weighed, and then suspended in 100 mM dibasic sodium phosphate, pH 6.5 containing 0.6 mg/mL cysteine and 2 mg/mL papain (Sigma P4762). The mixture was digested at 60 °C for 18–24 h with constant agitation. The samples were then microfuged at 10,000×
g for 15 min and the supernatant was used to quantitate the tissue GAGs using the Blyscan Sulfated GAG Assay (Biocolor Ltd., Carrickfergus, UK). The total amount of sulfated GAGs precipitated from each sample was determined from a standard curve using chondroitin 4-sulfate (Sigma C9819). The GAG levels are expressed as micrograms of GAGs per milligram of defatted, dried tissue.
4.12. GAG Determination in Mouse Urine
Urine was collected from individual mice via spontaneous urination into a weigh boat, filtered using a 0.22 μm syringe filter, and then stored at −80°C until assay. Urinary glycosaminoglycans (chondroitin sulfate, dermatan sulfate, and heparan sulfate) were quantified using UPLC MSMS as previously described [
44]. Briefly, GAGs were digested using methanolic HCl into three unique disaccharides that are specific for chondroitin sulfate, dermatan sulfate, or heparan sulfate. The dimers were separated by ultra-performance liquid chromatography (UPLC) and analyzed by electrospray ionization tandem mass spectrometry (MS/MS) using selected reaction monitoring. Calibration curves were generated using commercially available standards (Sigma).
4.13. Statistics
All statistics were calculated with ANOVA models using Prism 9 software. Sample sizes are indicated in figure legends and/or the materials and methods and were selected for each assay based on previous data. Among the biochemical endpoints, the GAG assay was the most variable and 8 mice per group were required to detect a 20% decrease in brain GAGs (with 85% power at a 0.05 significance level). We will therefore include 8 mice per cohort to ensure that our sample size supports adequate statistical power.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





