
Anti-Spike Antibodies Present in the Milk of SARS-CoV-2 Vaccinated Mothers Are Complement-Activating
 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Results
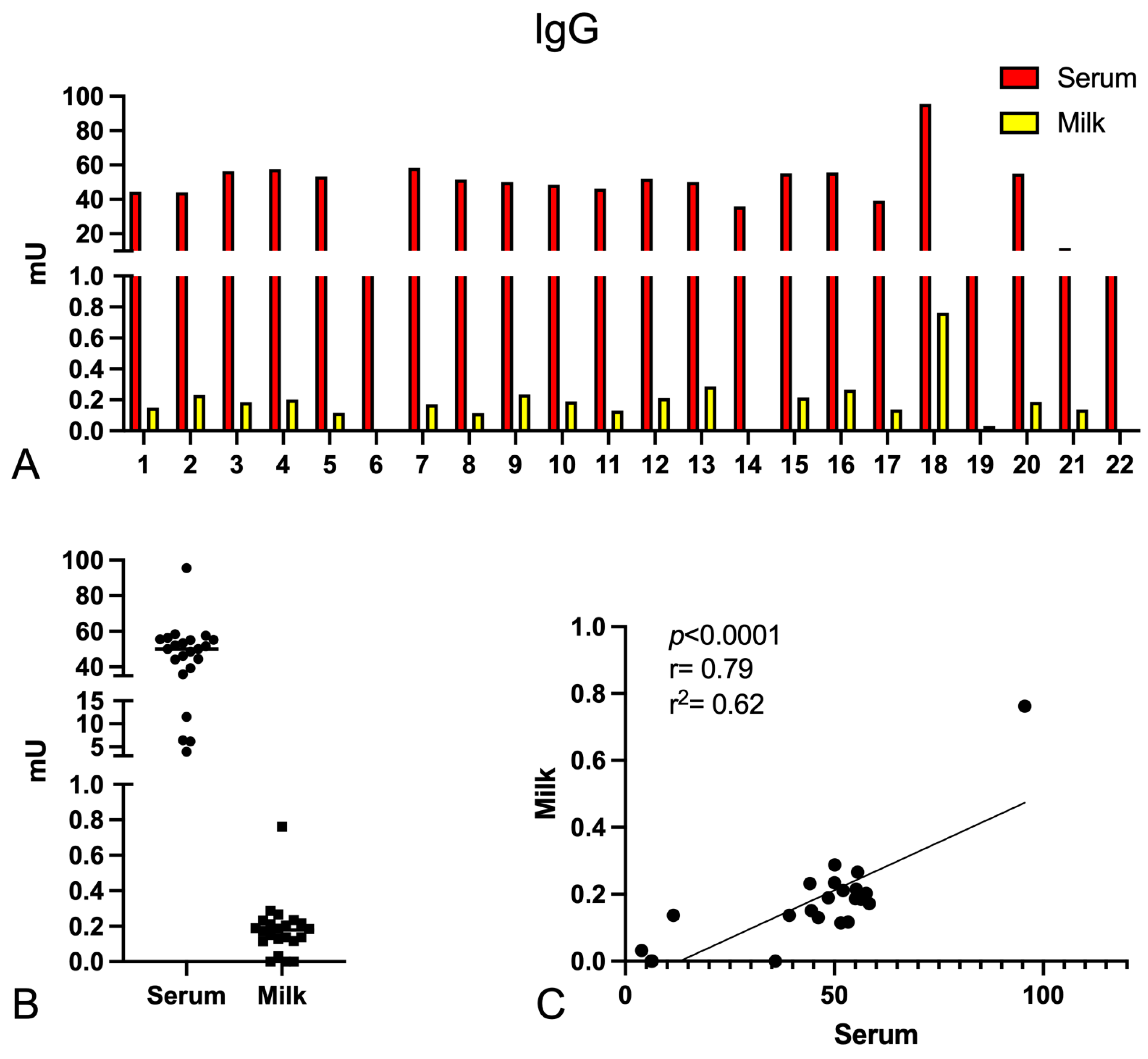
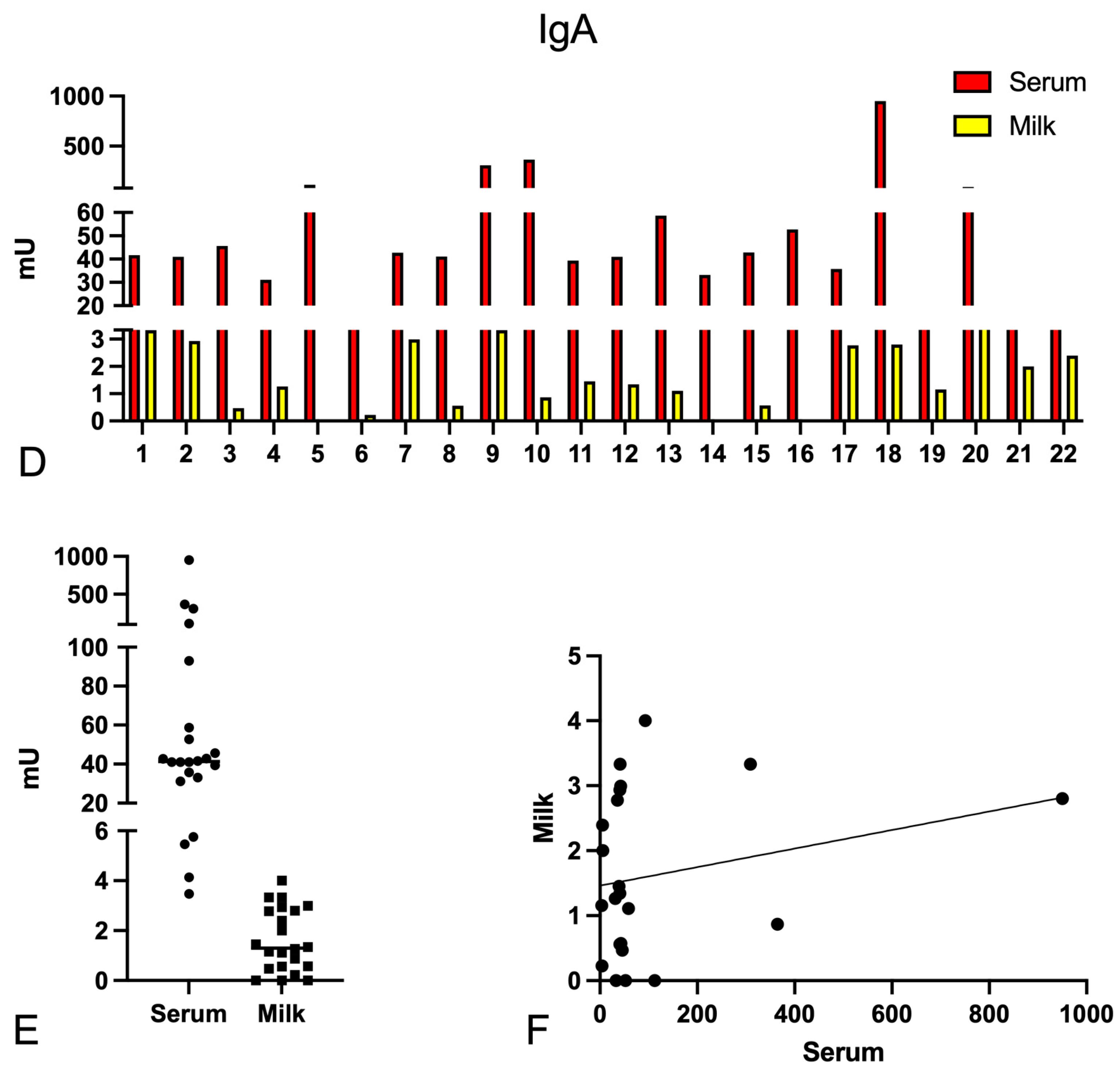
2.1. Determination of the Anti-Spike-Specific Antibody Titre in Serum and Milk of Vaccinated Mothers
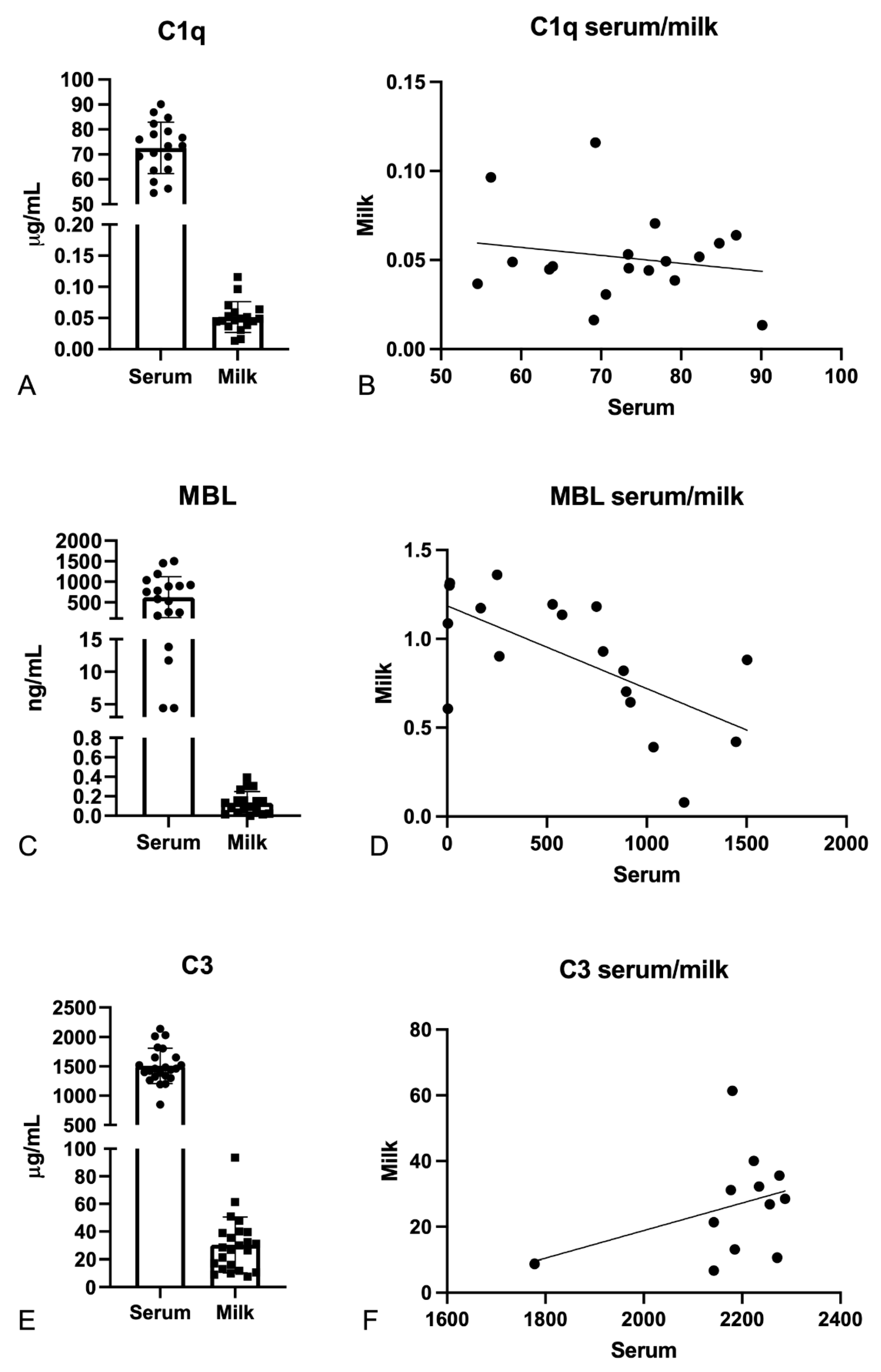
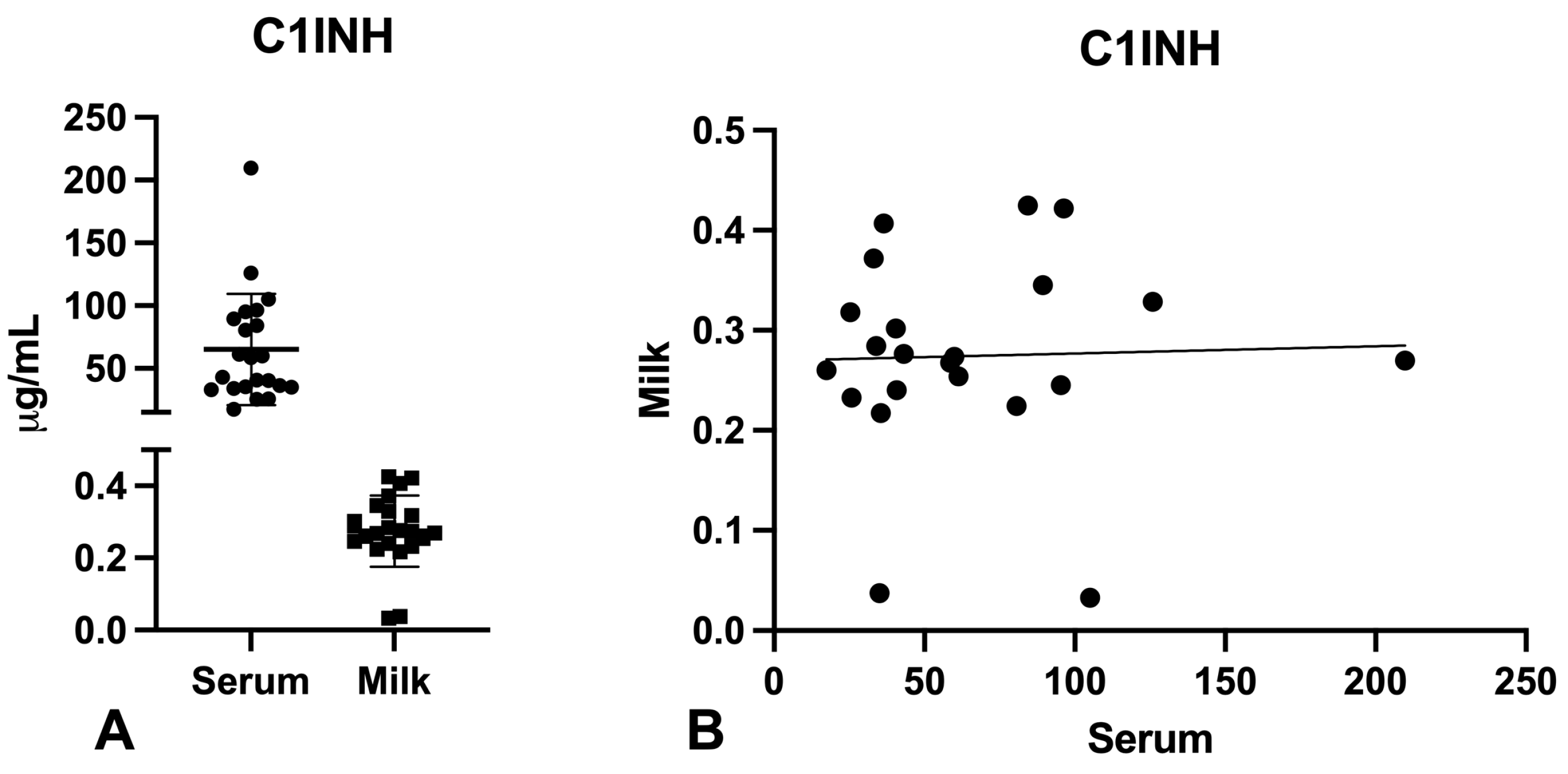
2.2. Evaluation of C1q, MBL, and C3 Levels in the Serum and Milk of Vaccinated Mothers
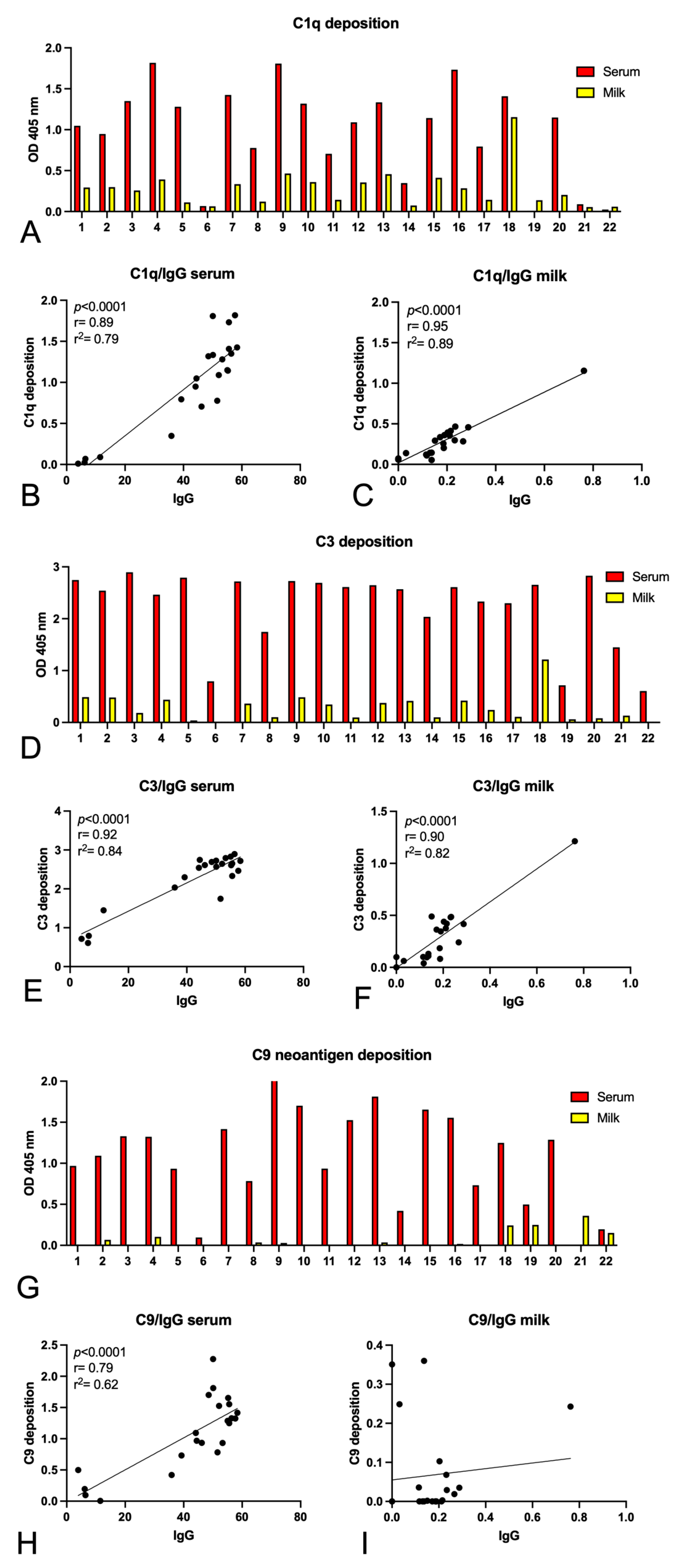
2.3. Complement Activation by Igs Anti-Spike Antibodies Present in Serum and Milk
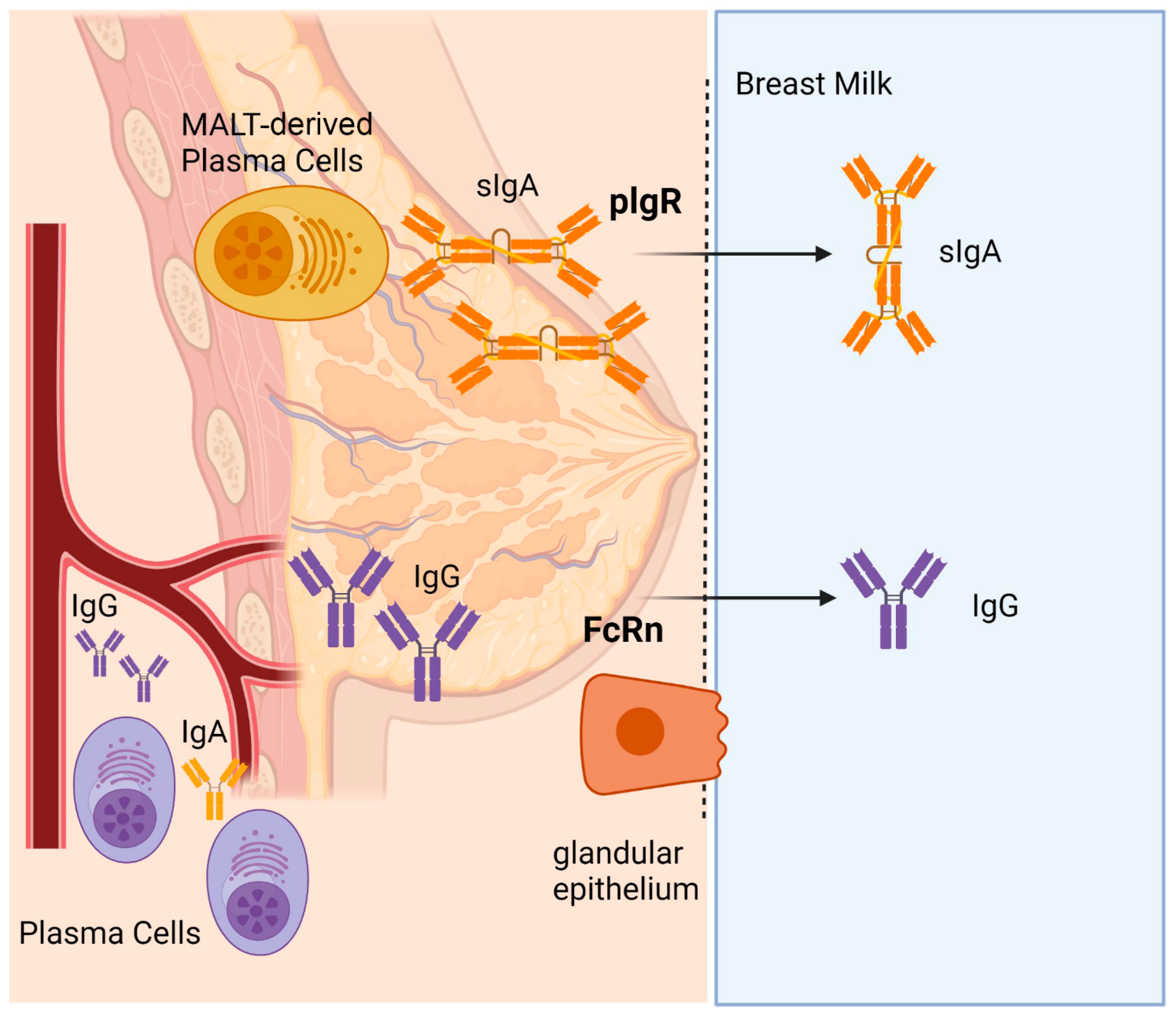
3. Discussion
4. Materials and Methods
4.1. Patient Enrolment and Sample Collection
4.2. Quantification of Anti-Spike IgG and IgA by Enzyme-Linked Immunosorbent Assay (ELISA)
4.3. Measurement of C1q, C3, and MBL Levels by Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Evaluation of Complement Pathway Functionality in Serum and Milk
4.5. Evaluation of Complement Activation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walport, M.J. Complement. First of two parts. N. Engl. J. Med. 2001, 344, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Walport, M.J. Complement. Second of two parts. N. Engl. J. Med. 2001, 344, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merle, N.S.; Noé, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part II: Role in Immunity. Front. Immunol. 2015, 6, 257. [Google Scholar] [CrossRef] [Green Version]
- Trouw, L.; Blom, A.; Gasque, P. Role of complement and complement regulators in the removal of apoptotic cells. Mol. Immunol. 2008, 45, 1199–1207. [Google Scholar] [CrossRef]
- Daha, M.R. Role of Complement in Innate Immunity and Infections. Crit. Rev. Immunol. 2010, 30, 47–52. [Google Scholar] [CrossRef]
- Goldberg, B.S.; Ackerman, M.E. Antibody-mediated complement activation in pathology and protection. Immunol. Cell Biol. 2020, 98, 305–317. [Google Scholar] [CrossRef]
- Jefferis, R. Isotype and glycoform selection for antibody therapeutics. Arch. Biochem. Biophys. 2012, 526, 159–166. [Google Scholar] [CrossRef]
- Roos, A.; Bouwman, L.H.; van Gijlswijk-Janssen, D.J.; Faber-Krol, M.C.; Stahl, G.L.; Daha, M.R. Human IgA Activates the Complement System Via the Mannan-Binding Lectin Pathway. J. Immunol. 2001, 167, 2861–2868. [Google Scholar] [CrossRef] [Green Version]
- Medjeral-Thomas, N.R.; Cook, H.T.; Pickering, M.C. Complement activation in IgA nephropathy. Semin. Immunopathol. 2021, 43, 679–690. [Google Scholar] [CrossRef]
- Afzali, B.; Noris, M.; Lambrecht, B.N.; Kemper, C. The state of complement in COVID-19. Nat. Rev. Immunol. 2021, 22, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Mangogna, A.; Balduit, A.; Aghamajidi, A.; Ricci, G.; Kishore, U.; Bulla, R. COVID-19, Pre-Eclampsia, and Complement System. Front. Immunol. 2021, 12, 775168. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Saha, S.; Butt, M.T.; Modi, R.K.; George, A.J.T.; Kishore, U. SARS-CoV-2: Pathogenic Mechanisms and Host Immune Response. Adv. Exp. Med. Biol. 2021, 1313, 99–134. [Google Scholar] [CrossRef] [PubMed]
- Macor, P.; Durigutto, P.; Mangogna, A.; Bussani, R.; De Maso, L.; D’Errico, S.; Zanon, M.; Pozzi, N.; Meroni, P.L.; Tedesco, F. Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients. Biomedicines 2021, 9, 1003. [Google Scholar] [CrossRef]
- Ali, Y.M.; Ferrari, M.; Lynch, N.J.; Yaseen, S.; Dudler, T.; Gragerov, S.; Demopulos, G.; Heeney, J.L.; Schwaeble, W.J. Lectin Pathway Mediates Complement Activation by SARS-CoV-2 Proteins. Front. Immunol. 2021, 12, 714511. [Google Scholar] [CrossRef]
- Holter, J.C.; Pischke, S.E.; de Boer, E.; Lind, A.; Jenum, S.; Holten, A.R.; Tonby, K.; Barratt-Due, A.; Sokolova, M.; Schjalm, C.; et al. Systemic complement activation is associated with respiratory failure in COVID-19 hospitalized patients. Proc. Natl. Acad. Sci. USA 2020, 117, 25018–25025. [Google Scholar] [CrossRef]
- Rosenbloom, J.I.; Raghuraman, N.; Carter, E.B.; Kelly, J.C. Coronavirus disease 2019 infection and hypertensive disorders of pregnancy. Am. J. Obstet. Gynecol. 2021, 224, 623–624. [Google Scholar] [CrossRef]
- Ellington, S.; Olson, C.K. Safety of mRNA COVID-19 vaccines during pregnancy. Lancet Infect. Dis. 2022, 22, 1514–1515. [Google Scholar] [CrossRef]
- Prasad, S.; Kalafat, E.; Blakeway, H.; Townsend, R.; O’Brien, P.; Morris, E.; Draycott, T.; Thangaratinam, S.; Le Doare, K.; Ladhani, S.; et al. Systematic review and meta-analysis of the effectiveness and perinatal outcomes of COVID-19 vaccination in pregnancy. Nat. Commun. 2022, 13, 2414. [Google Scholar] [CrossRef]
- Amanat, F.; Krammer, F. SARS-CoV-2 Vaccines: Status Report. Immunity 2020, 52, 583–589. [Google Scholar] [CrossRef]
- Le, T.T.; Andreadakis, Z.; Kumar, A.; Román, R.G.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Uniacke-Lowe, T.; Ryan, A.C.; Kelly, A.L. The composition and physico-chemical properties of human milk: A review. Trends Food Sci. Technol. 2021, 112, 608–621. [Google Scholar] [CrossRef]
- Walker, A. Breast Milk as the Gold Standard for Protective Nutrients. J. Pediatr. 2010, 156, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.Å. Session 1: Feeding and infant development Breast-feeding and immune function. Proc. Nutr. Soc. 2007, 66, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, S.J.; Dickson, J.S.; Barnhart, H.M.; Toledo, R.T.; Eiten-Miller, R.R. IgA, IgG, IgM and Lactoferrin Contents of Human Milk During Early Lactation and the Effect of Processing and Storage. J. Food Prot. 1983, 46, 4–7. [Google Scholar] [CrossRef]
- Ogundele, O.M. Role and significance of the complement system in mucosal immunity: Particular reference to the human breast milk complement. Immunol. Cell Biol. 2001, 79, 1–10. [Google Scholar] [CrossRef]
- Narayanaswamy, V.M.; Pentecost, B.T.; Schoen, C.N.; Alfandari, D.; Schneider, S.S.; Baker, R.B.; Arcaro, K.F. Neutralizing Antibodies and Cytokines in Breast Milk After Coronavirus Disease 2019 (COVID-19) mRNA Vaccination. Obstet. Gynecol. 2021, 139, 181–191. [Google Scholar] [CrossRef]
- COVID-19 Vaccines. In Drugs and Lactation Database (LactMed); National Institute of Child Health and Human Development: Bethesda, MD, USA, 2006.
- Yeo, K.T.; Ni Chia, W.; Tan, C.W.; Ong, C.; Yeo, J.G.; Zhang, J.; Poh, S.L.; Lim, A.J.M.; Sim, K.H.Z.; Sutamam, N.; et al. Neutralizing Activity and SARS-CoV-2 Vaccine mRNA Persistence in Serum and Breastmilk After BNT162b2 Vaccination in Lactating Women. Front. Immunol. 2022, 12, 783975. [Google Scholar] [CrossRef]
- Olearo, F.; Radmanesh, L.-S.; Felber, N.; von Possel, R.; Emmerich, P.; Pekarek, N.; Pfefferle, S.; Nörz, D.; Hansen, G.; Diemert, A.; et al. Anti-SARS-CoV-2 antibodies in breast milk during lactation after infection or vaccination: A cohort study. J. Reprod. Immunol. 2022, 153, 103685. [Google Scholar] [CrossRef]
- Huang, F.S. Are SARS-CoV-2 Antibodies Detectable in Human Milk After Vaccination Against COVID-19? J. Pediatr. Infect. Dis. Soc. 2022, 11, 126. [Google Scholar] [CrossRef]
- Muyldermans, J.; De Weerdt, L.; De Brabandere, L.; Maertens, K.; Tommelein, E. The Effects of COVID-19 Vaccination on Lactating Women: A Systematic Review of the Literature. Front. Immunol. 2022, 13, 852928. [Google Scholar] [CrossRef] [PubMed]
- Young, B.E.; Seppo, A.E.; Diaz, N.; Rosen-Carole, C.; Nowak-Wegrzyn, A.; Vasquez, J.M.C.; Ferri-Huerta, R.; Nguyen-Contant, P.; Fitzgerald, T.; Sangster, M.Y.; et al. Association of Human Milk Antibody Induction, Persistence, and Neutralizing Capacity With SARS-CoV-2 Infection vs mRNA Vaccination. JAMA Pediatr. 2022, 176, 159. [Google Scholar] [CrossRef] [PubMed]
- Juncker, H.G.; Mulleners, S.J.; Ruhé, E.J.; Coenen, E.R.; Bakker, S.; van Doesburg, M.; Harinck, J.E.; Rood, R.D.; Bouhuijs, J.H.; Oomen, M.; et al. Comparing the human milk antibody response after vaccination with four COVID-19 vaccines: A prospective, longitudinal cohort study in the Netherlands. eClinicalMedicine 2022, 47, 101393. [Google Scholar] [CrossRef] [PubMed]
- Selma-Royo, M.; Bäuerl, C.; Mena-Tudela, D.; Aguilar-Camprubí, L.; Pérez-Cano, F.J.; Parra-Llorca, A.; Lerin, C.; Martínez-Costa, C.; Collado, M.C. Anti-SARS-CoV-2 IgA and IgG in human milk after vaccination is dependent on vaccine type and previous SARS-CoV-2 exposure: A longitudinal study. Genome Med. 2022, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Jarlhelt, I.; Nielsen, S.K.; Jahn, C.X.H.; Hansen, C.B.; Pérez-Alós, L.; Rosbjerg, A.; Bayarri-Olmos, R.; Skjoedt, M.-O.; Garred, P. SARS-CoV-2 Antibodies Mediate Complement and Cellular Driven Inflammation. Front. Immunol. 2021, 12, 767981. [Google Scholar] [CrossRef]
- Lamerton, R.E.; Marcial-Juarez, E.; Faustini, S.E.; Perez-Toledo, M.; Goodall, M.; Jossi, S.E.; Newby, M.L.; Chapple, I.; Dietrich, T.; Veenith, T.; et al. SARS-CoV-2 Spike- and Nucleoprotein-Specific Antibodies Induced After Vaccination or Infection Promote Classical Complement Activation. Front. Immunol. 2022, 13, 838780. [Google Scholar] [CrossRef]
- Charepe, N.; Gonçalves, J.; Juliano, A.M.; Lopes, D.G.; Canhão, H.; Soares, H.; Serrano, E.F. COVID-19 mRNA vaccine and antibody response in lactating women: A prospective cohort study. BMC Pregnancy Childbirth 2021, 21, 632. [Google Scholar] [CrossRef]
- Romero Ramirez, D.S.; Lara Perez, M.M.; Carretero Perez, M.; Suarez Hernandez, M.I.; Martin Pulido, S.; Pera Villacampa, L.; Fernandez Vilar, A.M.; Rivero Falero, M.; Gonzalez Carretero, P.; Reyes Millan, B.; et al. SARS-CoV-2 Antibodies in Breast Milk After Vaccination. Pediatrics 2021, 148, e2021052286. [Google Scholar] [CrossRef]
- Pace, R.M.; Williams, J.E.; Järvinen, K.M.; Belfort, M.B.; Pace, C.D.W.; Lackey, K.A.; Gogel, A.C.; Nguyen-Contant, P.; Kanagaiah, P.; Fitzgerald, T.; et al. COVID-19 and human milk: SARS-CoV-2, antibodies, and neutralizing capacity. medRxiv 2020. [Google Scholar] [CrossRef]
- Scrimin, F.; Campisciano, G.; Comar, M.; Ragazzon, C.; Davanzo, R.; Quadrifoglio, M.; Giangreco, M.; Stabile, G.; Ricci, G. IgG and IgA Antibodies Post SARS-CoV-2 Vaccine in the Breast Milk and Sera of Breastfeeding Women. Vaccines 2022, 10, 125. [Google Scholar] [CrossRef]
- Perl, S.H.; Uzan-Yulzari, A.; Klainer, H.; Asiskovich, L.; Youngster, M.; Rinott, E.; Youngster, I. SARS-CoV-2–Specific Antibodies in Breast Milk After COVID-19 Vaccination of Breastfeeding Women. JAMA 2021, 325, 2013. [Google Scholar] [CrossRef]
- Fox, A.; Marino, J.; Amanat, F.; Krammer, F.; Hahn-Holbrook, J.; Zolla-Pazner, S.; Powell, R.L. Robust and Specific Secretory IgA Against SARS-CoV-2 Detected in Human Milk. iScience 2020, 23, 101735. [Google Scholar] [CrossRef] [PubMed]
- van Gils, M.J.; Lavell, A.; van der Straten, K.; Appelman, B.; Bontjer, I.; Poniman, M.; Burger, J.A.; Oomen, M.; Bouhuijs, J.H.; van Vught, L.A.; et al. Antibody responses against SARS-CoV-2 variants induced by four different SARS-CoV-2 vaccines in health care workers in the Netherlands: A prospective cohort study. PLOS Med. 2022, 19, e1003991. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.R.; von Andrian, U.H. Differentiation and homing of IgA-secreting cells. Mucosal Immunol. 2008, 1, 96–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underdown, B.J. IgA. In Encyclopedia of Immunology, 2nd ed.; Delves, P.J., Ed.; Elsevier: Oxford, UK, 1998; pp. 1196–1199. [Google Scholar]
- Atyeo, C.; Alter, G. The multifaceted roles of breast milk antibodies. Cell 2021, 184, 1486–1499. [Google Scholar] [CrossRef]
- Rio-Aige, K.; Azagra-Boronat, I.; Castell, M.; Selma-Royo, M.; Collado, M.; Rodríguez-Lagunas, M.; Pérez-Cano, F. The Breast Milk Immunoglobulinome. Nutrients 2021, 13, 1810. [Google Scholar] [CrossRef]
- Hunagund, S.; Golan, Y.; Asiodu, I.V.; Prahl, M.; Gaw, S.L. Effects of Vaccination Against Influenza, Pertussis, and COVID-19 on Human Milk Antibodies: Current Evidence and Implications for Health Equity. Front. Immunol. 2022, 13, 910383. [Google Scholar] [CrossRef]
- Doria-Rose, N.; Suthar, M.S.; Makowski, M.; O’Connell, S.; McDermott, A.B.; Flach, B.; Ledgerwood, J.E.; Mascola, J.R.; Graham, B.S.; Lin, B.C.; et al. Antibody Persistence through 6 Months after the Second Dose of mRNA-1273 Vaccine for Covid-19. N. Engl. J. Med. 2021, 384, 2259–2261. [Google Scholar] [CrossRef]
- Sobieszczyk, M.E.; Maaske, J.; Falsey, A.R.; Sproule, S.; Robb, M.L.; Frenck, R.W.; Tieu, H.-V.; Mayer, K.H.; Corey, L.; Neuzil, K.M.; et al. Durability of protection and immunogenicity of AZD1222 (ChAdOx1 nCoV-19) COVID-19 vaccine over 6 months. J. Clin. Investig. 2022, 132, e160565. [Google Scholar] [CrossRef]
- Brunner, W.M.; Freilich, D.; Victory, J.; Krupa, N.; Scribani, M.B.; Jenkins, P.; Lasher, E.G.; Fink, A.; Shah, A.; Cross, P.; et al. Comparison of antibody response durability of mRNA-1273, BNT162b2, and Ad26.COV2.S SARS-CoV-2 vaccines in healthcare workers. Int. J. Infect. Dis. 2022, 123, 183–191. [Google Scholar] [CrossRef]
- Jamshidi, E.; Asgary, A.; Shafiekhani, P.; Khajeamiri, Y.; Mohamed, K.; Esmaily, H.; Jamal Rahi, S.; Mansouri, N. Longevity of immunity following COVID-19 vaccination: A comprehensive review of the currently approved vaccines. Hum. Vaccin Immunother. 2022, 18, 2037384. [Google Scholar] [CrossRef] [PubMed]
- Cole, F.S.; Schneeberger, E.E.; Lichtenberg, A.N.; Colten, H.R. Complement biosynthesis in human breast-milk macrophages and blood monocytes. Immunology 1982, 46, 429–441. [Google Scholar] [PubMed]
- McClelland, D.B.; McGrath, J.; Samson, R.R. Antimicrobial factors in human milk. Studies of concentration and transfer to the infant during the early stages of lactation. Acta Paediatr. Scand. Suppl. 1978, 271, 1–20. [Google Scholar]
- Ogundele, M.O. Complement-mediated bactericidal activity of human milk to a serum-susceptible strain of E. coli 0111. J. Appl. Microbiol. 1999, 87, 689–696. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | n = 22 |
|---|---|
| Age, mean (±SD) | 36 (±3) |
| Previous COVID-19 infection, n | 3 |
| Vaccine type Pfizer–BioNtech, n | 17 |
| Vaccine type Oxford–AstraZeneca, n | 4 |
| Vaccine type Oxford–AstraZeneca + Pfizer–BioNtech, n | 1 |
| Days between collection and the 2nd dose of vaccine, mean (±SD) | 27 (±13) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agostinis, C.; Toffoli, M.; Balduit, A.; Mangogna, A.; Yasmin, H.; Ragazzon, C.; Pegoraro, S.; Campisciano, G.; Stabile, G.; Zito, G.; et al. Anti-Spike Antibodies Present in the Milk of SARS-CoV-2 Vaccinated Mothers Are Complement-Activating. Int. J. Mol. Sci. 2023, 24, 4395. https://doi.org/10.3390/ijms24054395
Agostinis C, Toffoli M, Balduit A, Mangogna A, Yasmin H, Ragazzon C, Pegoraro S, Campisciano G, Stabile G, Zito G, et al. Anti-Spike Antibodies Present in the Milk of SARS-CoV-2 Vaccinated Mothers Are Complement-Activating. International Journal of Molecular Sciences. 2023; 24(5):4395. https://doi.org/10.3390/ijms24054395
Chicago/Turabian StyleAgostinis, Chiara, Miriam Toffoli, Andrea Balduit, Alessandro Mangogna, Hadida Yasmin, Chiara Ragazzon, Silvia Pegoraro, Giuseppina Campisciano, Guglielmo Stabile, Gabriella Zito, and et al. 2023. "Anti-Spike Antibodies Present in the Milk of SARS-CoV-2 Vaccinated Mothers Are Complement-Activating" International Journal of Molecular Sciences 24, no. 5: 4395. https://doi.org/10.3390/ijms24054395